
新疆天山北坡不同盐湖微生物菌群结构及其影响因子
2021-10-12 01:13:06李二阳马雪莉吕光辉
生态学报 2021年18期
李二阳,马雪莉,吕 杰,马 媛,3,*,吕光辉,3
1 新疆大学资源与环境科学学院,乌鲁木齐 830046 2 新疆大学生命科学与技术学院,乌鲁木齐 830046 3 绿洲生态教育部重点实验室,乌鲁木齐 830046
新疆面积大于1 km2的天然湖泊有139个,20世纪50年代以来受干旱气候和上游农业灌溉用水的影响,成盐作用强烈,近半数湖泊已演化到盐湖发展阶段,许多盐湖甚至发展到自析盐阶段[1-3]。根据湖水含盐量的大小,可将新疆湖泊划分为淡水湖、微咸水湖(半咸水湖)、咸水湖和盐湖(卤水湖)四大类型,湖水总盐含量≤1 g/L称为淡水湖,2—34 g/L称为半咸水湖,35—49 g/L称为咸水湖,50 g/L以上即为盐湖[1]。另外新疆湖泊多形成于盆地及风蚀作用形成的丘间洼地,基岩持续风化和淋滤以及地史上古盐的淋溶,对湖泊盐分不断进行补给,不同湖泊成盐母质的差异,造成湖水化学成分的差异,形成了包括碳酸盐、硫酸盐、硼酸盐、氯化物和硝酸盐类多类型蒸发盐类的沉积,特别是硝酸盐类沉积矿物,是新疆盐湖所独有的[1,3]。盐湖作为高盐浓度的特殊环境,含有多种极端嗜盐微生物,盐环境下微生物群落适应高盐离子环境,盐离子对微生物群落的多样性和组成起着重要作用[4],但不同盐湖水化学性质差异是否制约微生物多样性未见报道。因此,研究盐湖水化学性质对微生物菌群结构及演变的影响,同时掌握盐湖微生物种质资源分布规律,对盐湖资源开发利用具有重要意义[5-6]。
新疆不同盐湖中蕴含着丰富的耐盐及嗜盐微生物资源,但针对这一资源的研究较少,早期研究成果多以嗜盐微生物纯培养工作为主[7-8]。近些年采用克隆文库技术对新疆盐湖微生物菌群结构开展了有意义的研究,但克隆文库技术通量较小,仅能获得湖泊主要微生物菌群,无法准确获得微生物群落结构与环境因子互作关系[9-11]。近几年高通量测序技术快速发展,并成功应用于青藏高原盐湖微生物多样性研究[12-13],但利用高通量测序技术研究新疆盐湖微生物多样性研究鲜有报道。
现有研究显示不同湖泊成盐母质的差异,造成湖水化学成分的差异,从而影响湖泊原核微生物菌群结构。此外在盐湖发展过程中,随湖水盐浓度变化,微生物菌群结构动态变化过程无法还原。基于上述研究现状,本文拟解决以下两个主要科学问题,一是新疆天山北坡5个不同盐浓度湖泊湖底沉积物细菌和古菌多样性及菌群结构变化规律及特点。二是新疆天山北坡不同盐湖湖底沉积物菌群结构的主要环境驱动因子。
1 材料与方法
1.1 样品采集
