
不同水盐条件对三江藨草生长和生物量分配的影响
2021-10-09 01:52张超凡潘丽铭姚明远李伟东马琼芳
生态学报 2021年16期
张超凡,燕 红,潘丽铭,姚明远,刘 庚,李伟东,齐 巍,马琼芳,*
1 吉林省林业科学研究院, 长春 130033 2 长白山湿地与生态吉林省联合重点实验室, 长春 130102 3 吉林龙湾国家级自然保护区管理局, 通化 135102
湿地是连接陆地生态系统与水生生态系统之间的过渡带,其独特的生态环境在物种基因保护、稳定全球气候及资源利用等方面发挥着重要的作用[1]。在湿地生态系统中,水文情势和盐分是影响湿地植被分布与演替的关键因素[2]。水位波动和洪水等水文过程是湿地最重要的生态过程之一,对湿地中物种分布格局和植被组成具有决定性作用[3- 4]。盐分是盐碱湿地的主控环境因子,湿地盐渍化直接导致生物多样性的丧失和生态服务功能的下降。目前,全球气候变化和人类活动导致的水文情势改变与盐分聚集已造成大面积的湿地退化和盐渍化,并严重威胁全球湿地生态系统的稳定和健康[2,5- 7]。在此背景下,开展湿地水盐条件变化和植物生理生态相互关系的研究,揭示生境变化对植物种群分布格局的影响,有利于维护湿地生态系统的稳定和健康。
湿地植被作为湿地生态系统的三大重要组成成分之一,能综合反映湿地生态环境的基本特点与功能特性[8]。植物为了抵御环境胁迫,可以通过调整其功能性状和生物量分配格局来增强对逆境的适应,提高获取限制性资源的能力,维持自身的生长发育,该策略是植物对环境异质性胁迫生态适应对策的重要体现[9- 10]。由于特殊的生长环境,湿地植物经常遭受水文条件的大幅度变化,在长期的适应进化过程中形成了调整生活史、形态学特征和生理生化等方面的各种生存策略[11]。因此,研究湿地植被对不同水盐环境下的生态响应特征和规律,对于了解湿地植物在环境胁迫下的生存策略、植被分布格局及演替规律等方面有着重要的理论意义和实践价值。
莫莫格国家级自然保护区位于松嫩平原西部边缘,是典型的盐碱湿地类型保护区,也是濒危物种白鹤(Grusleucogeranus)、丹顶鹤(Grusjaponensis)、东方白鹳(Ciconiaboyciana)等珍禽的重要栖息地、繁殖地和东亚候鸟南北迁徙主干线上的重要停歇中转站。该保护区广泛分布着三江藨草(Scirpusnipponicus)群落。三江藨草的地下球茎中含有丰富的淀粉和蛋白质,是白鹤和其他一些植食性水禽的重要食源,其生长情况直接影响水禽的生境质量。近年来,莫莫格保护区由于降水量减少等气候变化和人为活动干扰,湿地盐碱化加剧,部分区域的三江藨草原生生境淹没,无法正常生长发育[12- 13]。因此,本研究以莫莫格湿地典型湿地植物三江藨草为研究对象,探究水位和盐度变化对三江藨草生长和生物量分配的影响,弥补水、盐相互作用对内陆盐沼植物影响研究方面的不足,有助于深入阐明三江藨草对水盐条件变化的适应性及其响应机制,为保护珍稀物种白鹤栖息环境、指导湿地水盐管理和植被生态恢复提供科学依据。
1 材料与方法
1.1 研究区概况
