
LncRNA SNHG17通过诱导上皮间质转化促进结直肠癌转移
2021-10-11 01:12耿长辉张春玲高萍何苗李慧峰姚旖旎刘颖新
中华结直肠疾病电子杂志 2021年4期
耿长辉 张春玲 高萍 何苗 李慧峰 姚旖旎 刘颖新
结直肠癌是人类最常见、最具侵袭性的恶性肿瘤之一,是全球第三大癌症致死原因[1-2]。虽然近些年来手术技术及药物治疗技术不断进步,但结直肠癌患者的复发率及死亡率还是居高不下。因此,研究结直肠癌发展过程中肿瘤发生的分子机制,寻找新的有效治疗靶点,对于提高患者生存率至关重要。
长链非编码RNA是近十几年来发现的长度超过200个核苷酸的非编码转录本,没有完整的功能性开放阅读框[3-5]。近年来,其对细胞生物学行为的影响,尤其是对肿瘤细胞的影响,已逐渐引起人们的重视。越来越多的研究表明,lncRNA在肿瘤中异常表达,参与了肿瘤的发生与发展的过程[6-9]。
Small nucleolar RNA host gene 17(SNHG17)是lncRNA的成员之一。它位于第20号染色体q11.23上,长1 186 nt。最新证据表明,SNHG17与多种肿瘤的发生有关,包括骨肉瘤、卵巢癌、口腔鳞癌、前列腺癌和脑胶质瘤等[10-14]。然而,它们在结直肠癌发生与发展中的具体作用仍然知之甚少。在这项研究中,我们旨在探究lncRNA SNHG17在结直肠癌细胞系和组织中的表达情况及其对结直肠癌的调控作用,争取为结直肠癌治疗寻找一个新的靶点。
材料与方法
一、细胞系
人正常结肠上皮细胞系NCM460和人结肠癌细胞(HCT116,HT29,SW480,SW620)购自the American Type Culture Collection(ATCC,马纳萨斯,美国)。人结肠癌细胞系LOVO购自上海生物科学研究所(上海,中国)。HCT116和SW480在添加10%胎牛血清(GIBCO,Carlsbad,CA)的McCOY's 5a培养基(江苏凯基生物科技有限公司,中国)和Dulbecco's modified Eagle's medium(DMEM)培养基(GIBCO,美国)中培养。所有细胞在37°C的含5%CO2湿培养箱内培养。
二、细胞转染
过表达病毒和Lenti-shSNHG17及其相应的对照载体是从吉凯公司(上海,中国)购买的。转染后,利用嘌呤霉素(Sigma,美国)进行筛选传代获得稳转细胞株。shRNA序列如下:(shNC:5'-TTCTCCGAACGTGTCACGT-3'; shSNHG17-1: 5'-GAUUGUCAGCUGACCUCUGUCCUGU-3';shSNHG17-2:5'-CGUGUCUUCAAGAAGGGCUGAGCU-3')。
三、RNA提取和qRT-PCR实验
用TRIzol试剂(Invitrogen,美国)从结直肠癌细胞系中提取总RNA,并根据生产厂家的说明,用高容量cDNA逆转录试剂盒(Applied Biosystems, CA) 逆 转 录 2 μg 总 RNA。 用Nanodrop 2000光谱仪(Thermo Fisher Scientific)测定DNA浓度。定量PCR用0.8 μg cDNA与SYBR Green试剂(Thermo Fisher Scientific,USA)应用Biosystem 7500qPCR系统(Applied Biosystems)进行qPCR扩增。以β-actin为内源基因,用2-ΔΔCT方法比较相对表达。引物序列如下:
SNHG17-F:5'-GTTCCTGGGGCTTGGATGAT-3'SNHG17-R:5'-GATCTAAGGCTGAGACCCACG-3'GAPDH-F:5'-AGCCACATCGCTCAGACAC-3'GAPDH-R:5'-GCCCAATACGACCAAATCC3'
四、Transwell侵袭实验
