
红松叶片与土壤有机碳、氮、磷、钙的空间分布及其化学计量学特征
2021-10-12 01:49魏亚伟秦胜金杜天雨周永斌
沈阳农业大学学报 2021年4期
魏亚伟,张 彤,刘 静,韩 叙,秦胜金,杜天雨,周永斌
(1.沈阳农业大学林学院,沈阳 110161;2.辽宁辽河平原森林生态体系定位研究站,辽宁昌图 112500)
生态化学计量主要指植物的元素(主要是碳、氮、磷)组成及其相互关系[1],是揭示元素相互作用与制约的变化规律[2]、元素平衡与生态交互作用影响的一种有效方法[3],也是研究生态系统营养元素分配、循环过程以及限制性元素判断的一种新兴手段[4]。近年来,随着生态学的快速发展,生态化学计量学的研究已经成为当前研究的热点之一,受到国内外学者的广泛关注[5],同时也为揭示森林土壤养分的循环过程、利用规律及反馈作用机制提供了新思路[2]。例如,对不同植被类型、不同演替阶段的植物器官的化学计量特征进行研究,对比植物组织中C、N、P化学计量比的差异,探讨不同植被类型或不同演替阶段对养分的影响规律[6-7]。此外,关于森林和草原生态系统C∶N∶P生态化学计量学特征及其驱动因素[5]、全球与区域、功能群及生态系统、个体水平等不同尺度上的生态化学计量学特征等[5]、氮沉降或氮、磷添加等对生态系统化学计量学特征的影响等也有较多报道[8-9]。
钙是地壳中含量最多的五种元素之一,也是自然界中丰度最高的元素之一。同时,钙作为植物必需的营养元素之一,对维持森林生态系统稳定有重要作用。钙既是植物细胞壁的主要组成成分,又是植物生理过程中的重要信号调节因子[10],在植物生长发育以及适应逆境胁迫过程中处在中心的调控地位[11]。