
外源胰岛素和能量限饲对鸡PPP1R3C表达的效应
2021-09-27 09:03:26高林歌邵冰豪朱星浩郭钰君黄艳群
畜牧兽医学报 2021年9期
高林歌,邵冰豪,朱星浩,陈 博,郭钰君,黄艳群,陈 文
(河南农业大学 饲料营养河南省工程实验室,郑州 450002)
本试验拟通过研究外源胰岛素和限饲处理对PPP1R3C的表达效应,揭示鸡PPP1R3C的表达调控特性和潜在功能。蛋白磷酸酶1(PP1)全酶是由催化亚基(PP1c)和调节亚基组成,其活性主要靠调节亚基的磷酸化和变构调控来控制[1]。胰岛素的一个基础作用是通过促进葡萄糖进入肌肉和脂肪组织合成糖原,并抑制肝糖原的输出来降低血糖浓度[2]。肌肉和肝中糖原的含量受两种关键的酶调控,即糖原磷酸化酶(GP)和糖原合成酶(GS)[3]。GS和GP的活性受磷酸化和去磷酸化的调控,GS受蛋白激酶A和糖原合成酶激酶3(GSK3)的磷酸化抑制,通过糖原合成酶磷酸酶(GSP)去磷酸化激活,而GP可被磷酸化激酶激活,并被蛋白磷酸酶1的去磷酸化而抑制[4]。根据GenBank数据库,共有7个调节亚基被推断为人类糖原靶向亚基,而这7个 编码调节亚基的基因分别被称为PPP1R3A~PPP1R3G[5]。它们都包含有PP1c调节基序[6]、糖原结合结构域和底物结合结构域[7-8]。PP1对糖原代谢酶具有调节作用,因此在糖代谢过程中起着非常重要的作用。
PPP1R3C是蛋白磷酸酶1调节亚基3C,在人体内又被称为PPP1R5或PTG,主要在骨骼肌和肝中高度表达[9-10]。目前,PPP1R3C的研究主要集中于哺乳动物的糖原代谢酶活性调节以及糖原的合成[11-12]。Zhai等[13]通过敲除小鼠PPP1R3C基因发现,小鼠胰岛素敏感性显著增强且能量消耗增加。另有研究表明,敲除小鼠原代肝细胞中的PPP1R3C降低了肝糖生成,与正常小鼠相比PPP1R3C在肥胖小鼠体内表达量升高[14]。PPP1R3C也是一种新型的低氧诱导因子的靶基因,Mohindra等[15]发现,在短期缺氧条件下PPP1R3C在印度鲶鱼的胰岛素敏感组织(肌肉和肝)中表达量显著增加。另外,PPP1R3C也与一些疾病的发生相关,Chown等[16]敲除PPP1R3C降低了大鼠骨骼肌葡聚糖的积累,减少了成年大鼠葡聚糖体病的发生;Dang等[17]发现,苓桂术甘汤能抑制大鼠PPP1R3C的表达,从而减轻非酒精性脂肪肝病。PPP1R3C在鸡上鲜有研究报道,Ji等[18]利用基因芯片技术发现,PPP1R3C是背部和腿部皮肤的一个差异表达基因,Tian等[19]通过转录组测序发现,PPP1R3C是铬中毒引起鸡肝代谢异常的一个差异表达基因。
目前,关于PPP1R3C在家禽方面尚未有系统的研究。本试验分析了鸡PPP1R3C在不同组织和不同发育时期的表达特性,及外源胰岛素和能量限饲对鸡PPP1R3C的表达效应。掌握鸡PPP1R3C的时空表达特性,揭示外源胰岛素和能量限饲对鸡PPP1R3C表达的影响,相关研究将为深入揭示鸡PPP1R3C基因生物学功能奠定基础。
1 材料与方法
1.1 试验动物及处理