莫莫格国家级自然保护区位于松嫩平原西部吉林省白城市镇赉县东南端,地理坐标为北纬45°42′25″—46°18′00″,东经123°27′00″—124°4′34″。保护区2013年加入国际重要湿地名录,总面积14.4万 hm2,其中80%以上为湿地,是东亚候鸟南北迁徙的主要途经地。样地海拔约为142 m,区内地势较平坦,相对高差仅2—10 m。气候春秋干旱、夏季多雨、冬季少雪,属温带大陆性季风气候,年均温度4.2℃,多年平均降水量391.8 mm,年平均蒸发量达1553 mm。该地区土壤类型主要为沼泽土,沿江、河地区分布有草甸土、黑钙土和冲积土,有机质含量一般在1.00—9.37%之间。区内植被属大兴安岭植物区系,以低湿草甸和草原草甸为基本类型。
1.2 实验材料
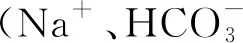
1.3 实验方法
实验共持续60 d,实验结束时,将塑料桶中三江藨草整株取出,在室内用水冲洗干净后测定三江藨草植株的株高、分株数。测量后,将三江藨草分解为根、球茎、地上部分共3个部分,在65℃条件下烘干至恒重,记录其生物量参数(球茎生物量、地下生物量、地上生物量、单株总生物量),并计算生物量分配指标。其中,生物量分配指标包括球茎生物量分配(球茎生物量/总生物量)、地下生物量分配(地下生物量/总生物量)、地上生物量分配(地上生物量/总生物量)。
1.4 数据处理
通过双因素方差分析(Two-way ANOVA)方法分析不同水位、盐度处理及二者交互作用对三江藨草各功能性状指标的影响,采用Tukey多重比较方法进行后续检验,统计显著性水平设置为0.05。数据在分析前对方差的齐次性和正态分布性作检验,不符合要求的进行数据转换使之符合统计要求。数据的处理分析及绘图采用统计软件包IBM SPSS Statistics 22和SigmaPlot 12.5统计软件完成。
2 结果
2.1 水位和盐浓度对三江藨草生长的影响
双因素方差分析(表1)表明,仅水位梯度对三江藨草株高具有显著影响(P<0.05),盐度及两者交互作用则不存在显著影响(P>0.05)。在相同盐浓度梯度下,随着水位的增加,三江藨草成株株高均呈现先增加后下降的趋势(图1),在300、1000 mg/L和2000 mg/L盐度处理中,株高于20 cm水位处取得最大值,分别为1.61、1.45 m和1.6 m;在3000、4000 mg/L和5000 mg/L盐度处理中,于0 cm处取得最大值,分别为1.67、1.36 m和1.5 m。在相同水位梯度下,株高随盐度变化没有规律的趋势,且仅在-10 cm和20 cm水位时,株高在不同盐度间有显著性差异(P<0.05),均表现为低盐度处理高于高盐度处理,其余水位条件下株高没有显著差别。

表1 不同水位和盐度梯度下三江藨草功能性状响应的双因素方差分析
如图1所示,在相同盐度条件下,三江藨草成株株数随水位升高呈波动变化趋势,且水位梯度及其与盐度处理的交互作用对株数的影响不显著(P>0.05,表1)。在相同水位条件下,仅在0和-10 cm处,即三江藨草植株处于水面以上的状态时,株数在不同盐度间有显著性差异(P<0.05),3000 mg/L盐度处理的成株株数取得最大值,盐度处理显著影响三江藨草成株的株数(表1),但随着水位的升高,不同盐度处理间成株株数差异不显著。
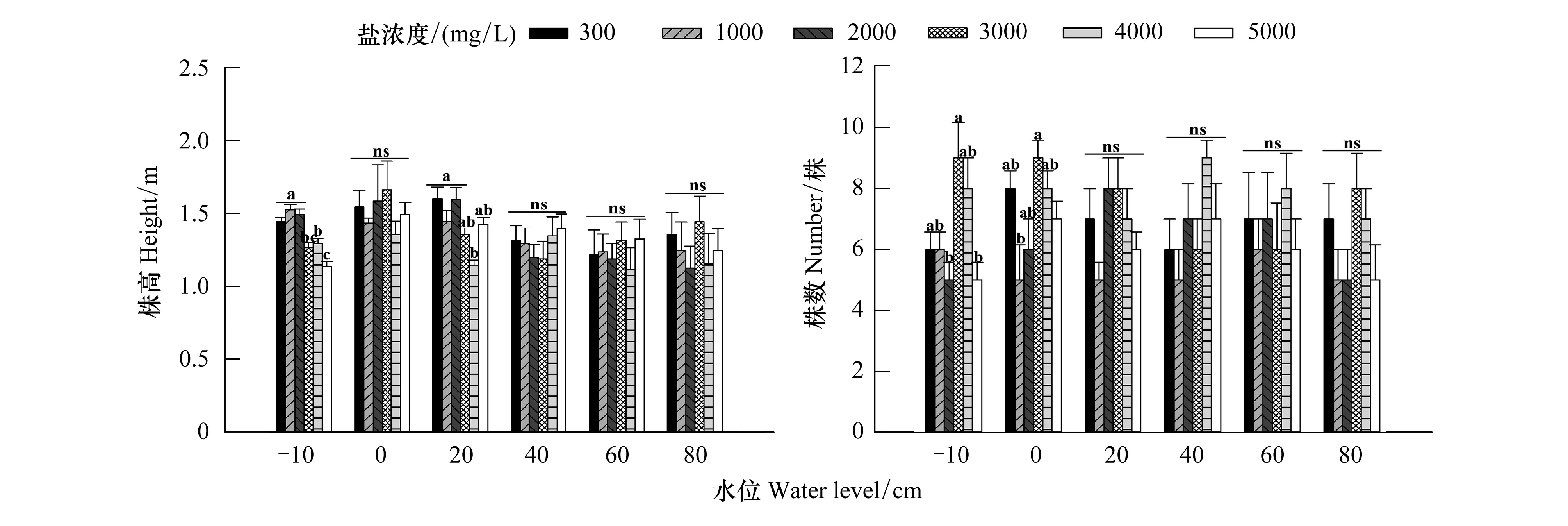
图1 水位和盐浓度对三江藨草生长的影响(平均值±标准误差)Fig.1 The effect of water level and salinity concentration treatments on growth of S. nipponicus (Mean ± SE)不同小写字母代表在相同水位条件下,不同盐分之间差异显著 (P<0.05),ns表示差异不显著
