
海域隔离对桔小实蝇种群形态特征的影响
2021-10-14 02:35易春燕郑春艳许益镌
陕西农业科学 2021年8期
易春燕,郑春艳,刘 旭,曾 玲,许益镌
(1.四川省农业科学院 植物保护研究所/农业部西南作物有害生物综合治理重点实验室 ,四川 成都 610016;2.华南农业大学 昆虫学系昆虫生态研究室 ,广州 510640;3.华南农业大学 生命科学学院 ,广州 510640)
形态特征是遗传特征的外部表现,是遗传和环境因子的共同作用的结果。物种个体成功入侵且成功繁殖,都会对入侵地环境作出相应反应,从而有一个相对适应生存环境的形态特 征,这种适应性不仅与地理隔离程度有关,而且受自然选择压力的影响[1~2]。
地理阻隔是限制物种扩散和影响遗传多样性的重要因素,因此,岛屿作为一种天然的屏 障在影响昆虫的遗传多样性上起着关键作用,因为岛屿相对封闭,降低了岛屿种群与大陆种 群之间的交流。岛屿与内陆相比较往往具有更简单和独特的生态环境,而岛屿生物的竞争者 和捕食者也可能也相对较少,具有不同食物、环境、栖息场所[3]。岛屿上的生物有两种来源方式,一种是岛屿与大陆分离之前就已经有分布的,另外一种就是后来自由扩散来的或者人 为有意无意引进的[4~5]。而引进的这些物种在适应岛屿的生态环境时将在分子水平或者形态上产生一些改变。有研究表明,岛屿生物因为生物多样性或气候条件的变化个体尺寸会做一 些改变[6]。
桔小实蝇Bactroceradorsalis(Hendel),又名东方果实蝇(orientalfruitfly),俗称果蛆, 隶属于昆虫 Insecta,双翅目 Diptera,实蝇科 Tephritidae,果实蝇属BactoceraMacquart,是一种世界危险性检疫害虫。该虫为杂食性害虫,寄主范围广泛,可危害番石榴、杨桃、香蕉等 46 科 250 多种果树及蔬菜[7~10]。桔小实蝇具备很强的迁飞能力,在密度高的情况下,会主动向外界迁移扩散,在降落在不适合的寄主上后,会再次起飞直到找到适合的寄主。