试验一:取相同批次的肉鸡种蛋,于孵化箱37 ℃孵化,在孵化期间于14和19胚龄各挑选10只取胸肌等组织样。待孵化后在相同条件下饲养,自由采食、饮水,饲粮配制参照《鸡饲养标准》(NY/T 33—2004),分别饲喂至7和21日龄,各取10只雌性肉鸡胸肌组织样,于液氮速冻后-80 ℃保存。
试验二:采用相同批次的雄性AA肉鸡于相同条件下饲养,自由采食、饮水,饲粮配制参照《鸡饲养标准》(NY/T 33—2004)。参考本实验室前期的试验分组[20],随机取40只体重相近的24日龄的鸡分为两组(每组20只),试验开始前两组均禁食12 h。试验组腹腔注射0.05 mL·kg-1的胰岛素处理,记为INS组;对照组腹腔注射同剂量的磷酸盐缓冲液(PBS缓冲液)处理,记为PBS组。试验期间所有鸡只均保持自由饮水,试验所用胰岛素用PBS进行稀释(胰岛素∶PBS=1∶9),所用胰岛素为诺和锐短效胰岛素(胰岛素规格3 mL=300 IU)。禁食后在胰岛素和PBS注射之前(0 min)屠宰5只,在胰岛素和PBS注射后15、120和240 min各屠宰5只,采取胸肌、肝和腹脂等组织迅速置于液氮中速冻后-80 ℃ 保存。并以0 min组织样为基础状态,检测PPP1R3C在不同组织中的表达情况。
试验三:相同批次的雌性AA肉鸡于相同条件下饲养、自由采食、饮水,饲粮配制参照《鸡饲养标准》(NY/T 33—2004),饲养至18日龄时随机挑取40只体重相近的鸡分为对照组和能量限饲组。对照组自由采食常规日粮,能量限饲组则实施30%的能量限制(在对照组平均日采食量的80%基础上饲喂低能日粮)的处理,日粮配方参考本实验室前期研究的能量限饲配方[21]。试验进行至48天时,每组各屠宰10只鸡,并采集胸肌和肝等组织置于液氮中速冻后-80 ℃保存。
试验四:取相同批次的雌性AA肉鸡于相同条件下饲养、自由采食、饮水,饲料标准参照《鸡饲养标准》(NY/T 33—2004),饲喂至7日龄时随机挑选30只体重相近的鸡分为3组(对照组、能量限饲组和蛋白限饲组)。对照组自由采食常规日粮;能量限饲组自由采食能量限制饲粮(实行15%的能量限制);蛋白限饲组自由采食蛋白限制饲粮(实行15%的蛋白限制),能量限制饲粮和蛋白限制饲粮的其他营养水平与常规日粮均一致,饲粮配方参照本实验室前期的限制配方[22]。试验进行至21日龄时,每组各屠宰10只鸡,并取胸肌等组织样于液氮中速冻后-80 ℃保存。
1.2 引物设计与合成
根据PPP1R3C基因序列(登录号:XM_423102.5),采用Primer Premier5.0软件设计引物,通过生工生物工程(上海)股份有限公司合成引物。本试验所用的PPP1R3C及β-actin(登录号:NM_205518.1)基因引物序列见表1。

表1 qRT-PCR 引物信息
1.3 RNA提取和cDNA的合成