2019年8月分别对新疆天山北坡艾比湖(82°45′48″E,44°55′4″N)、柴窝堡湖(87°53′35″E,43°31′28″N)、盐湖(88°8′17″E,43°23′39″N)、巴里坤湖(92°46′52″E,43°36′26″N)和伊吾湖(94°14′46″E,43°21′47″N)5个湖泊湖底沉积物进行采集,5个盐湖采样位置如图1所示。艾比湖、柴窝堡湖、盐湖、巴里坤湖和伊吾湖后文分别按EB、CWB、YH、Bar和YW简称表示。每个湖泊采集湖水下3份沉积物样本,采集深度为湖底0—20 cm湖底底泥,采集后装入无菌聚丙烯折角袋中均匀混合,每份样品间隔均超过30 m。沉积物样本车载冰箱4℃运载至实验室。每个样本分成两个部分,一部分样品进行理化性质测试,另一部分样品进行总DNA提取进行扩增子测序分析。

图1 采样点位置图Fig.1 Sampling sites
1.2 研究方法
1.2.1湖底沉积物理化因子分析
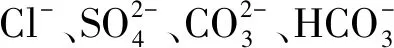
1.2.2湖底沉积物基因组DNA提取
采用FastDNA® SPIN Kit for Soil 土壤基因组DNA提取试剂盒对样本中基因组DNA进行提取,操作步骤按照试剂盒说明进行。基因组DNA利用琼脂糖凝胶检测DNA的长度和完整性,利用NanoDrop 2000检测DNA的浓度和纯度,符合要求后委托诺禾致源进行扩增子测序分析。
1.2.3PCR扩增及扩增子测序
基于Ion S5TMXL测序平台,利用单端测序的方法对样本中细菌和古菌进行扩增子测序分析。细菌为16S V4—V5区,测序引物为515f/926r[15]。古菌为16S V4区,测序引物为Arch519F/Arch915R[16-17]。使用带10 bp Barcode序列的简并引物对不同的样本进行扩增,根据PCR产物浓度进行等量混样,混匀后样本进行琼脂糖胶电泳,切割目标条带,使用Thermo Fisher公司GeneJET胶回收试剂盒对产物进行回收纯化。纯化样本使用Thermo Fisher公司的Ion Plus Fragment Library Kit试剂盒进行文库构建,构建好的文库经过Qubit定量和文库检测合格后,进行测序分析。柴窝堡湖、盐湖、艾比湖、巴里坤湖和伊吾湖细菌注册号为:PRJNA662635;古菌注册号为:PRJNA662632。
1.2.4不同盐湖原核微生物菌群结构分析
测序原始数据使用Cutadapt软件[18]对低质量序列进行剪切,根据Barcode序列拆分不同样本,并截去Barcode和引物序列。再使用vsearch[19]软件将测序数据与物种注释数据库进行比对去除嵌合体序列获得有效数据用于后续分析。利用Uparse[20]软件对所有样品有效数据按照97%进行聚类,聚类生成OTUs(Operational Taxonomic Units)数据集,筛选数据集中出现频数最高的序列作为代表序列。代表序列采用Mothur软件与SILVA数据库[21]进行物种注释,在界、门、纲、目、科、属不同分类水平上进行分析。
1.2.5数据分析