据国内外研究报道发现,在日本小笠原群岛上,桔小实蝇能飞越 50 km 的海面[11]。此外,桔小实蝇会借助外在因素可远距离迁飞,桔小实蝇可以从台湾本岛随东北风飞到 27 km 以外的小琉球岛,桔小实蝇这种远距离迁飞能力使其分布范围更广[12]。虽然桔小实蝇具备一定的飞翔能力,但是海域等天然屏障却有可能影响其种群的遗传和分化。
目前,已有不少文献利用分子手段报道了地理隔离对桔小实蝇种群遗传多样性的影响[13~15]),而海域对桔小实蝇形态特征及遗传分化的影响却鲜有研究。本文通过测量华南地区5个离岛种群与 9 个大陆桔小实蝇种群形态特征的 3 个指标,分别为三个横脉的长度(L1、L2、L3)和体现形态细微变异的翅脉角(α、β、γ)以及翅面积,并对岛屿种群与大陆种群的比 较进行分析,这将对进一步探究桔小实蝇在特殊地理环境下的遗传多样性和传播能力提供理 论依据。
大陆的几个采样点均具有桔小实蝇适宜的寄主植物,且环境均比较适宜桔小实蝇生存。 各岛屿距离大陆的直线距离在 4.8-38.4 km,虽然有的岛屿上有种植香蕉、木瓜等植物,但是种植的其它桔小实蝇寄主植物比较少,桔小实蝇的种群规模小(上川岛除外),各岛都有进行旅游业的开发,贩卖从大陆运输的水果和蔬菜。
1 材料与方法
1.1 材料
桔小实蝇成虫样本采自华南地区的 5 个离岛:涠洲岛、硇洲岛、上川岛、荷包岛、东澳岛,以及华南地区湛江市等几个大陆种群(见表 1)。采集时用甲基丁香酚引诱剂桔小实蝇雄虫,每个地区至少 20 头,浸泡在无水乙醇中,4℃保存,以备进行后续试验。
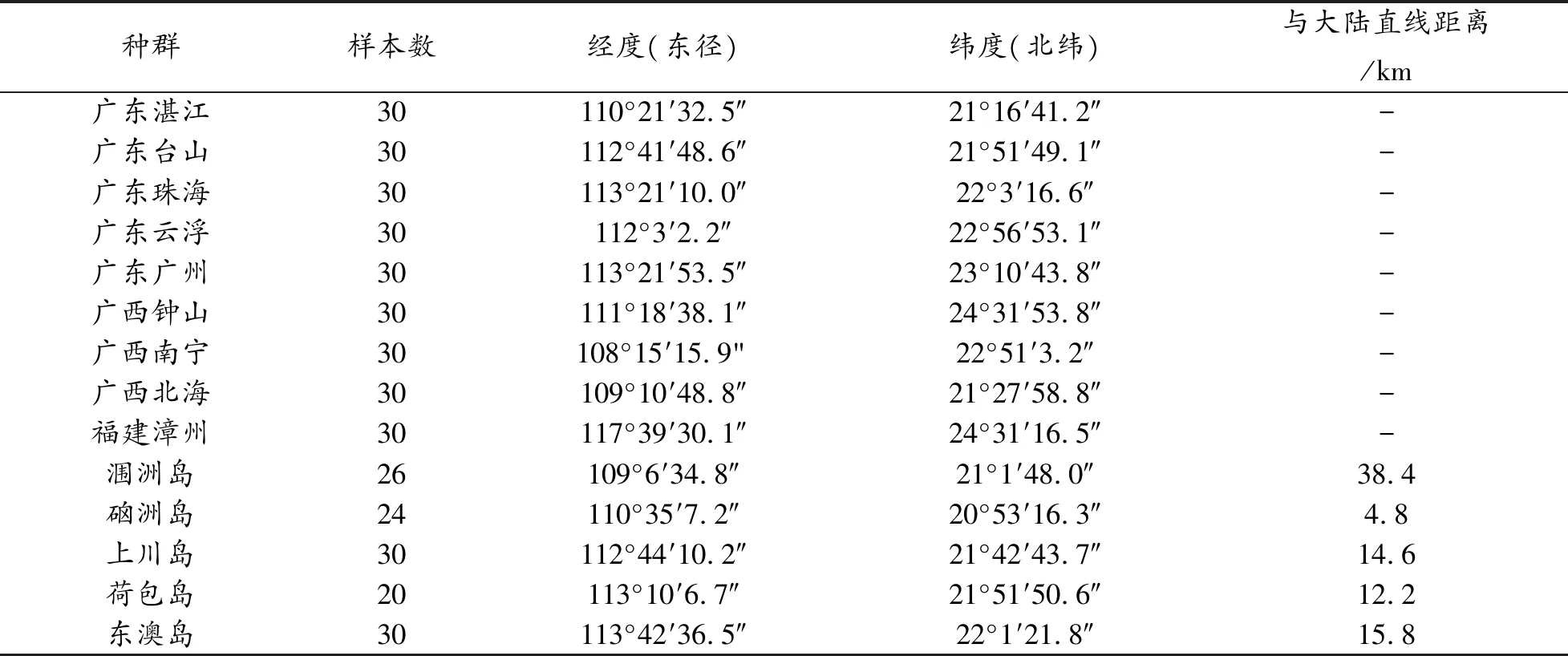
表1 样本信息
1.2 形态测定
有研究者利用身体大小的指标对黑腹果蝇的遗传结构和遗传差异进行了研究,研究表明翅膀的大小和身体大小具有高度的相关性,而翅膀的面积与翅膀大小成正相关[16~17]。因此, 我们参照果蝇的形态标记方法对桔小实蝇样本进行标记,测定了翅面积大小、三个横脉长度、 三个翅脉角。