采用TRIzol法提取肉鸡各组织RNA,利用1.0%琼脂糖凝胶电泳检测总RNA质量,利用分光光度计测定总RNA的浓度以及OD260 nm/OD280 nm。所用反转录试剂盒为HiScript Ⅲ 1 st Strand cDNA Synthesis Kit(+gDNA wiper)(Vazyme,南京)。反转录体系包括两步:1)去除基因组DNA:Total RNA 1 μg、5×gDNA wiper Mix 2 μL,用ddH2O补充至10 μL; 反应程序:42 ℃ 2 min。2)合成cDNA:第一步的混合液10 μL、10×RT Mix 2 μL、HiScript Ⅲ Enzyme Mix 2 μL、Oligo(dT)20VN 1 μL、Random hexamers 1 μL、ddH2O 4 μL;反应程序:37 ℃ 15 min,85 ℃ 5 s,cDNA于-20 ℃保存。
1.4 qRT-PCR检测基因表达量
利用qRT-PCR技术检测PPP1R3C的相对表达量。所用qRT-PCR试剂盒为ChamQ Universal SYBR qPCR Master Mix(Vazyme,南京),qPCR反应体系(20 μL)包括:cDNA模板2 μL,上、下游引物各0.4 μL,2×ChamQ Universal SYBR qPCR Master Mix 10 μL,加ddH2O补充至20 μL;反应程序:95 ℃ 30 s;95 ℃ 10 s,60 ℃ 30 s,共35个循环。以β-actin作为内参,每个反应体系均设置3个生物学重复和技术重复。所有qRT-PCR结果均采用2-ΔΔCt法计算相对表达量[23]。
1.5 数据分析
本试验利用SPSS 25.0进行数据分析。除同一时间点胰岛素和PBS间的显著性采用t检验外,其余数据均采用单因素方差分析,并进行Duncan’s多重比较。以P<0.05为差异显著性判断标准。使用GraphPad Prism 8.0绘制图表。
2 结 果
2.1 鸡PPP1R3C基因组织表达分析
利用采集的试验二肉鸡基础状态(胰岛素和PBS注射之前,0 min)组织样品,首先利用表1中的引物信息,通过RT-PCR扩增检测鸡PPP1R3C在各个组织中的表达量(图1A),RT-PCR结果显示,PPP1R3C在心、胸肌和腿肌中高表达,在脾、肌胃和腺胃等组织表达较低。然后利用qRT-PCR定量检测PPP1R3C在AA肉鸡不同组织中mRNA 的相对表达量(图1B),研究发现,PPP1R3C在胸肌中的表达量最高,而在脾组织中最低。胸肌中PPP1R3C mRNA水平显著高于心肌和腿肌等其余8个组织(P<0.05),PPP1R3C在心肌中的表达量显著高于肝、脾、肺、肌胃、腺胃和腹脂(P<0.05)。而PPP1R3C在其他组织的表达量没有显著差异(P>0.05)。

A.琼脂糖凝胶电泳检测PPP1R3C组织表达谱;B.qRT-PCR检测PPP1R3C 组织表达谱。β-actin为内参基因。不同字母表示P<0.05;相同字母表示P>0.05。Marker.DNA相对分子质量标准
2.2 PPP1R3C在鸡不同发育时期胸肌中的表达
采用试验一中采集的组织样品检测鸡不同发育时期胸肌中PPP1R3C的表达量(图2)。结果表明,PPP1R3C表达量呈现了明显随鸡的发育而升高的趋势。在胚胎发育早期(14胚龄)表达比较低,随后各检测时间点PPP1R3C的表达水平均极显著高于前一个检测时间点(P<0.01)。
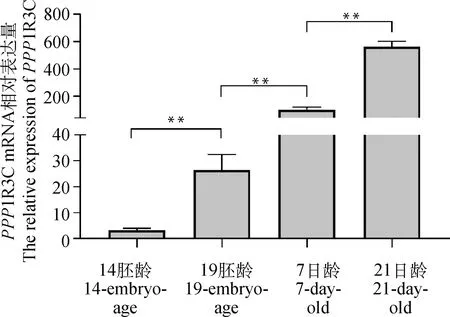
**.P<0.01
2.3 外源胰岛素对鸡PPP1R3C表达的影响