近年来,随着长期定位观测在生物地球化学循环等方面的应用,发现钙对生物地球化学循环的大多数环节都有影响,并表现出明显的生态效应,如植物发芽能力、幼苗成活率、森林生产力及群落演替等[12-13]。不仅如此,钙缺失还将改变森林生态系统中的物种组成和生物多样性[14-16]。而环境的酸化将对生态系统中钙的平衡造成巨大影响,如钙供应不足会严重影响植物的正常生长,尤其在环境胁迫时将更加明显[17],钙已经成为限制森林生态系统生产力的主要因素之一[18-19]。随着氮沉降的增多,钙有可能取代氮而限制树木的生长[20]。因此,探讨森林生态系统钙元素的空间分布特征、及其与化学计量学特征的关系对更好地理解森林生态系统的生物地球化学循环具有重要的科学意义。
阔叶红松林是我国东北地区的主要森林类型之一,同时也是东北温带地区的地带性顶极植被群落[21]。然而,关于阔叶红松林生态系统钙元素却鲜有报道。本研究以阔叶红松林中的红松为研究对象,研究其土壤及叶片钙元素的空间变异性、以及与土壤、叶片C、N、P及其化学计量学特征的相互关系,以期为东北阔叶红松林的科学管理及未来气候变化条件下更好地预测森林生态系统的养分循环提供参考。
1 材料与方法
1.1 研究区概况
阔叶红松林主要分布在我国东北的长白山和小兴安岭地区。该地区是我国典型的北温带湿润气候区,受温带大陆性气候控制,冬季寒冷干燥且漫长,夏季湿热多雨且短暂,年均温1.8~4.8℃,年降水量550~700mm,降水多集中在夏季,但由于该地区常年的低温,年蒸发量较小,加上森林自身的调节作用,该区能常年保持湿润。