将桔小实蝇的翅膀用镊子小心拔取,压片。用带有图形采集器与电脑形态测量系统(Axiovision SE64 Rel.4.9.lnk)的德国蔡司 V20 体式显微镜(SteREO Discovery.V20,上海冉超光电科技有限公司)联用技术在 15X 下拍照和形态测量。选取了 A-H 八个翅脉交接点做一个多边形,以这个多边形的面积作为翅膀的面积来计算,同时测量三个横脉的长度(L1、L2、 L3)和体现形态细微变异的翅脉角(α、β、γ),如图 1。

图1 形态测定指标(翅面积 S:以 A-H 八根线段围成的多边形面积来计算,横脉长 L1、L2、L3,翅脉角α、β、γ)。
1.3 数据处理方法
使用SPSS 22.0 软件计算各形态参数的平均值和标准差,并进行单因素方差分析和Tukey多重比较,对翅面积大小与纬度进行相关性分析。将所有种群分成大陆组与岛屿组对各形态指标进行T检验,检验岛屿与大陆桔小实蝇种群是否存在显著差异。
2 结果与分析
2.1 形态差异
各参数的平均值见表2,横脉长度 L1 449.5~493.5,L2 605.5~655.7,L3 793.5~860.3, 翅膀面积 S 7 940 096~9 021 667.6,且翅脉长度(L1、L2、L3)与翅面积 S 显著性相关(p< 0.001);角度α65.3~67.5 度,β 59.4~60.3 度,γ 100.5~102.8度。通过差异显著性分析,各种群的翅面积(图 1)S (F(13, 390)=3.674,P<0.01)、横脉长度 L1(F(13, 390)=3.927,P<0.01)、 L2 (F(13, 390)= 2.854,P<0.01)、L3( F(13, 390)=3.659,P<0.01)存在明显差异;而三个翅脉角α(F(13, 390)= 2.275,P=0.838)、β(F(13, 390)=0.603,P=0.852)、γ(F(13, 390)= 1.5,P=0.114)没有明显差异,各参数情况见表2。