用R语言绘制环境因子和Alpha多样性箱线图,门和纲水平百分比堆积图,Tukey HSD检验(multcomp包)进行多重比较,添加显著性(P≤0.05)标记;pheatmap函数(pheatmap包)绘制属水平热图;基于UPGMA聚类和Unweighted Unifrac距离进行PCoA(principal co-ordinates analysis)分析;使用环境因子和门水平丰度数据进行RDA(redundancy analysis)分析;通过Bioenv函数(vegan包)分析影响盐湖微生物群落结构多样性的环境因子组合,并进行mantel检验。
2 结果与分析
2.1 不同盐湖湖底沉积物理化性质


图2 五个盐湖湖底沉积物理化因子(n=3)Fig.2 Sediment properties at the bottom of five salt lakes (n=3)
2.2 不同盐湖湖底沉积物原核微生物多样性及群落结构
2.2.1测序数据分析
城际铁路具有高效、快捷、舒适、大运量等特点,在加强区域内城市间有机联系和促进区域经济一体化的实现中扮演着越来越重要的角色。然而,城际铁路建设项目投资金额大、建设周期长、技术要求高,且具有明显的区域依赖性,依靠传统的融资模式进行大规模城际铁路建设难以为继。因此,积极拓宽城际铁路建设融资渠道,提高对社会资本的吸引强度,成为促进铁路可持续健康发展和城际交通网络加速形成的关键。
5个盐湖湖底沉积物细菌和古菌16S rRNA扩增子测序原始数据每个样本为8万条左右,经质控,聚类及物种注释,绘制稀释曲线。从稀有度曲线来看,所有样本在测序范围内均趋于饱和,表明测序量合理,能够比较真实地反映沉积物中原核微生物群落结构。细菌和古菌测序引物均扩增出了细菌和古菌两界微生物,细菌测序结果中古菌OTUs比例较低,但古菌测序结果中细菌OTUs比例较高,为了更好的研究湖底沉积物中细菌和古菌菌群结构变化趋势,在分析时剔除相应古菌和细菌OTUs,抽平后进行分析。另外细菌和古菌测序覆盖率均高于99%,表明测序深度能代表样本的真实情况。
2.2.2沉积物中原核微生物多样性
Alpha多样性计算结果显示(图3),5个盐湖湖底沉积物中细菌和古菌的多样性指数随总盐浓度的变化趋势不同。细菌的6个多样性指数与湖底沉积物总盐浓度呈相反的变化趋势(巴里坤湖除外),总盐浓度增大则细菌多样性下降;古菌的6个多样性指数在总盐浓度最高或最低时多样性更大。伊吾湖湖底沉积物细菌的Alpha多样性最低,多个丰富度指数显示其细菌群落丰度较低,Simpson指数显示其细菌菌群结构不均匀,PD(PD whole tree)指数显示其菌群进化差异最小;古菌的群落丰度和进化差异和细菌一致,Simpson指数显示其古菌菌群结构比较均匀。巴里坤湖细菌多个丰富度指数和PD指数较低,Simpson指数反而高,显示其细菌群落丰富度和进化差异尽管低,但是群落结构比较均匀;古菌的Alpha多样性指数皆较高,显示其不仅古菌群落丰富度和进化差异较大,而且群落结构均匀。柴窝堡湖细菌Alpha多样性指数最高,古菌Alpha多样性指数都较高,显示其细菌和古菌群落丰富度和进化差异较大,群落结构均匀。此外5个湖泊细菌richness、chao1和ACE丰富度指数远高于古菌,且细菌丰富度指数差异较大,古菌多样性指数差异相对较小。