阔叶红松林是该地区的顶极植被群落,以喜湿的红松(Pinus koraiensis)为优势种,伴生有紫椴(Tilia amurensis)、蒙古栎(Quercus mongolica)、色木槭(Acer mono)、榆树(Ulmus mongolica)、水曲柳(Fraxinus mandshurica)等,土壤类型以暗棕色森林土为主。
1.2 样地设置与样品采集
2015年7~8月分别在长白山林区的露水河、张广才岭地区的穆棱、小兴安岭地区的五营选择典型的天然阔叶红松林,林龄均为成、过熟林(表1)。在每个研究区分别选取典型的林分设置样地,样地面积均为20m╳20m,调查样地内的所有乔木,并根据红松的各项生长指标的平均值,在样地内选择3~5株中等大小、长势及冠幅较一致的健康标准木,利用高枝剪分别从每棵树的东、南、西、北4个方向、树冠上、中、下3个层次采集样树的叶片,混匀后作该样树的样品;同时,利用土钻在每株树的四周分别进行土壤样品的采集,采集深度分别为表层0~15cm(表层)和15~30 cm(下层),挑出其中的根系和石砾,混匀后带回室内分析。
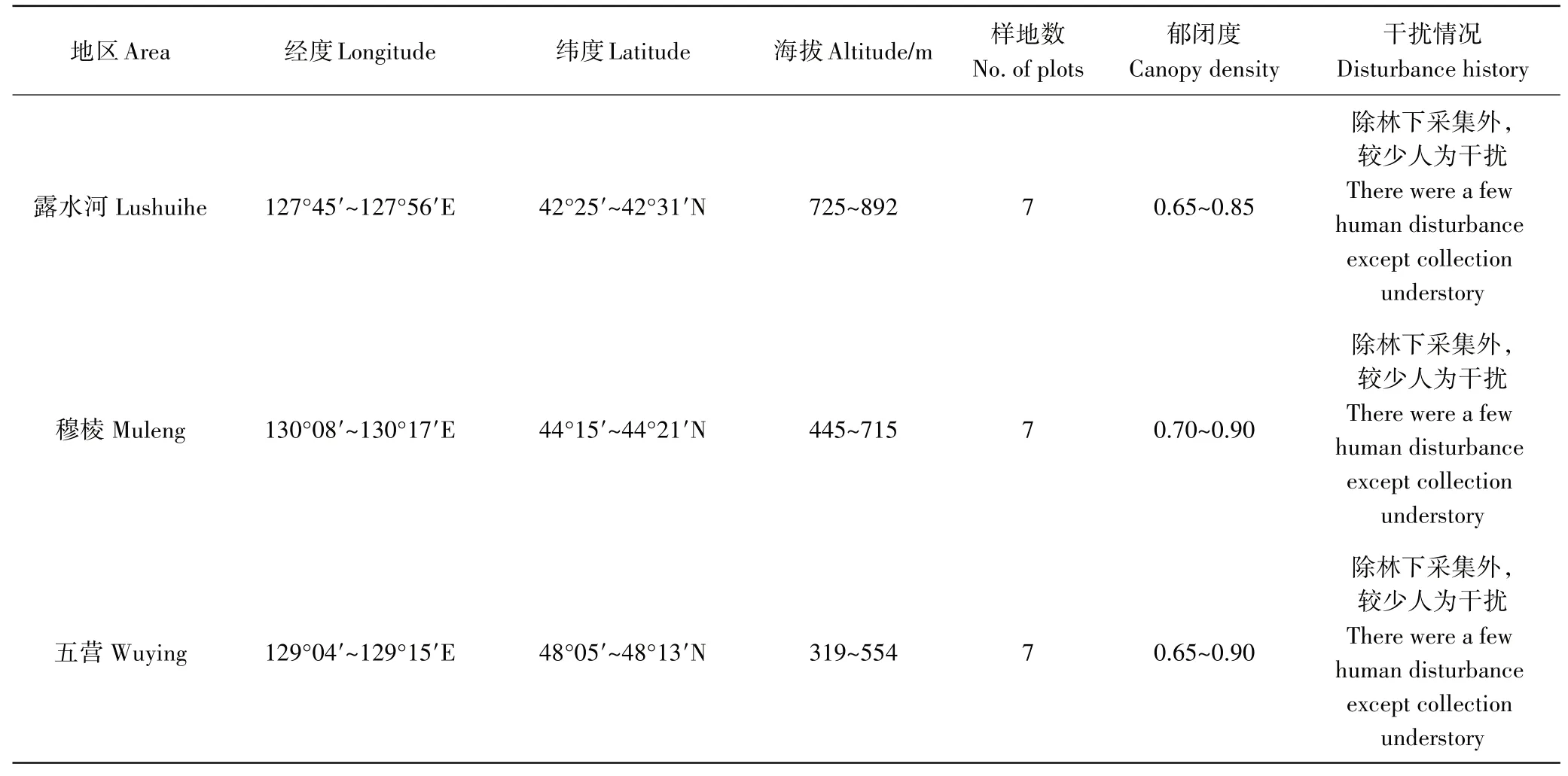
表1 样地基本概况Table 1 Basic information of the plots
将采集的叶片于105℃下杀青30min,然后65℃下烘干至恒量,并经粉碎机打磨后过100目筛。土壤样品经风干、粉碎后过100目筛。样品有机碳的测定采用重铬酸钾-外加热法,全氮的测定采用凯氏定氮法,全磷的测定采用硫酸高氯酸消煮-钼蓝比色法、分光光度计测定。
1.3 数据处理