利用试验二中采集到的样品检测了胰岛素注射0(胰岛素和PBS注射之前)、15、120和240 min后胸肌、肝、脂肪组织中的PPP1R3C mRNA表达量(以PBS为对照,图3)。在胸肌组织中(图3A),胰岛素注射显著降低了PPP1R3C在胸肌组织的表达水平,在胰岛素注射后120 min时PPP1R3C mRNA的表达水平显著低于0 min和15 min。PBS注射后PPP1R3C表达量在不同时间点间没有发生显著性变化(P>0.05)。在注射后120 min和240 min时,INS组与PBS组相比PPP1R3C表达量均显著下降(P<0.05),而在注射后15 min时INS组与PBS组的差异没有达到显著水平(P>0.05)。
在肝组织中(图3B),胰岛素注射显著降低了PPP1R3C在肝组织的mRNA表达水平,胰岛素注射后15 min时PPP1R3C表达量显著低于0 min(P<0.05),而15、120、240 min间PPP1R3C mRNA水平没有显著差异(P>0.05)。PBS注射后PPP1R3C的表达呈现波浪式的波动,但总体处于动态平衡。与PBS组相比,在胰岛素注射后120 min时PPP1R3C表达量显著降低(P<0.05),胰岛素注射后15和240 min时与PBS组的差异没有达到显著水平(P>0.05)。
在腹脂组织中(图3C),胰岛素注射后PPP1R3C mRNA水平呈现了完全不同于胸肌和肝脏组织的表达变化趋势。胰岛素注射后PPP1R3C mRNA表达水平迅速上调后又逐渐回落到0 min水平,胰岛素注射后15 min与0 min相比PPP1R3C表达量大约升高了2倍(P<0.05),而120和240 min时与0 min相比PPP1R3C表达量无显著差异(P>0.05)。PBS注射后PPP1R3C表达水平明显降低,15、120和240 min时PPP1R3C表达量均明显低于0 min,而PBS注射后15、120和240 min没有显著差异(P>0.05)。在注射后15 min时,INS组与PBS组相比PPP1R3C的表达水平显著升高(P<0.05),在注射后120和240 min时,INS组与PBS组相比PPP1R3C mRNA表达水平无显著差异(P>0.05)。
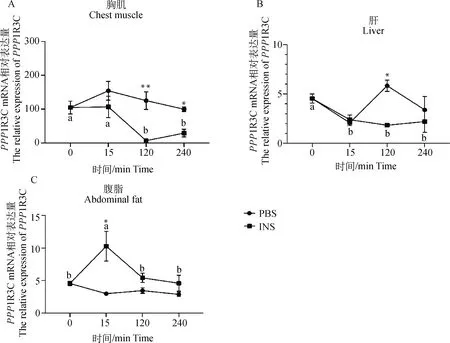
A.胰岛素/PBS注射后胸肌中PPP1R3C的表达;B.胰岛素/PBS注射后肝中PPP1R3C的表达;C.胰岛素/PBS注射后腹脂中PPP1R3C的表达。INS组不同时间点间,不同字母表示P<0.05,相同字母表示P>0.05。同一时间点不同处理间,*表示P<0.05,**表示P<0.01。INS.胰岛素处理组;PBS.对照组
2.4 30%能量限饲对PPP1R3C表达量的影响
由于能量限饲可能导致体内胰岛素分泌水平的改变,本研究进一步开展了能量限饲对PPP1R3C在胸肌和肝表达效应的研究。结果表明,30%的能量限制极显著降低了PPP1R3C在肉鸡胸肌和肝中的表达量(P<0.05)。在胸肌组织中限饲组PPP1R3C的表达量大约降低了73%(图4A),在肝组织中限饲组PPP1R3C表达量大约降低了94%(图4B)。