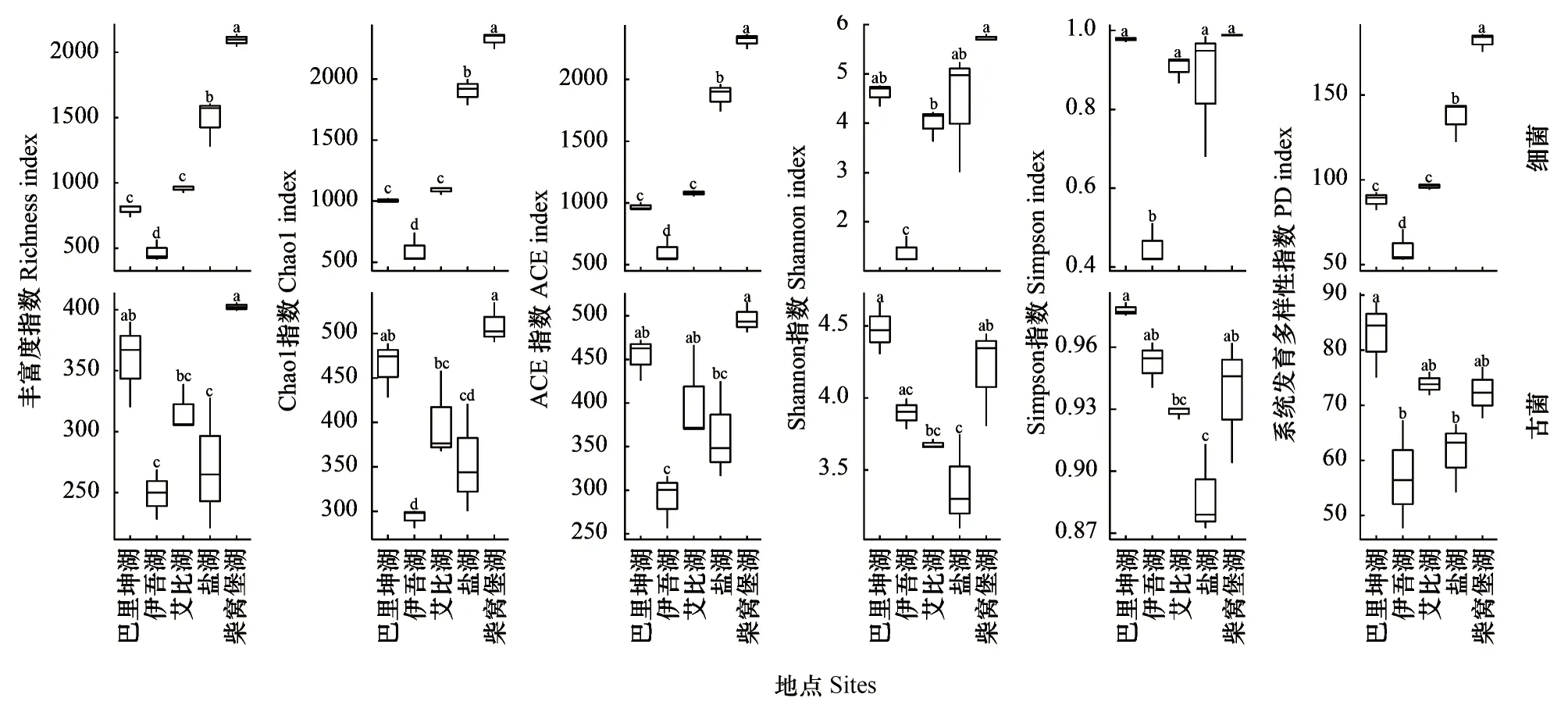
图3 5个盐湖湖底沉积中细菌和古菌α多样性指数(n=3)Fig.3 Alpha-diversity indices of bacterial and archaeal community of five salt lakes sediment (n=3)ACE:基于丰度的覆盖估计值 Abundance-based coverage estimator;PD:系统发育多样性Phylogenetic diversity
2.2.3沉积物中原核微生物群落组成和结构
将5个盐湖湖底沉积物样本中的OTUs代表性序列在SILVA数据库中进行比对,在门、纲、目、科、属不同的分类水平进行统计,共检测获得细菌58门、68纲、138目、253科和560属;古菌4门、8纲、12目、21科和60属。如图4所示,5个盐湖细菌在门水平上相对丰度排名前10的分别为变形菌门(Proteobacteria)、蓝藻门(Cyanobacteria)、拟杆菌门(Bacteroidetes)、芽单胞菌门(Gemmatimonadetes)、厚壁菌门(Firmicutes)、绿弯菌门(Chloroflexi)、未鉴定细菌(unidentified-bacteria)、放线菌门(Actinobacteria)、异常球菌-栖热菌门(Deinococcus-Thermus)和酸杆菌门(Acidobacteria)。其中变形菌门为主要类群,在伊吾湖相对丰度达92%,在盐湖相对丰度不足25%,在不同盐浓度的湖泊当中所占比例差异较大。变形菌门共包含γ-变形菌纲(Gammaproteobacteria)、Δ-变形菌纲(Deltaproteobacteria)和α-变形菌纲(Alphaproteobacteria)3个纲,γ-变形菌纲所占比例最大。除变形菌门以外,拟杆菌门和蓝藻门相对丰度在不同盐湖当中差别不大。从结果中可以看出,不同盐湖细菌丰度较高前10个门的相对丰度变化并非仅随总盐浓度高低发生规律性变化。
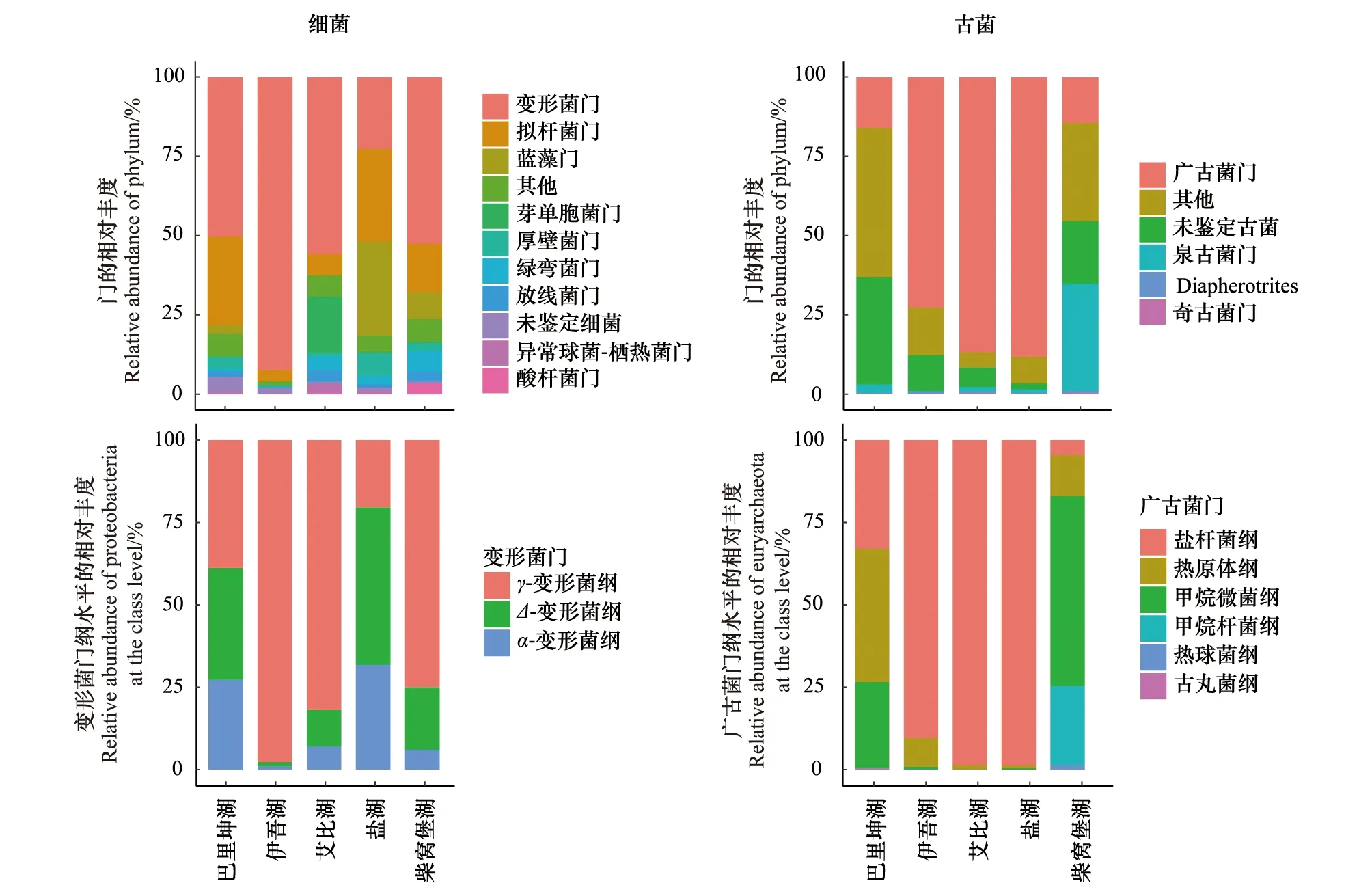
图4 5个盐湖湖底沉积中微生物相对丰度 Fig.4 The relative abundance of microbial community in five salt lakes sediment
5个盐湖古菌在门水平进行分类,结果为广古菌门(Euryarchaeota)、Othes、未鉴定古菌(unidentified-Archaea)、泉古菌门(Crenarchaeota)、Diapherotrites和奇古菌门(Thaumarchaeota)(图4)。从5个盐湖湖底沉积物中鉴定出4个门的古菌,其中广古菌门为盐湖、艾比湖和伊吾湖古菌主要类群,泉古菌门为柴窝堡湖古菌主要类群,巴里坤湖多以未鉴定的古菌和其他门类为主。