2.2 水位和盐浓度对三江藨草生物量积累的影响
三江藨草的球茎、地下、地上生物量和单株总生物量在不同的水位梯度、盐度及其交互作用下均存在显著差异(P<0.05,表2)。在300、4000、5000 mg/L盐度处理下,三江藨草的球茎、地下、地上生物量和单株总生物量均随水位的升高呈现先上升后下降的趋势,其中球茎生物量于40 cm水位处达到最大值,地下、地上生物量和单株总生物量于60 cm处达到最大值(图2);在1000、2000、3000 mg/L盐度处理下,三江藨草的球茎、地下、地上生物量和单株总生物量变化趋势一致,均随水位的升高而升高,并于80 cm水位处达到最大值(图2)。
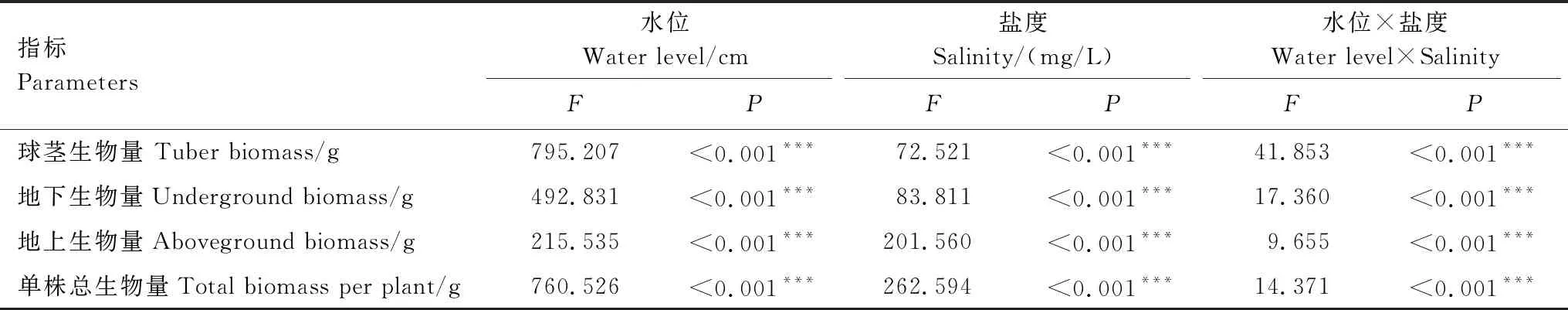
表2 不同水位和盐度梯度下三江藨草生物量响应的双因素方差分析
在-10 cm水位条件下,除300 mg/L盐度处理有球茎产生,其余盐度处理均无球茎的生成,故未在图中显示。总体上,在相同水位条件下,三江藨草的球茎、地下、地上和单株总生物量随盐度的升高呈先上升后下降的趋势(图2),具体表现为:1000、2000、3000 mg/L盐度处理显著高于300、4000、5000 mg/L盐度处理。

图2 水位和盐浓度对三江藨草生物量的影响Fig.2 The effect of water level and salinity concentration treatments on biomass of S. nipponicus
2.3 水位和盐浓度对三江藨草生物量分配的影响
三江藨草的球茎生物量分配、地下生物量分配和地上生物量分配在不同的水位梯度、盐度及其交互作用下均存在显著差异(P<0.05)。如图3所示,在300、3000 mg/L和4000 mg/L盐度处理下,三江藨草球茎生物量分配随水位的升高,表现为先升高后下降的单峰变化趋势,均在40 cm水位处达到最高值,分别占单株总生物的38.5%、36.21%和47.41%,在1000 mg/L和2000 mg/L盐度处理下,球茎生物量分配呈上升趋势,在5000 mg/L盐度处理下则表现为40 cm和80 cm水位处的双峰变化趋势;三江藨草地下生物量分配在300、4000 mg/L和5000 mg/L盐度处理时,随水位的升高呈现先上升后下降的趋势,均于60 cm水位处取得最大值,分别为53.9%、48.36%和48.69%,在其余盐度处理条件下,随水位变化普遍呈现上升趋势;三江藨草地上生物量分配在300、4000 mg/L和5000 mg/L盐度处理时表现为“U型”变化,于60 cm水位处取得最小值,分别为46.1%、51.64%和51.31%,其余则普遍呈下降趋势。
在0—40 cm水位条件下,三江藨草球茎生物量分配随水位的升高表现为先下降后升高的变化趋势,300、4000 mg/L和5000 mg/L盐度处理显著高于其他处理,而在60 cm和80 cm水位处,300 mg/L盐度处理显著低于其他处理(图3);三江藨草地下生物量分配在-10—60 cm水位条件时均表现为300 mg/L盐度处理显著高于其他处理,而在80 cm水位时表现为:300、4000 mg/L和5000 mg/L盐度处理显著低于于其他处理(图3);三江藨草地上生物量分配随水位变化趋势与地下生物量分配相反(图3)。