在小室上层铺入基质胶后,于孵箱静置4小时后,在小室上层中加入200 μL无血清培养基,并接种2×105个结直肠癌细胞,下层中加入600 μL的完全培养基。培养24~48小时后,用多聚甲醛固定下层细胞15分钟,并用0.1%的结晶紫染色。用Nikon显微镜拍照,用Image J软件计算细胞数。
五、划痕实验
在六孔板中接种2×105个结直肠癌细胞,待细胞长到100%密度的时候,用200 μL的枪头进行划线。用PBS洗涤细胞碎片及悬浮死细胞后分别于0小时和48小时用显微镜拍照,用Image J软件计算面积。
六、Western blot实验和抗体
采用蛋白质印迹法在10%的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)上分离蛋白质,然后将其转移到硝酸纤维素膜上。将印迹与抗体在4°C下孵育过夜。洗涤三次后,将膜与二抗在室温下孵育2小时。蛋白条带通过ECL(北京,中国)可视化。
抗体使用及来源如下:Vimentin(Cell Signaling Technology# 5741), N-Cadherin(Cell Signaling Technology##13116),E-Cadherin(Cell Signaling Technology##3195),GAPDH(ZSGB-Bio#TA-8)。
七、数据库
GEPIA(http://gepia.cancer-pku.cn/)是一种癌症表达谱数据的交互式web服务器。GEPIA包含肿瘤/正常组织的差异表达分析、不同癌症类型或病理阶段的分析、生存分析及相关性分析。本研究利用了GEPIA的差异表达分析分析了数据库中结直肠癌患者的肿瘤组织及正常组织的SNHG17的表达水平。
八、统计学方法
所有统计分析均采用SPSS 23.0软件进行分析(IBM)。数据表示为至少三个独立实验的平均标准偏差。两组之间采用t检验或方差分析。Prism software version 8(GraphPad软件)用于绘图。P<0.05表示差异有统计学意义。
结 果
一、SNHG17在结直肠癌细胞系及组织中高表达
首先,利用GEPIA数据库分析了SNHG17在结直肠癌患者正常组织和肿瘤组织中的表达,发现其在肿瘤组织中的表达要明显上调(图1A)。随后提取了结肠癌细胞系和正常结肠上皮细胞的RNA,通过逆转录及qRT-PCR发现lncRNA SNHG17在结直肠癌细胞系中表达也明显增高(图1B)。而后分别将表达水平最低的SW480细胞进行SNHG17的过表达(图1C),表达水平最高的HCT116细胞进行SNHG17的敲降(图1D),进行后续的功能实验。
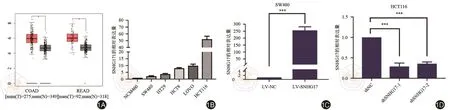
图1 LncRNA SNHG17在结直肠癌中的表达情况及稳转细胞系的构建。1A:结直肠癌组织中SNHG17的表达水平;1B:qRT-PCR分析各结肠癌细胞系SNHG17的表达水平;1C,1D:qRT-PCR检测慢病毒转染构建稳转细胞系SNHG17表达水平。P<0.05表示为*;P<0.01代表为**;P<0.001表示为***
二、SNHG17促进了结直肠癌细胞的迁移与侵袭
进行划痕实验并进行分析,发现过表达SNHG17后细胞的迁移能力明显增加(图2A),敲降SNHG17后细胞的迁移能力明显下降(图2B)。Transwell侵袭实验也表明随着SNHG17的表达增加,其侵袭能力明显增强(图2C)。当敲降SNHG17后,其侵袭能力明显下降(图2D)。
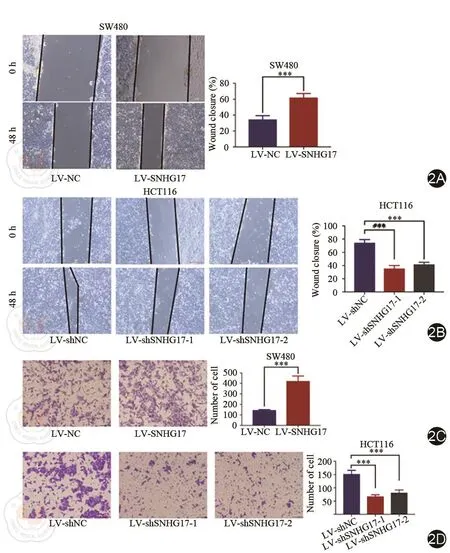
图2 LncRNA SNHG17在结直肠癌中的作用。2A,2B:划痕实验验证各细胞系迁移能力;2C,2D:Transwell实验验证各细胞系侵袭能力。P<0.05表示为*;P<0.01代表为**;P<0.001表示为***
三、SNHG17诱导结直肠癌细胞发生上皮间质转化