5个湖泊相对丰度最高广古菌门共包含盐杆菌纲(Halobacteria)、热原体纲(Thermoplasmata)、甲烷微菌纲(Methanomicrobia)、甲烷杆菌纲(Methanobacteria)、热球菌纲(Thermococci)和古丸菌纲(Archaeoglobi)7个古菌纲,其中Halobacteria纲占有很高的比例(图4)。从结果中同样可以得出,不同盐湖古菌丰度较高门的相对丰度也不随总盐浓度高低发生规律性变化,不考虑总盐浓度最低的柴窝堡湖的话,随总盐浓度增大,广古菌门所占比例在降低,未鉴定的古菌门所占比例在增大。
根据5个盐湖湖底沉积物样本在属水平的物种注释及丰度信息,选取丰度排名前35的属,在样本和物种两个层面进行聚类分析,分别绘制细菌和古菌属水平聚类热图。湖底沉积物样本中细菌属水平菌群结构聚类热图如图5所示,丰度前35个属分别属于细菌的6个门,其中放线菌门包含1个属、拟杆菌门9个属、蓝藻门1个属、异常球菌-栖热菌门1个属、厚壁菌门3个属、变形菌门19个属和未鉴定细菌1个属。古菌属水平聚类热图如图5所示,丰度前35个属分别属于古菌的3个门,泉古菌门2个属、Diapherotrites 1个属、广古菌门29个属、未鉴定古菌1个属和Unassigned 2个属。聚类结果还显示不同盐湖均出现了相应的微生物菌群,不同盐湖细菌和古菌优势属种类均不相同,有其独特菌群结构类型。指示物种分析(Indicator Species Analysis)显示5个湖泊优势细菌属数量大小为:巴里坤湖>盐湖>艾比湖>柴窝堡湖>伊吾湖;5个湖泊优势古菌属数量大小为:伊吾湖>艾比湖>巴里坤湖>盐湖=柴窝堡湖。其中伊吾湖的优势细菌属只有1个(Stenotrophomonas),而古菌优势属有8个。巴里坤湖的优势细菌属有Halothiobacillus、Desulfosalsimonas、Desulfovermiculus、Thioalkalivibrio、Tangfeifania、Dethiosulfatibacter、Roseovarius、Desulfonatronobacter和Desulfotignum,优势细菌属均为嗜盐微生物属,其中一些属与硫元素代谢相关[23]。巴里坤湖的优势古菌属有Methanolobus和Methanohalophilus,与甲烷代谢相关[24]。伊吾湖古菌中Halolamina、Halorubellus、Halapricum、Haloplanus、Halodesulfurarchaeum、Natronoarchaeum和Haloquadratum属占优势,也均是一些嗜盐古菌属。盐浓度高的巴里坤湖拥有更多的优势细菌属,盐浓度高的伊吾湖拥有更多的优势古菌属,盐浓度低的柴窝堡湖拥有的优势菌属较少。Simachew等[25]在盐浓度梯度增加时的研究显示两个最丰富的OTU在属水平上分别分为Halorubrum和Halorhabdus,它们都属于古细菌嗜盐菌科。巴丹吉林沙漠盐湖沉积物中属水平上高丰度聚类分支主要集中在盐单胞菌科(Syntrophomonadaceae),超高盐度沉积物中的优势菌属是norank_Syntrophomonadaceae,该菌属属于盐单胞菌科(Syntrophomonadaceae)[26]。可以推断,随着盐湖盐浓度的升高,微生物群落会由不耐盐的微生物群落向耐盐或嗜盐微生物群落进行演化。