采用Microsoft Excel 2013软件对数据进行处理和绘图。采用SPSS 17.0软件对数据进行统计分析,正态分布性检验采用K-S检验(one sample Kolmogorov-Smirnov test)。对植物叶片、土壤的C、N、P、Ca含量及其化学计量比数据进行以10为底的对数转换,使其符合正态分布;采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较不同土层C、N、P和Ca含量特征的差异性,各元素间及与其化学计量特征间的相关分析采用Pearson相关。
2 结果与分析
2.1 阔叶红松林土壤、叶片有机碳、全氮、全磷、钙元素的含量特征
由表2和图1可知,阔叶红松林表层土壤有机碳(TOC)、全氮(TN)、全磷(TP)的变化范围分别为2.3%~11.0%,0.2%~0.8%,0.02%~0.1%,土壤水溶性钙的含量相对较少,仅为25.33~86.30mg·g-1;下层土壤各元素含量较表层均有不同程度的减少,TOC、TN、TP和Ca分别为0.6%~6.8%,0.1%~0.5%,0.02%~0.1%,16.77~82.37mg·g-1。整体来说,土壤各元素的空间变异性均较大(变异系数CV>20%)。
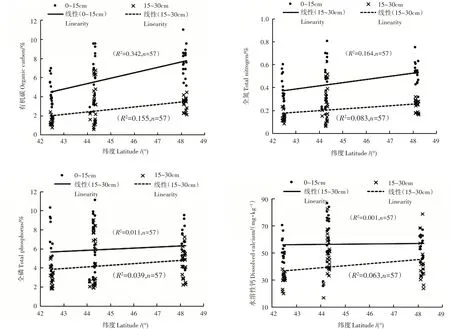
图1 土壤有机碳、全氮、全磷和水溶性钙的含量特征Figure 1 Characteristics of organic carbon,total N,total P,dissolved Ca concentration in soil
叶片的TOC、TN和TP均较土壤显著升高,分别为48.8%~51.2%,1.2%~1.7%和0.1%~0.3%,但叶片Ca含量较土壤显著降低,仅为1.88~4.29mg·g-1(表2和图2)。叶片SOC和TN的变异性相对较小,变异系数仅为0.9%和9.1%,但叶片TP和Ca的变异系数均较大(CV>20%)。