上皮间质转化是肿瘤细胞转移能力的重要原因之一。为探讨SNHG17促进结直肠癌细胞转移的机制,我们进行western blot实验,发现当SNHG17表达增加的时候,间质细胞标志物N-cadherin和Vementin的表达水平明显增加,上皮标志物E-cadherin的表达水平明显下降(图3A)。我们还发现当敲降SNHG17的时候,间质细胞标志物N-cadherin和Vementin的表达水平明显减少,上皮细胞标志物E-cadherin的表达水平明显上升(图3B)。SNHG17促进了结肠癌细胞的上皮间质转化。
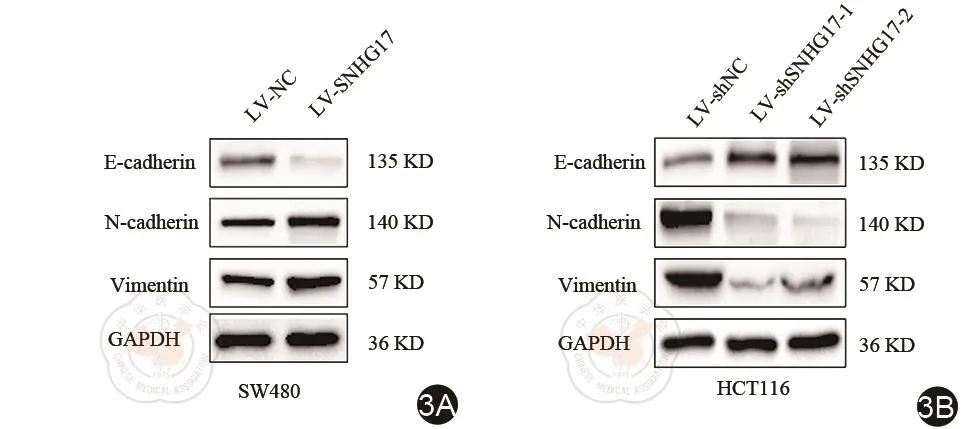
图3 SNHG17促进了结直肠癌细胞上皮间质转化。3A,3B:Western blot实验分析各细胞系蛋白表达水平
讨 论
结直肠癌因其高发病率和高死亡率一直是癌症工作者研究的热点话题。在结直肠癌发生与发展中的关键分子及机制有望提升患者的治疗与预后价值。近些年来,长链非编码RNA在结直肠癌中发生的作用也越来越多。有学者发现了m6A诱导的lncRNA GAS5对YAP信号的衰减在结直肠癌进展中的新机制可能为CRC治疗提供了一种有前景的途径[15]。还有研究表明lncRNA GLCC1在糖饥饿下在CRC细胞中显著上调,通过与HSP90伴侣直接相互作用稳定c-Myc转录因子的泛素化,并进一步指定c-Myc靶基因(如LDHA)的转录修饰模式,从而促进糖酵解来支持细胞生存和增殖[16]。另外,m6A诱导的lncRNA RP11通过上调Zeb1促进结直肠癌细胞的扩散[17]。然而lncRNA SNHG17在结直肠癌转移中的机制还鲜有报道。我们前期通过TCGA数据库分析发现lncRNA SNHG17在结直肠癌肿瘤组织中高表达,并且通过qRT-PCR证明其在结直肠癌细胞株中表达也上调了。而后我们进行功能实验,通过敲降和过表达SNHG17证明了lncRNA SNHG17可以促进结直肠癌细胞的迁移和侵袭。上皮间质转化指上皮到间质细胞的转化,它赋予细胞转移和侵袭的能力,包括干细胞特征、减少凋亡与衰老和促进免疫抑制,不仅在发育过程中起着关键的作用,而且还参与组织愈合、器官纤维化和癌症发生等过程[18-20]。为了探究SNHG17与上皮间质转化之间的关系,我们通过western blot实验验证了lncRNA SNHG17促进了间质细胞标志物N-cadherin和Vementin的表达,抑制了上皮细胞标志物E-cadherin的表达,诱导了上皮间质转化的发生。以上研究表明lncRNA SNHG17通过诱导上皮间质转化促进了结直肠癌细胞的迁移及侵袭。
本研究中尚有未解的科学问题:(1)lncRNA SNHG17在结直肠癌中表达增高的机制或者原因是什么;(2)lncRNA SNHG17通过什么分子机制诱导了上皮间质转化:由于lncRNA无法编码蛋白,故其产生的生物学功能最终的效应一定是改变了下游的相应蛋白,可以进一步去研究lncRNA SNHG17通过何种靶蛋白进一步影响了上皮间质转化;(3)lncRNA在细胞和组织内的定位如何:如果是主要在细胞核中表达,它能否通过调控染色质层面或者组蛋白修饰水平等调控下游基因的表达;如果主要在细胞质中表达,是否调控蛋白质的翻译或翻译后修饰,是否通过ceRNA机制调控下游靶基因的表达。以上三个问题都有待进一步的研究去阐明。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国现代医生(2022年21期)2022-08-22
中国典型病例大全(2022年7期)2022-04-22
健康体检与管理(2022年2期)2022-04-15
科学与生活(2021年16期)2021-11-25
青岛大学学报(医学版)(2021年5期)2021-11-17
天津医科大学学报(2021年2期)2021-03-29
饮食科学(2016年7期)2016-07-27
饮食与健康·下旬刊(2016年9期)2016-05-14
中国实用医药(2009年5期)2009-05-08