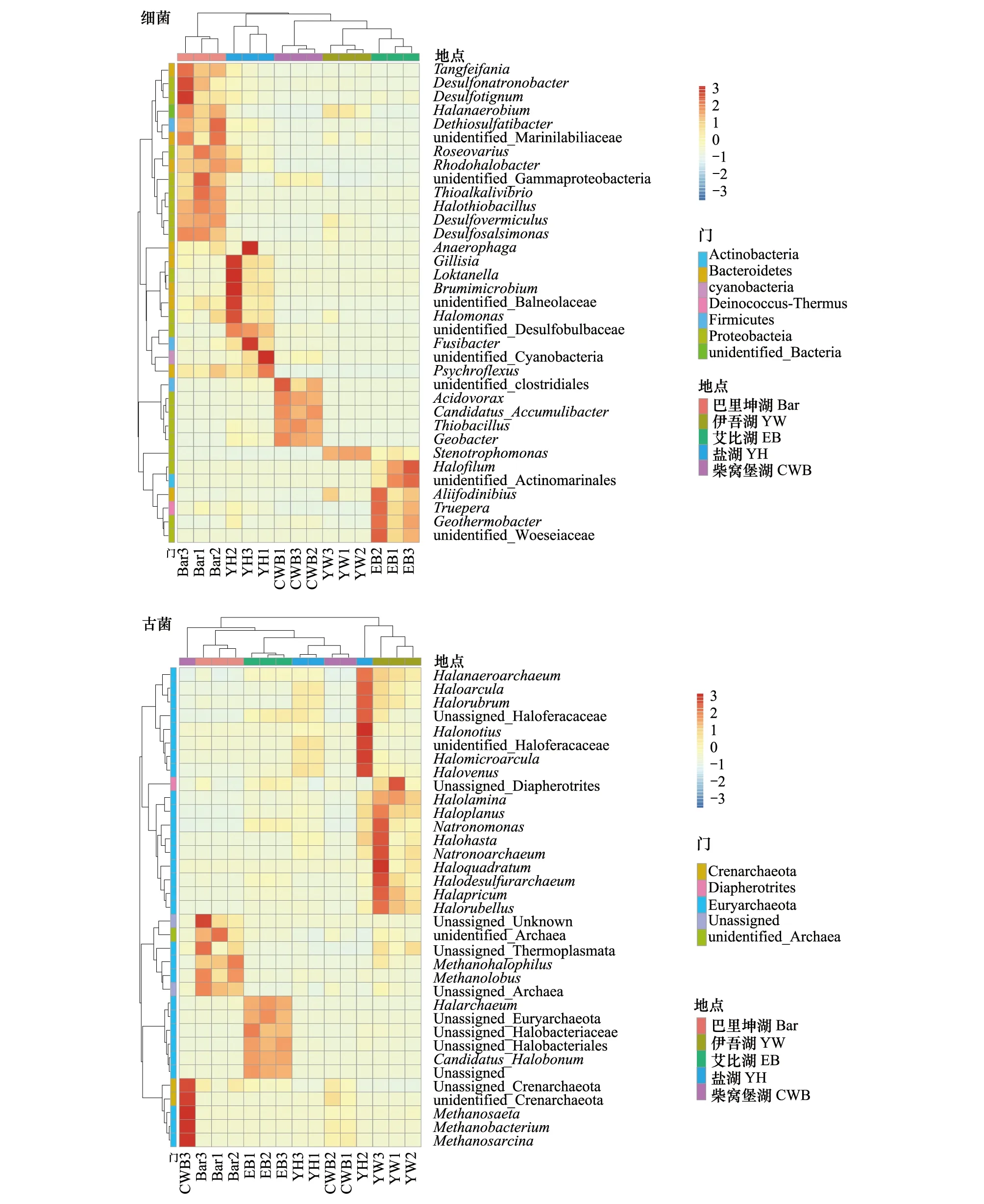
图5 5个盐湖湖底沉积中微生物属水平相对丰度聚类热图Fig.5 Top 35 genera heatmaps of microbial community in five salt lakes sediment 属图例(自上而下、自左至右,仅列出有对应中文名称的属):盐厌氧菌属Halanaerobium;玫瑰杆菌属 Roseovarius;硫碱弧菌属 Thioalkalivibrio;盐硫杆菌属 Halothiobacillus;盐单胞菌属 Halomonas;小纺锤状菌属 Fusibacter;冷弯菌属 Psychroflexus;嗜酸菌属 Acidovorax;硫杆菌属 Thiobacillus;土杆菌属 Geobacter;寡养单胞菌属 Stenotrophomonas;特吕珀菌属 Truepera;盐盒菌属 Haloarcula;盐红菌属 Halorubrum;盐薄片菌属 Halolamina;盐盘菌属 Haloplanus;盐碱单孢菌属 Natronomonas;盐方菌属 Haloquadratum;甲烷嗜盐菌属 Methanohalophilus;甲烷叶菌属 Methanolobus;甲烷鬃菌属 Methanosaeta;甲烷杆菌属 Methanobacterium;甲烷八叠球菌属 Methanosarcina
2.2.4不同盐湖原核微生物群落构成比较分析
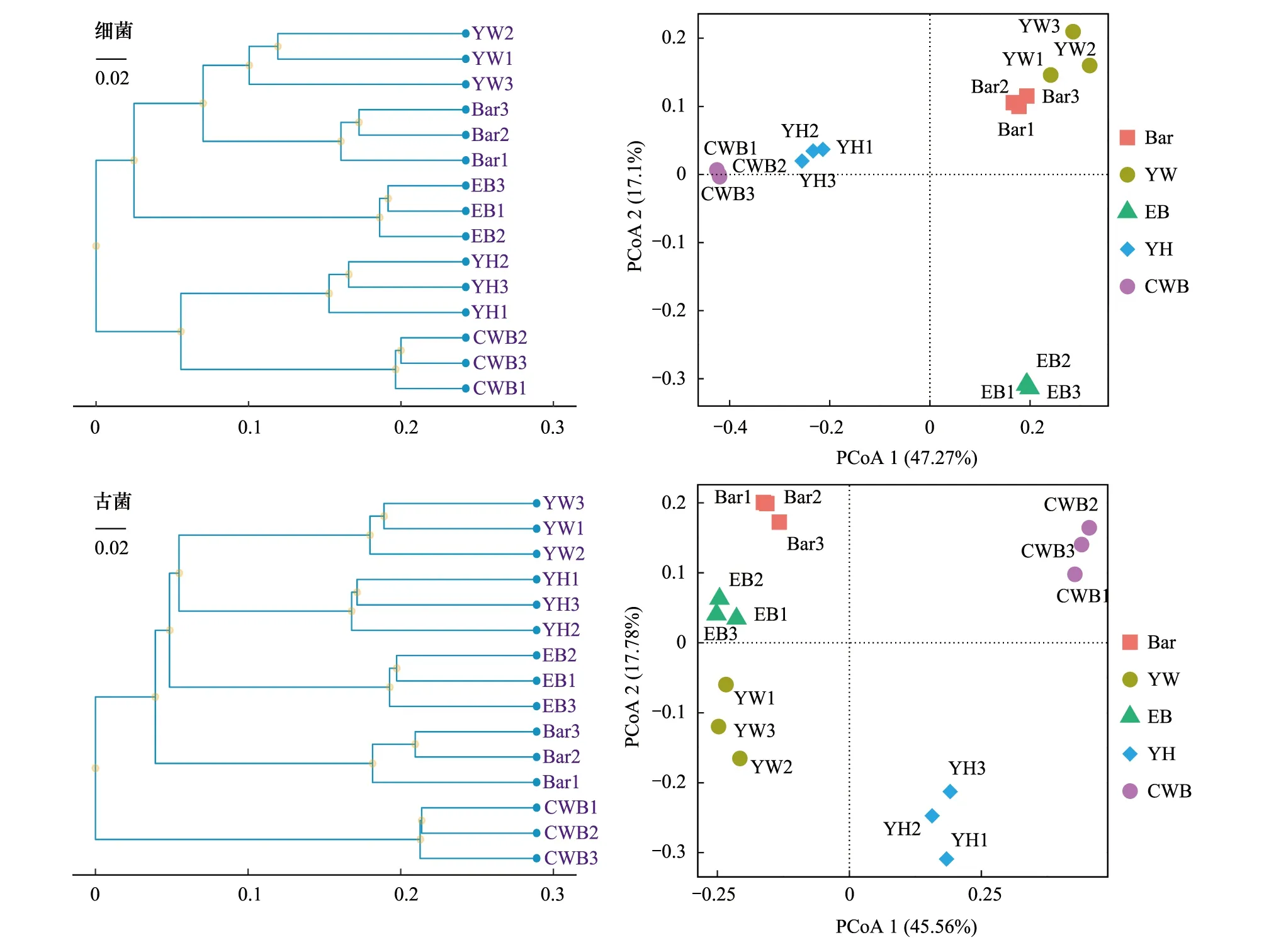
图6 5个盐湖湖底沉积物中微生物OTUs UPGMA聚类和主坐标分析 Fig.6 UPGMA cluster and principal co-ordinates analysis of microbial community at OTU levels in five salt lakes sediment
2.3 不同盐湖原核微生物多样性与环境因子相关性分析
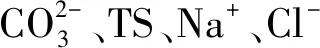
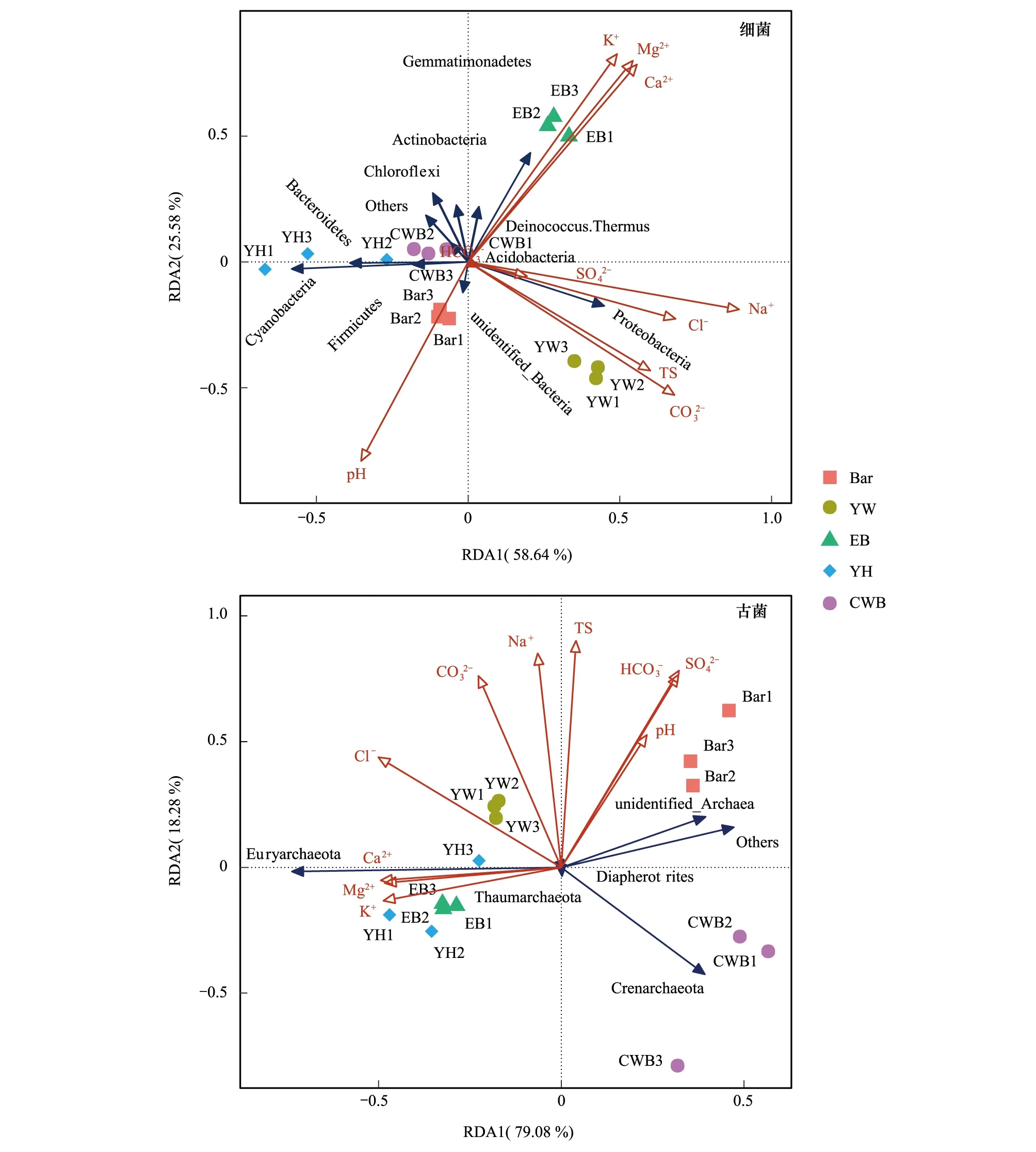
图7 5个盐湖湖底沉积物中原核微生物种类与理化因子相关性分析Fig.7 Correlation analysis of prokaryotic microorganisms and chemical properties in five salt lakes sediment
2.4 环境因子对盐湖湖底沉积物菌群结构的影响