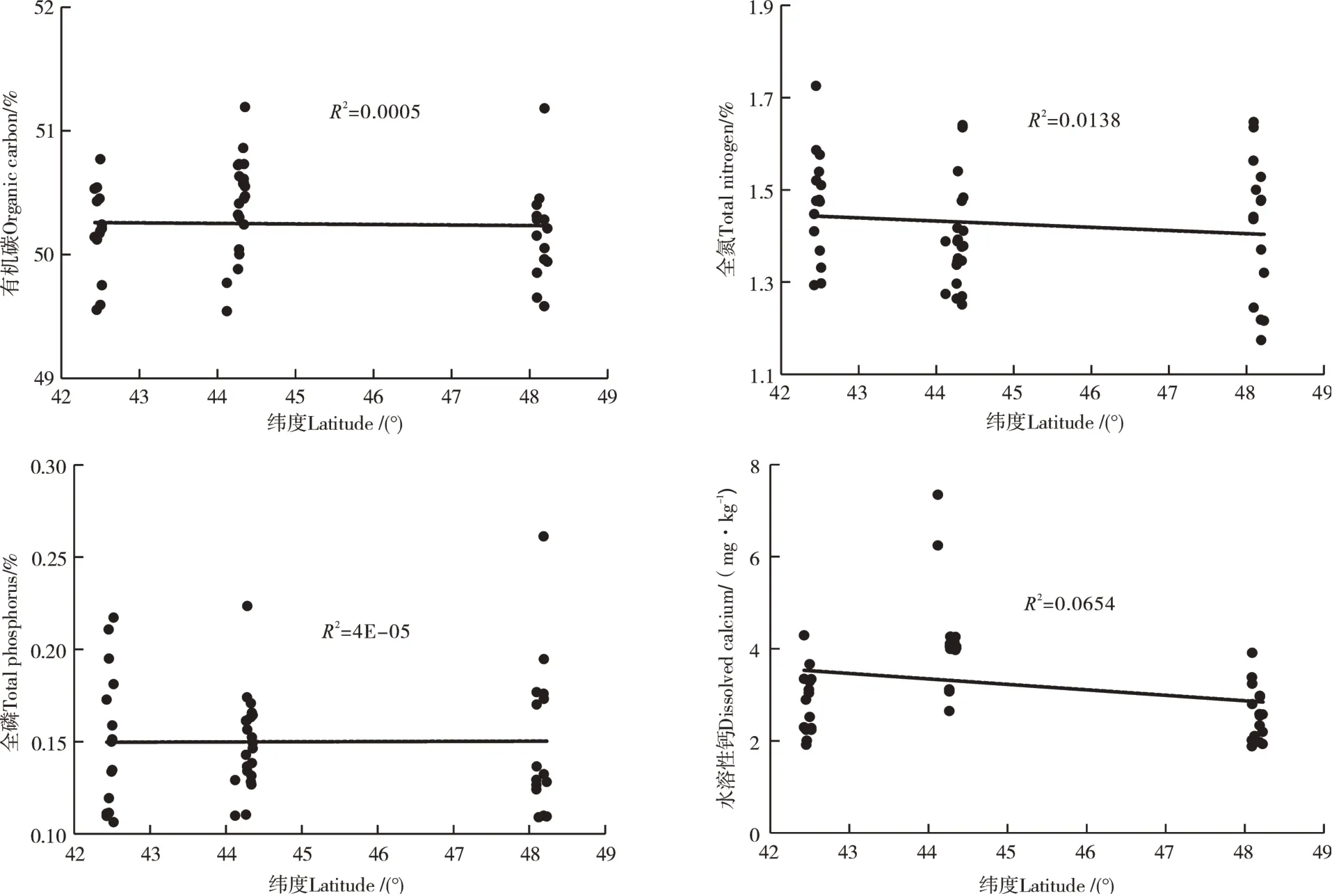
图2 叶片有机碳、全氮、全磷和钙的含量特征Figure 2 Characteristics of organic carbon,total N,total P,dissolved Ca concentration in leaf
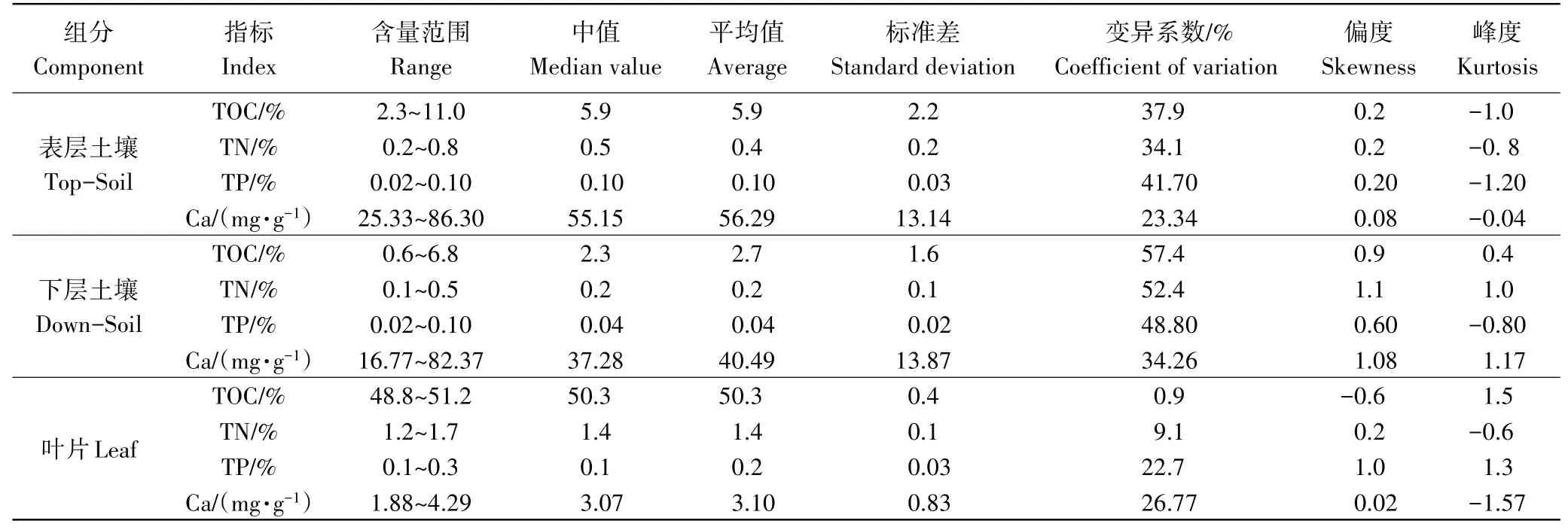
表2 土壤及叶片C、N、P、Ca含量特征Table 2 Characteristics of C,N,P,Ca concentration of soil and leaf
2.2 阔叶红松林土壤、叶片有机碳、全氮、全磷的化学计量学特征
表层土壤C/N、C/P、N/P的变化范围分别为10.50~18.62,41.24~195.79,3.50~13.08,均高于下层土壤(表3和图3);叶片的C/N和C/P均较大,分别为28.73~43.60和189.81~472.22,显著高于土壤,而N/P与土壤的差异较小,为5.65~13.76;土壤和叶片的C/N变异性均较小,变异系数均小于15%,而C/P和N/P的变异性则均相对较大(表3)。
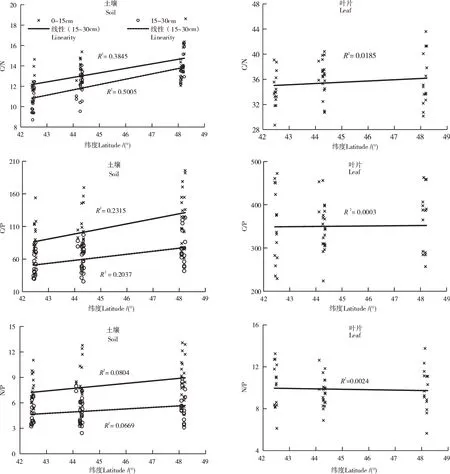
图3 土壤、叶片中C、N、P的化学计量学特征Figure 3 Characteristics of soil and leaf C,N,P stoichiometry.