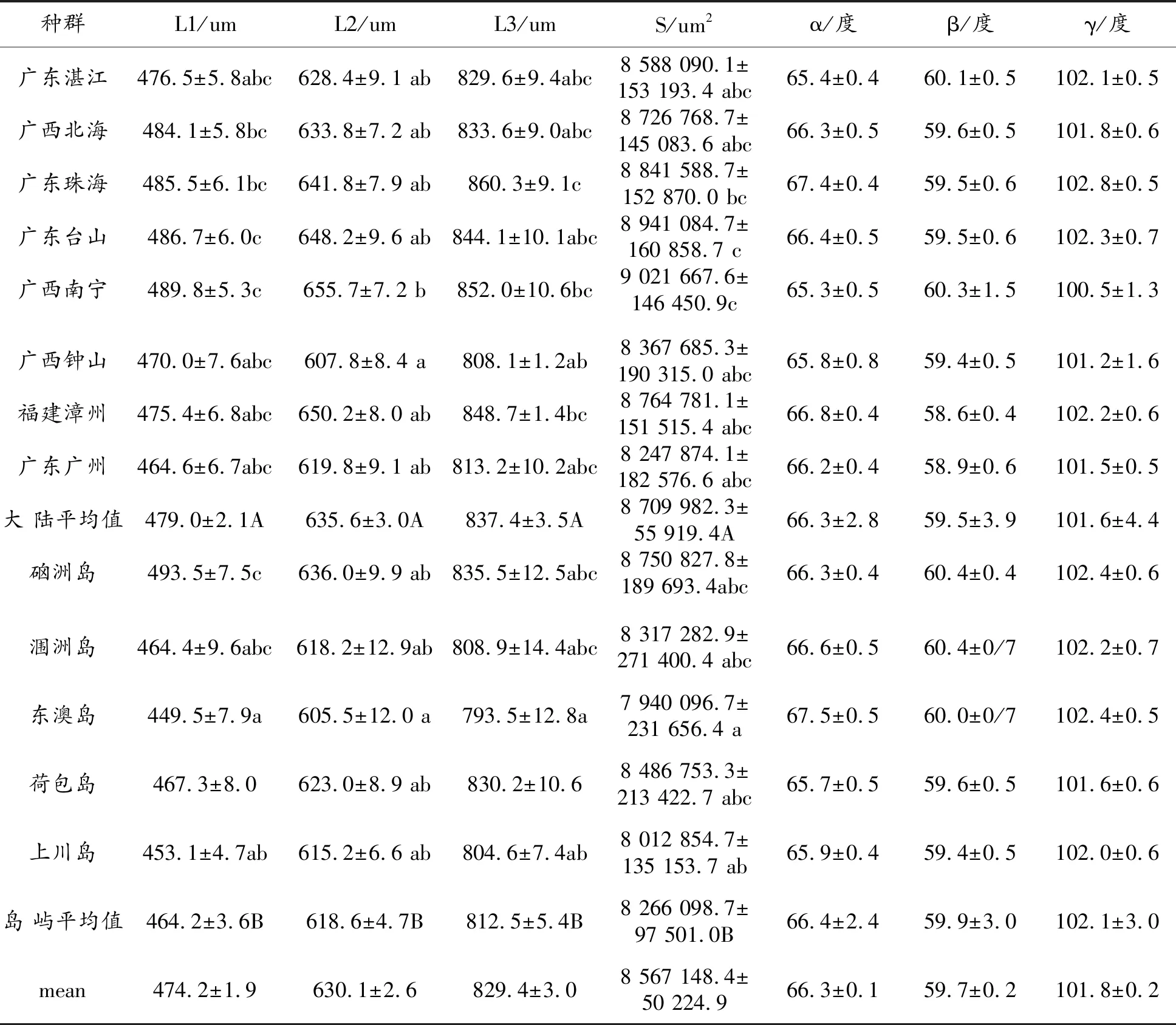
表2 各种群形态差异分析
将种群分为大陆种群与岛屿种群进行 T 检验发现, 岛屿种群各参数中的翅面积 S(t=3.949,df=216.7,P<0.01)、翅脉长度 L1(t=3.523,df=220,P<0.01)、L2(t=3.152,df=402,P<0.01)、L3(t=3.983,df=402,P<0.01)显著性小于大陆种群,各翅脉角未存在显著差异(表 2) 。
2.2 相关性分析
2.2.1 形态遗传标记的相关性分析 对5个岛屿种群及9个大陆种群的400头桔小实蝇各形态标记进行相关性分析,分析各标记的相关程度。反映桔小实蝇个体大小的三个横脉长度之间相关性显著相关系数均大于0.768(p 均小于 0.01),横脉长度与翅膀面积大小之间也具有显著相关性,相关系数均大于0.856(p<0.01)(图 2)。三个翅脉角标记间,∠1-∠2 无相关性;∠1-∠3 存在显著性相关,相关系数为 0.139(p<0.01);∠2-∠3 呈显著负相关,相关系数为-0.151(p<0.01)。
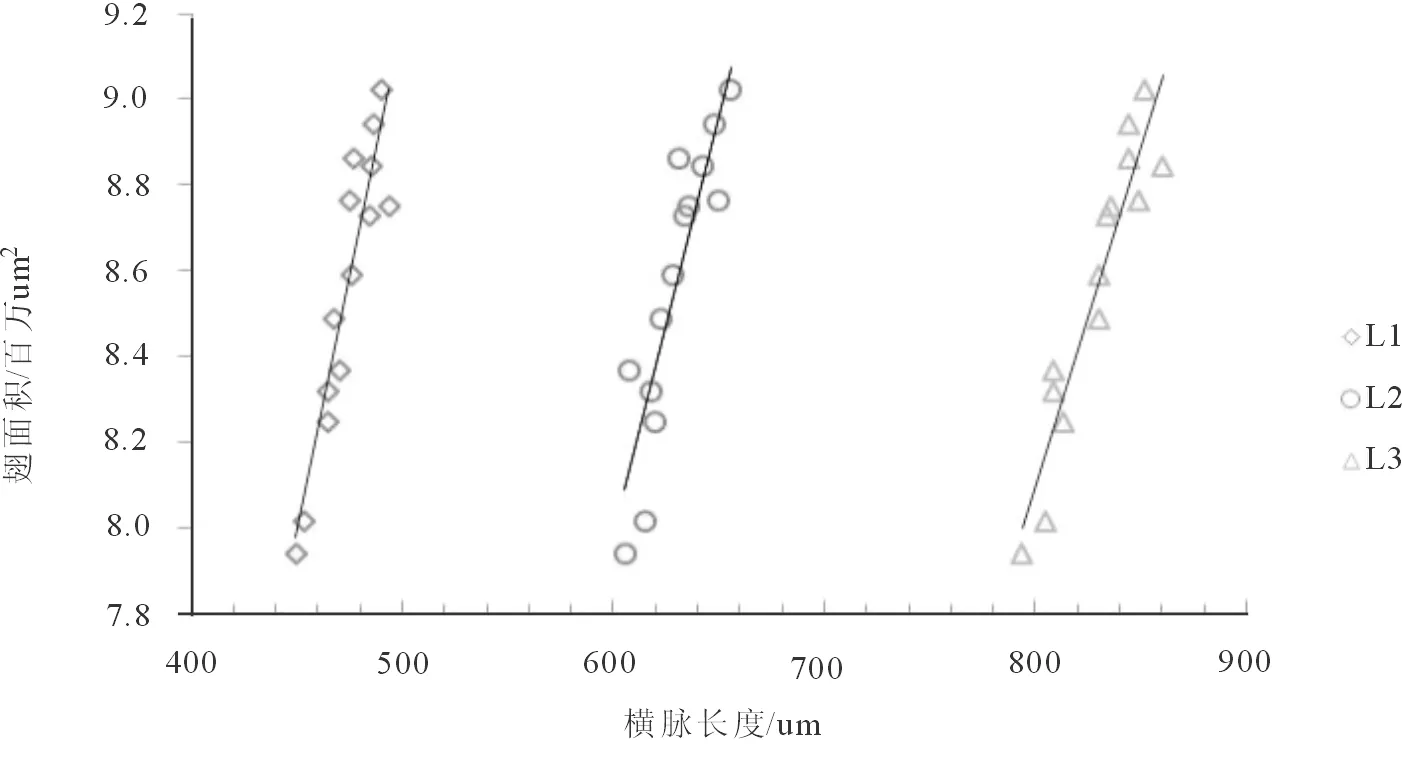
图2 横脉长度与翅面积大小的相关性分析
2.2.2 个体大小与纬度相关性分析 经研究显示大陆种群与岛屿种群的形态大小之间存在显著差异,为了研究是否因纬度差异引起的,我们将个体大小与纬度大小进行了分析。由图 3 翅面积大小与纬度相关性分析可以看出,个体大小与纬度无显著相关( P=0.561)。
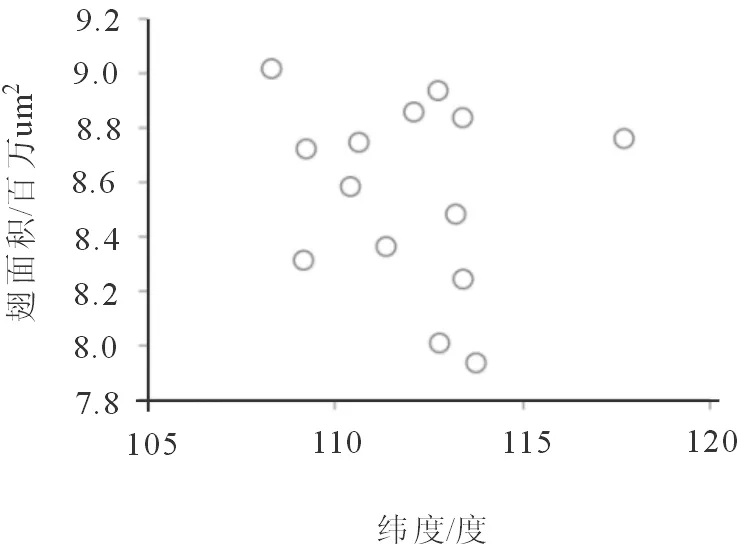
图3 纬度与翅面积大小的相关性分析
3 讨论与结论