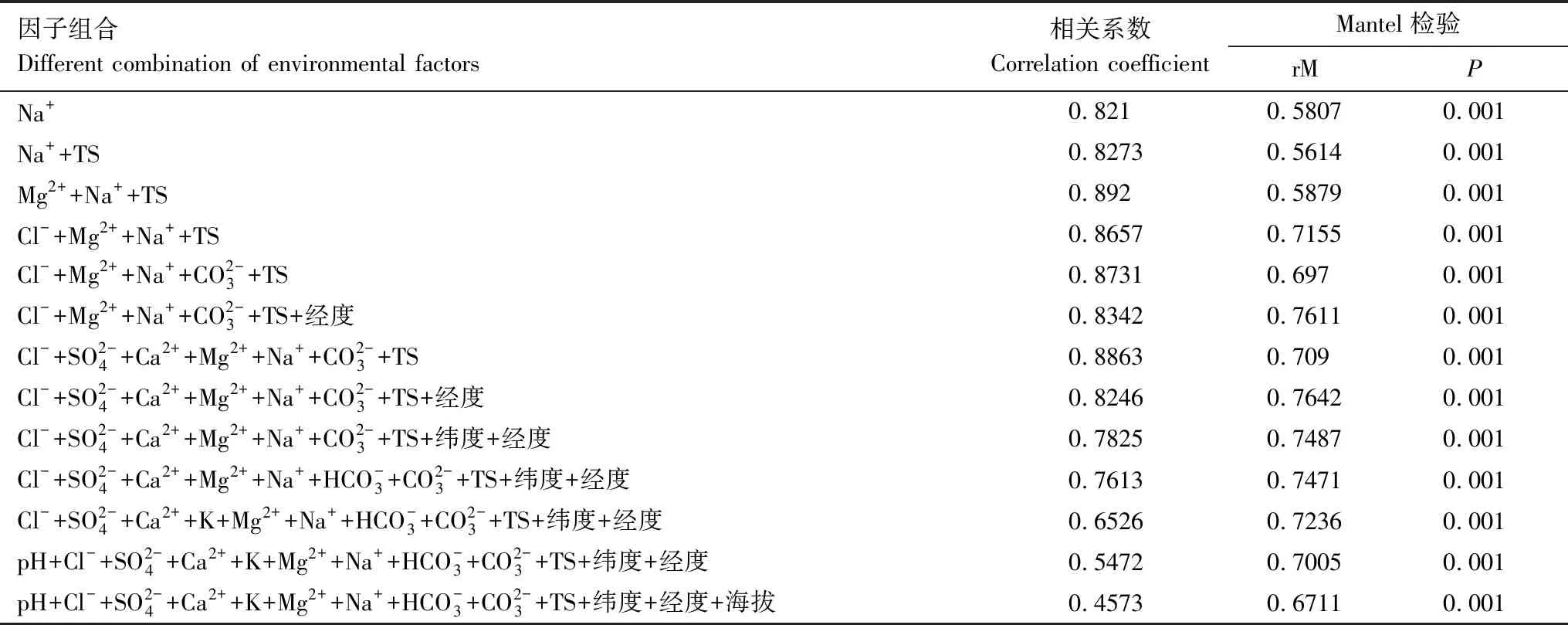
表1 不同环境因子组合与细菌群落结构的相关关系Table 1 The relationship between different combination of environmental variables and bacterial community

表2 不同环境因子组合与古菌群落结构的相关关系Table 2 The relationship between different combination of environmental variables and Archaeal community
3 讨论
分子生态学研究能够很好的揭示不同地域、不同湖泊现有演化阶段条件下微生物群落特征,但很难针对一个特定盐湖,研究它在历史进化(咸化或淡化)过程中微生物群落结构的变化。2014年Izhitskiy等[27]采集了咸海不同湖泊样本进行分析,发现不同湖泊虽起源相同,但水流补给等因素不同,导致不同湖泊盐度存在较大差异,微生物群落结构也表现出不同的特征。Shurigin等[28]也发现大咸海中具有独特的群落结构。Simachew等[25]在研究苏打浓缩池中盐浓度增加时原核微生物多样性时发现随着盐度的增加,观察到从细菌到古菌的显着群落转移。目前的研究代表了时间和空间上的“快照”,微生物的多样性可能会随着季节和持续的干旱或降雨而改变[28],无法经历湖泊整个进化过程,所以只有通过现代湖泊微生物群落结构的研究推演盐湖进化过程中微生物菌落结构变化规律[29]。因此,本文采用高通量测序技术研究新疆天山北坡5个不同盐浓度湖泊湖底沉积物微生物菌群结构,从而推演新疆天山北坡盐湖进化过程中微生物菌群结构演变成因。
3.1 盐湖湖底沉积物微生物多样性与结构
天山以北的准噶尔盆地,地形向西北开口,未完全封闭,使得来自大西洋的盛行西风和来自北冰洋的气流,能够顺着山口进入,向南碰到天山北坡抬升,从而形成地形雨,产生降水。天山北坡的降水量要多于天山南坡,使得天山北坡的水分条件比南坡要好。此外天山北坡中山带生长有针叶森林,针叶森林涵养了大量水源,也正因如此天山北坡形成较多的天然湖泊。实验选取的天山北坡5个盐湖细菌在门水平上相对丰度依次为变形菌门、蓝藻门、拟杆菌门、芽单胞菌门、厚壁菌门、绿弯菌门、放线菌门、异常球菌-栖热菌门以及酸杆菌门,其中变形菌门为主要类群。张彦茹[30]采用克隆文库法研究内蒙古浑善达克地区桑根达来、达各淖和扎格斯台淖三个盐碱湖湖水细菌群落结构时发现,桑根达来盐碱湖细菌群落主要以变形菌门为主,达各淖盐湖主要以蓝藻门为主,扎格斯台淖盐湖主要以放线菌门为主。Ben等[31]采用高通量扩增子测序的方法研究突尼斯南部杰里德大盐湖雨季湖底沉积物微生物菌群结构时,结果显示沉积物中细菌群落以变形菌门为主。多个研究发现,盐湖细菌类群中多以变形菌门、厚壁菌门、拟杆菌门和酸杆菌门为主[32],但因各个盐湖理化成分差异,造成优势类群比例方面有所不同。将本研究5个盐湖与青藏高原茶卡盐湖和小柴旦盐湖湖底沉积物中细菌菌群结构研究结果进行比对,青藏高原两个盐湖海拔分别为3017 m和3171 m,厚壁菌门为主要细菌类群,因此推测除理化成分影响外,海拔也会对菌群结构造成影响。