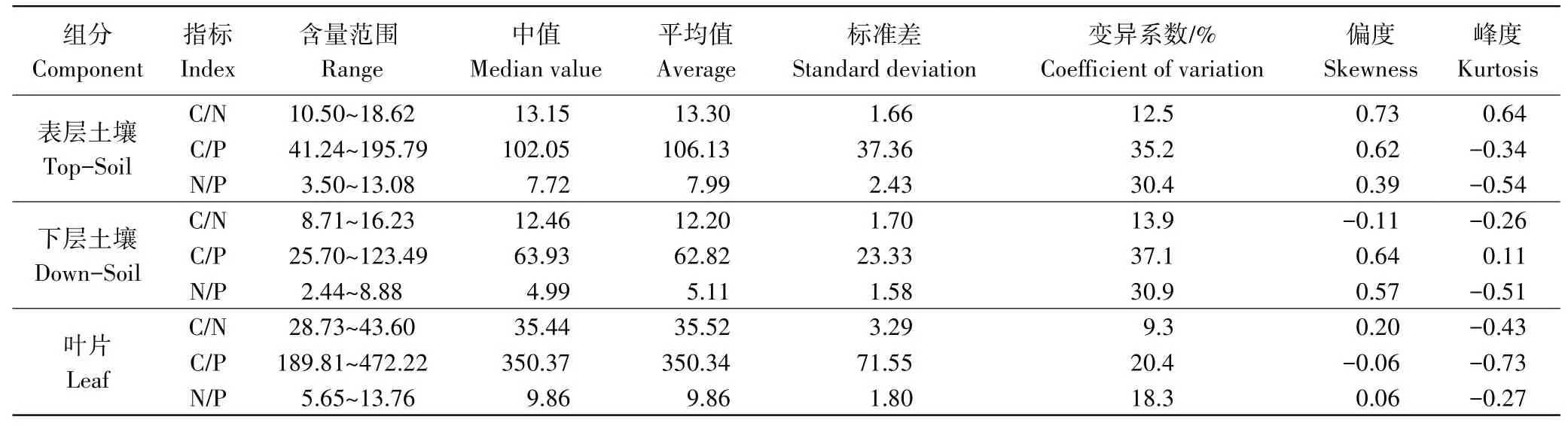
表3 土壤及叶片C、N、P化学计量学特征Table 3 Characteristics of soil and leaf C,N,P stoichiometry
2.3 阔叶红松林土壤、叶片钙与有机碳、全氮、全磷及其化学计量学的相关性
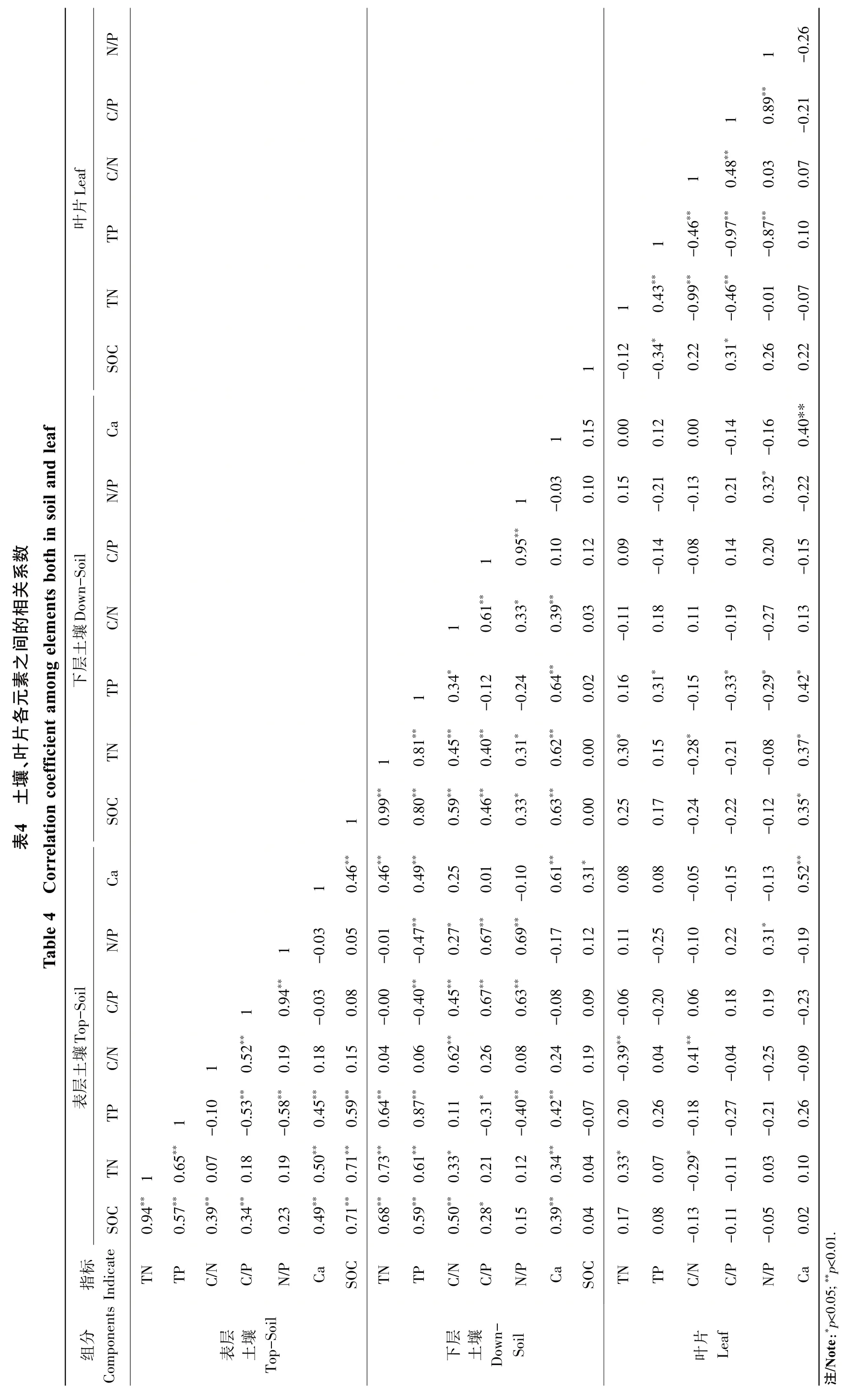
由表4可知,表层土壤水溶性钙含量与表层土壤和下层土壤SOC、TN和TP、以及下层土壤Ca、叶片SOC和Ca含量均呈现显著正相关关系(p<0.05),但与C、N、P化学计量学特征的关系均不显著;深层土壤水溶性钙含量与表层土壤和下层土壤SOC、TN、TP、以及表层土壤Ca、深层土壤C/N和叶片Ca含量均呈现极显著正相关关系(p<0.01);叶片Ca仅与土壤Ca及深层土壤SOC、TN和TP呈现显著的正相关。此外,表层土壤SOC与土壤TN、TP、C/N和C/P均呈显著正相关;土壤TN和TP与叶片TN、TP呈显著正相关,与C/N、C/P、N/P则呈显著负相关;叶片TN和TP与叶片C/N、C/P、N/P也均呈现极显著负相关。
3 讨论与结论
森林土壤是森林生态系统的重要组分和养分储存库[2],土壤养分含量在空间和时间上均表现出巨大的差异性[22]。本研究中,有机碳、全氮、全磷及其化学计量比均随纬度的增加呈现出一定空间变异性(图1和图2)。