通过比较研究发现桔小实蝇岛屿种群形态指标当中的翅面积大小、横脉长度明显小于大陆种群,形态差异当中的翅脉角没有显著差异。虽然纬度差异对影响海岛种群的形态差异 上的作用不大,但海域隔离使得岛屿桔小实蝇在适应海岛环境的时候在形态上发生了一些变 化。有研究表明岛屿生物会因为食物、天敌、温度、风等生物因素、非生物因素的环境压力 驱动着岛屿生物在形态大小、体色等形态特征上做出改变[6, 18~19]。据采样时调查,虽然有的岛屿上有种植香蕉、木瓜等植物,但是种植的桔小实蝇的其它寄主植物比较少,这可能是岛 屿桔小实蝇形态偏小的其中一个原因;另外,海岛上的海风很大,经常还会有飓风,桔小实 蝇也许是通过改变形态大小来抵御这种环境。
形态遗传学研究方法操作简单,而且快速、成本低,但是也容易受数据测量分析等因素的影响,因此需要我们在应用时多加注意,制定规范的测量操作,以便保证测量的准确度。 本研究采用了翅面积、三个横脉长度和三个翅脉角等三个主要形态指标,在今后的研究当中可以适当增加一些形态遗传指标,以提高结果的可信度。我们在研究过程中发现三个横脉的长度与翅面积具有显著相关性,可以选取具有高度相关性的形态指标中的部分作为代表性指标,这样可以降低一些测定工作量[20~21]。
形态上的差异往往有其内在的分子机制。Thorpe[22]研究大加纳利岛的西加纳利蜥蜴时发现有 30%的形态特征与 mtDNA 中的 cytb、COⅠ 和 12s rRNA 有显著相关性。为了深入研究形态差异的原因,我们也可以进一步利用微卫星与 COⅠ标记等分子手段研究桔小实蝇岛屿种群在分子水平上的变异,验证形态与分子变异是否有相关性。
猜你喜欢
今日农业(2022年15期)2022-09-20
湖北农业科学(2022年9期)2022-06-01
中国南方果树(2022年2期)2022-04-15
思维与智慧·上半月(2022年4期)2022-04-08
四川文学(2020年10期)2020-02-06
生物学教学(2018年3期)2018-08-08
新农村(2018年32期)2018-04-02
中学生物学(2018年8期)2018-03-01
儿童故事画报(2015年5期)2015-07-22
时代青年(上半月)(2009年6期)2009-07-31