图3 水位和盐浓度对三江藨草生物量分配的影响Fig.3 The effect of water level and salinity concentration treatments on biomass allocation of S. nipponicus
3 讨论
水位波动是自然环境中不可避免的环境变量,其对湿地植被的影响是不容忽视的,植物的地上部分变化如株高、茎长、叶面积等是对水位变化最直观的响应[14]。本研究发现,水位变化显著影响三江藨草株高,但对其分株数无显著影响,随水位的升高三江藨草株高呈现先增加后下降的趋势,在0—20 cm水位处达到最大值。大量的野外及室内实验表明,在适宜的水位条件下,水位升高能够促进湿地植物株高生长[15- 16],而水位过高则会限制其生长[13]。由于淹水环境中氧气和光照资源的限制,植物需要通过伸长茎部或节间距生长到水面之上,来获取更多的资源[17- 18],促进空气与植物体间的交换,满足植物自身生长的需求。还有部分植物会在体内积累植物激素乙烯,赤霉素等诱导基因表达,使其节间伸长[19]。本研究中,三江藨草是莎草科藨草属(Scirpus)的大型挺水植物[20],在淹水条件下,挺水植物为减轻水淹胁迫作用造成的危害,通过茎长伸长的方式形成发达的通气组织是一种非常重要的生存机制[11,21],在20 cm水位处三江藨草的株高达到最大值。而当水深超过一定深度后,植物体内部O2运输受限,浓度降低,植物的生长速率降低[22- 23]。三江藨草茎秆柔软,呈三棱形,这种生理形态使其在高水位条件下易折断。因此当水位过高时,三江藨草株高降低,植株向上伸长生长逐步转变为横向生长,增强茎部的组织生长以保证在高水位条件下生存。Gibberd M R等[24]研究表明,多数湿地植物组织间孔隙度在水淹后迅速提高,且孔隙度随着直径的增加而增加。
三江藨草各部分生物量及生物量分配均受到水位的显著影响,就生物量而言,当水位超过40 cm时,三江藨草各部分生物量较高,表明高水位更利于三江藨草的生长。生物量的变化是湿地植物应对外界环境改变的综合响应[25],湿生植物和挺水植物对水位变化的响应具有相似的规律[14],荆三棱[26]、菖蒲[27]、梭鱼草[28]等生物量积累随着水位升高有所增加。这可能是受植物固有的生态位影响,在植物自身耐受阈值内水位上升,有利于湿地植物的生长,如芦苇通常出现在湿地中较高区域的上部边缘,这些区域一般淹水较浅或仅有间歇性淹水[29],古勇波等综合三江藨草幼苗的生长特征和生物量累积对不同水位处理的响应,认为其生长的最适宜水位为35 cm[30]。在生物量分配方面,三江藨草的球茎生物量分配和地下生物量分配随水位升高而增加,地上生物量分配则表现为下降。这与以往研究不同,大多数研究表明[26,30- 31],随着水位的增加,植物通常会通过增加地上部分生物量分配比例以增加与空气的接触面积,提高氧气的获取速率。这可能是由于随着水深的增加,三江藨草地下生物量中球茎生物量的比重增加,即高水位环境中,三江藨草以无性繁殖方式达到生存和发展的目的。尽管地下生物量呈上升趋势,但地上生物量总体高于地下,可以满足三江藨草在高水位环境中对氧气和光照的需求。