同时,东北阔叶红松林土壤有较高的土壤养分含量,且主要集中在表层土壤,这与以往的研究结果类似,长白山和小兴安岭针阔混交林0~20cm土壤贮存了其剖面总有机碳的50%以上[23]。本研究中表层土壤有机碳和全氮含量,分别为2.28%~10.99%和0.16%~0.80%,远高于全国土壤平均水平1.11%和0.11%,全磷的变化范围(0.02%~0.11%)与全国土壤的平均水平相当(0.06%)[24]。土壤有机碳和氮主要来自于地表枯落物的分解输入,土壤磷主要来源于母质,较少来自于植物分解[2],这说明该地区具有较高的土壤有机碳固持能力,而该地区常年的低温则是限制母质分解并导致土壤磷的含量相对较低的原因之一。叶是植物养分的主要储存库,也是进行光合、呼吸和蒸腾等一系列生理过程的主要场所。东北阔叶红松林叶的有机碳、全氮和全磷含量分别为48.77%~51.19%、1.17%~1.73%、0.11%~0.26%,其中有机碳含量明显高于我国东部南北样带森林生态系统植物(48.0%)和全球陆生植物(46.2%),而全氮和全磷含量与我国陆生植物[(1.75%~2.02%)和(0.13%~0.20%)]和全球陆生植物(2.01%和0.18%)相比均较低[25-26]。这说明该地区红松具有较高的碳蓄积能力[27];植物叶片对环境的变化较为敏感,尤其氮、磷含量受自身属性、生长阶段以及外在环境的影响较大[28],东部地区常年的低温对植物本身的某些物理特性和一系列的新陈代谢过程的影响导致了叶片对氮和磷等部分元素较低的吸收[29]。
?