5个盐湖古菌在门水平上相对丰度依次为广古菌门、泉古菌门、Diapherotrites和奇古菌门。张欣等[12]在研究茶卡盐湖湖底沉积物古菌类群时发现,古菌主要属于广古菌门(96.5%)、盐纳古菌门(1.13%)和未确定分类地位的古菌门(2.16%)。刘静等[13]在研究小柴旦盐湖湖底沉积物中古菌群落结构时,主要类群为沃斯古菌门(80.84%)、广古菌门(7.55%)、奇古菌门(0.95%)和其他(0.67%)。此外,邵冠军[33]对艾比湖湖底沉积物的古菌多样性进行研究,发现古菌类群多集中在广古菌门(96.6%),其次为盐纳古菌门(1.1%)。而本次研究5个湖泊中均未扩增出盐纳古菌门,分析发现进行研究所采用的简并引物不同,会造成研究结果的不同,甚至会导致研究结果物种相对丰度有所差异[34]。此外对5个盐湖的古菌Alpha多样性指数比较发现,richness、ACE和chao1丰富度指数不像细菌对应丰富度指数那样存在很大的差异,这与孙超[35]研究新疆玛纳斯盐湖和阿洪库鲁木湖原核微生物多样性类似。5个盐湖中细菌的多样性远高于古菌,与赵婉雨[36]在达布逊盐湖和刘静[13]在小柴旦盐湖的研究一致。
3.2 5个盐湖湖泊菌群结构主要驱动因子

4 结论
新疆湖泊受干旱气候等因素的影响,成盐作用强烈,加之近半个世纪湖泊上游农业引水灌溉,加剧了盐湖的演化过程。盐湖在演化发展过程中,极端嗜盐菌可能会在低盐离子浓度环境中存活,但可能不占据优势生长地位,从而造成在不同环境中种类的差异[49]。因此本研究结果显示新疆天山北坡盐湖湖底沉积物中细菌群落结构主要受Na+和TS浓度影响,古菌群落结构主要受Na+和TS浓度影响外,还受其他理化因子共同影响,5个盐湖均演化出了相应的微生物菌群,不同盐湖细菌和古菌优势属种类均不相同,有其独特菌群结构类型。其中盐湖中特殊卤水成分会对微生物群落结构产生重大影响,从而孕育特殊的微生物资源,对开发利用极端微生物资源提供参考。
猜你喜欢
东坡赤壁诗词(2023年1期)2023-05-30 10:48:04
土壤学报(2022年3期)2022-08-26 12:15:26
大自然探索(2022年5期)2022-07-11 03:10:33
草业学报(2022年5期)2022-07-08 06:24:40
知识就是力量(2022年6期)2022-06-16 20:19:36
领导文萃(2021年19期)2021-11-05 16:39:10
草业学报(2021年3期)2021-03-22 02:22:00
草业学报(2018年10期)2018-10-19 05:25:44
电镀与环保(2017年5期)2017-12-19 12:06:03
诗潮(2016年1期)2016-12-08 11:06:36