盐分主要是通过离子毒害、渗透胁迫或二者交互作用对植物产生影响[32]。在本次试验中,盐浓度对三江藨草分株数有显著影响,但盐度对其株高无显著影响。已有实验证明植物对盐胁迫具有一定的耐受性,在植物的耐受阈值内,一定的盐分会促进植物生长[33],如盐地碱蓬在地下水位较浅时,添加1%的NaCl溶液能显著增加其分株数[34],三江藨草幼苗在pH为7.57,Na+浓度为25 mmol/L时,其分株数增加,随着盐碱水平的增加,其株高、分株数和球茎显著降低,甚至在高盐碱水平(pH为10.42,Na+浓度为200 mmol/L)处理下,三江藨草幼苗全部死亡[35]。本研究也得到了相似的结论,这可能是因为具有克隆繁殖特性的植物在受到盐胁迫时,会增强其根状茎或其他克隆组织进行无性繁殖的能力[36]。同时本研究发现盐度对分株数的影响受水位的限制,适宜盐度促进浅水域植株的分株数增加,Salter 等[37]对MelaleucaericifoliaSm.的研究也表明,盐度可能会限制水生植物对水的耐受范围。在盐胁迫条件下,环境中的渗透压发生改变,植物需要消耗大量的ATP[38]或通过其他生理生化方式进行调节,此时水位升高会加剧环境对植物的压迫,致使其生长、繁殖能力下降。
盐分也显著影响三江藨草各部分生物量及生物量分配,且盐分和水位的交互作用对三江藨草生长影响显著,在低浓度和高浓度盐分处理时,三江藨草生物量于40 cm水位达到最大,而中浓度盐分环境中,三江藨草生物量于80 cm水位达到最大,这进一步说明适宜的盐分处理可以增加三江藨草对水深的耐受程度,而高盐分会降低其对水位的耐受阈值。同时,本研究发现高盐分导致三江藨草地下生物量分配减少。这可能是由于湿地植物主要依靠根系吸收环境中盐分等营养物质,通过减少地下部分的比重可以减少对盐分的吸收,从而降低盐分胁迫对植物的危害[39],如盐地碱蓬、芦苇和海滨锦葵等随盐处理浓度的增加,地下部分生物量减少,表明植物在高盐条件下,可以通过减少地下部分如根和根状茎的生物量分配比例来增强自身的适应性[34,40]。
4 结论
三江藨草的生长和生物量分配对环境中水盐条件变化的响应显著,随着水位和盐度的升高,三江藨草球茎、地下、地上和单株总生物量均呈先增加后降低的趋势,株高和株数呈波动变化,适宜的水位能够促进三江藨草的株高增加和生物量积累。高水位环境中,植株对地下部分(包括根和球茎)的投资增加,表明三江藨草侧重无性繁殖方式来适应水位升高的变化。盐分和水位的交互作用对三江藨草生长影响显著,在低浓度和高浓度盐分处理时,三江藨草生物量于40 cm水位达到最大,而中浓度盐分环境中,三江藨草生物量于80 cm水位达到最大,说明适宜的盐分处理可以提高三江藨草对水深的耐受程度,而高盐分会降低其对水位的耐受阈值。本研究结果表明,三江藨草适应生长的水盐环境范围较广,在40—80 cm水位和1000—3000 mg/L盐分处理环境中生长状态较好。研究结果对松嫩平原湿地退化和盐渍化背景下三江藨草种群分布格局和演替规律的预测具有重要的参考价值。
猜你喜欢
新疆农业科学(2022年7期)2022-08-10
中国农学通报(2022年16期)2022-07-08
农业科技与信息(2022年9期)2022-06-18
中国麻业科学(2021年6期)2022-01-21
中国现代中药(2020年4期)2020-06-10
临床检验杂志(电子版)(2020年1期)2020-04-03
西南农业学报(2018年9期)2018-10-19
学苑创造·B版(2015年12期)2016-06-23
中国新技术新产品(2015年3期)2015-07-19
河北渔业(2014年3期)2014-07-08