相比土壤有机碳、全氮和全磷含量对土壤质量的直接反映,生态系统碳、氮、磷的化学计量比特征是反映C、N、P循环与平衡的重要参数,如土壤C/N在一定程度上指示着有机质的分解速率,C/P反映土壤中磷的有效磷高低,土壤N/P则作为土壤养分限制类型的预测指标[2]。本研究中不同区域植被和土壤的C∶N、C∶P和N∶P之间存在较大的差异性,这与全国土壤C∶N∶P分布特征一致[24]。东北阔叶红松林表层和下层土壤C∶N分别为13.3和12.2,均与我国陆地土壤和森林土壤C∶N的比值(12.3和13.7)接近[24,30],这说明该地区较高的土壤有机碳和全氮的固持能力。与我国陆地土壤和森林土壤相比,该地区有较高的土壤C∶P和N∶P比值,尤其表层土壤的C:P高达106.13,这进一步验证了该地区较低的土壤全磷水平。生态系统中叶片C∶N和C∶P不仅代表着不同植物或群落固碳效率的高低[31],也能反应出植物对N、P的利用效率,能在一定程度上判断外界环境对植物生长所需N、P的供应状况[2]。该地区红松叶片的C∶N和C∶P分别为35.52和350.34,均高于全球平均水平(C:N为22.5,C∶P为233.2)[32]和我国东部南北样带森林生态系统植物的水平(C∶N和C∶P分别为29.1和313.9)[27],相反,红松叶片的N∶P(9.86)却明显低于全球平均水平(12.7)[32]和我国东部南北样带森林生态系统植物的水平(11.5)[27]。氮和磷作为陆地生态系统最主要的限制性养分元素,在植物体结构和功能上存在一定的联系,其含量能够直接影响植物的生长[33]。已有研究表明,当叶片N∶P<14时,植物生长主要受N元素限制,当N∶P>16时,植物生长主要受P限制;处于两者之间时N、P同时限制或养分充足不受限制[34]。这进一步证明该地区土壤全磷含量的低水平,而对土壤有机碳则具有较强的固持和保存能力。
土壤中钙的含量既受成土母质的影响,同时也与成土过程中地球化学作用强度有一定关系[35]。本研究中,土壤钙含量和植物钙含量的变化范围分别为1.67%~8.63%和0.19%~0.43%,这均与以往研究相似,即植物体内钙含量一般为0.1%~5.0%[36],北方土壤的钙含量一般在1%~10%。钙供应不足会严重影响植物的正常生长,提高土壤钙含量可以增加植物的抗逆性[17]。钙对生物地球化学循环的大多数环节都有重大影响,本研究发现,不同土层土壤钙含量以及植物叶片钙含量之间均存在极显著的正相关性,这与以往研究结果类似,植物钙主要来自于土壤,且在植物体内难以转移[37]。土壤钙均与表层和下层土壤有机碳、全氮和全磷含量表现出极显著的相关性,而叶片钙含量则受下层土壤有机碳、全氮和全磷的影响较大,呈现出显著的正相关性,其具体机制有待于进一步研究。此外,表层土壤钙含量和叶片钙含量均与土壤和叶片的C∶P和N∶P呈现出明显的负相关关系,但并未达到显著水平,这可能与钙作为重要的信号调节因子,通过调控气孔开闭以及钙调蛋白活性,对植物碳、氮、水代谢过程产生影响等有关[38],而森林生态系统中钙与各元素间的关系以及钙对各元素吸收和保存的调节机制仍有待于进一步的深入研究。
此外,本研究中叶片有机碳含量与叶片的全氮和全磷均呈的负相关,而氮和磷呈显著的正相关,这与黄小波等[39]研究结果相似,植物C含量变化与N、P含量变化呈现相反趋势,这符合高等陆生植物C、N、P元素计量的叶C与N、P的负相关性以及N与P的正相关性这一普遍规律[40]。
猜你喜欢
小哥白尼(野生动物)(2021年9期)2022-01-17
中国林副特产(2020年5期)2020-10-09
防护林科技(2017年7期)2017-09-01
浙江农业学报(2016年7期)2016-06-15
现代农业(2016年3期)2016-04-14
中国农业文摘-农业工程(2016年5期)2016-04-12
现代农业(2016年4期)2016-02-28
土壤与作物(2015年3期)2015-12-08
环境科技(2015年2期)2015-11-08
满族文学(2014年5期)2014-09-22