A.30%能量限饲后胸肌中PPP1R3C的表达;B.30%能量限饲后肝脏中PPP1R3C的表达。**.P<0.01
2.5 15%能量限饲和15%蛋白限饲对胸肌中PPP1R3C表达的影响
为了验证能量限饲对PPP1R3C表达的影响是否具有普遍性,利用实验室构建的群体分析了15%的能量限饲和15%蛋白限饲对PPP1R3C表达的效应(图5)。结果表明,对照组与15%能量限饲组相比,胸肌中PPP1R3C表达没有显著差异(图5A),蛋白限饲与对照组相比,胸肌中PPP1R3C的表达没有显著性变化(图5B)。
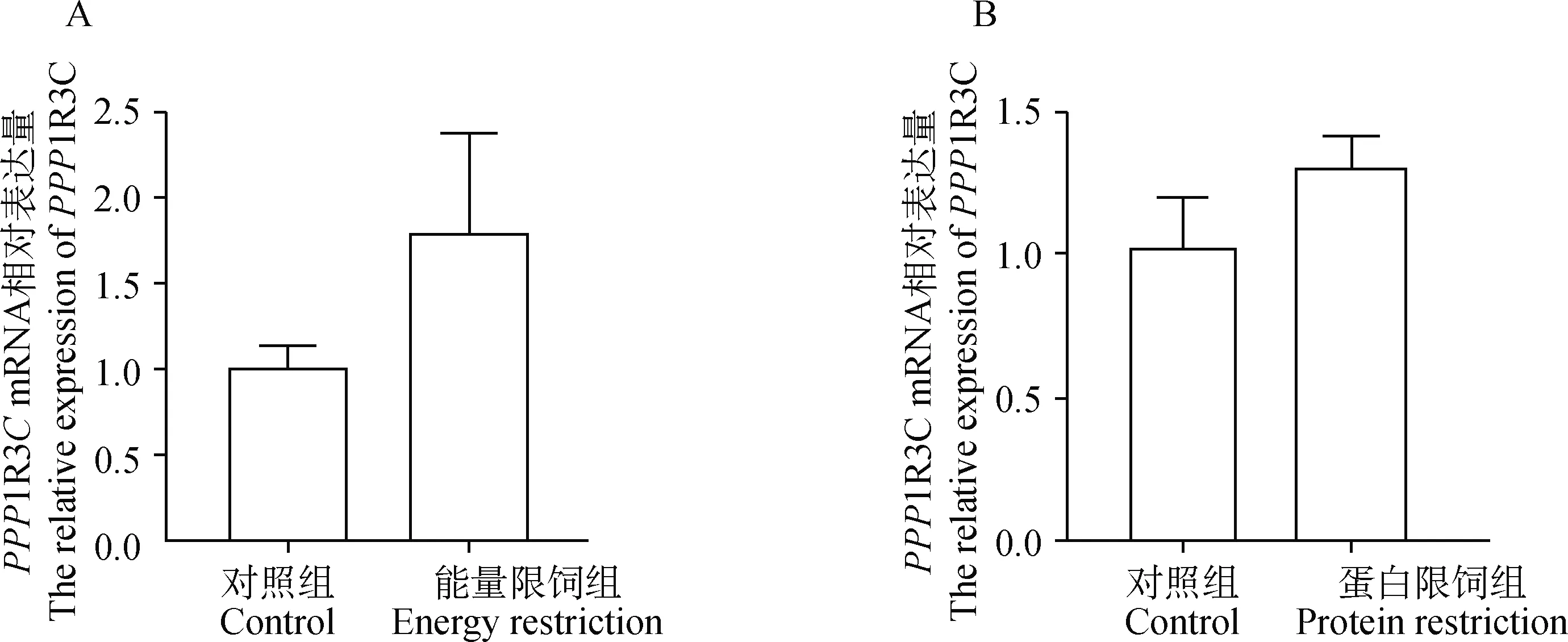
A.15%能量限饲后胸肌中PPP1R3C的表达;B.15%蛋白限饲后胸肌中PPP1R3C的表达
3 讨 论
本研究发现,鸡PPP1R3C具有和哺乳动物[24]相似的组织表达特性,也在骨骼肌中表达较高;时空表达结果显示,随着鸡体的发育胸肌中PPP1R3C的表达呈现升高的趋势,显示了鸡PPP1R3C在肌肉发育中的潜在重要功能。另外,本研究表明,鸡PPP1R3C是胰岛素敏感基因,外源胰岛素显著改变了肉鸡胸肌、肝和腹脂组织中PPP1R3C mRNA水平,且PPP1R3C对外源胰岛素的响应呈现了明显的组织特异性。本课题组前期的研究发现,0.05 mL·kg-1的胰岛素剂量在鸡中是耐受的,胰岛素注射能显著降低肉鸡的血糖水平,在胰岛素注射后120 min内肉鸡血糖浓度几乎呈线性下降,并在120 min时降至最低点[25],直到240 min时仍然保持在低水平[26]。本研究中,胰岛素注射后胸肌和肝组织中PPP1R3C表达呈现了和血糖相似的变化规律,显示了鸡胸肌和肝组织PPP1R3C对禁食血糖浓度的潜在正向调控作用。在哺乳动物中的研究也表明,PPP1R3C是胰岛素敏感基因,在胰岛素调节葡萄糖代谢的过程中起着重要的作用。肝过表达PPP1R3C显著降低了小鼠自由采食状态下血浆胰岛素水平,并提升了禁食状态下的血糖水平[27]。
胰岛素能够刺激肌肉和肝中的糖原合成,在进食的碳水化合物中有20%被贮存在肝,30%贮存在肌肉中[28-29]。有研究表明,人肌肉细胞过表达PPP1R3C时细胞中糖原的积累显著增加[30];Greenberg等[31]发现,在3T3-L1脂肪细胞中过表达PPP1R3C显著提高了脂肪细胞的糖原合成,当添加胰岛素后脂肪细胞中的糖原合成效率进一步增加;Jurczak等[32]发现,过表达PPP1R3C显著增加了小鼠脂肪细胞的糖原积累;当全身性敲减PPP1R3C时小鼠胸肌、肝和脂肪中糖原水平均下降,且小鼠胸肌、肝和脂肪中糖原合成和葡萄糖摄取减少,而附睾脂肪中葡萄糖转运补偿性增加[23-24,33]。上述的研究显示,组织PPP1R3C表达水平与糖原水平呈正相关,高水平的PPP1R3C能够促进细胞中糖原的合成,低表达的PPP1R3C降低了细胞中糖原的积累。本研究中,外源胰岛素降低了肉鸡胸肌和肝中PPP1R3C的表达,上调了脂肪组织(15 min)的mRNA水平,提示鸡体在对外源胰岛素刺激下,一定程度上可能通过降低胸肌和肝脏PPP1R3C水平而减少胸肌和肝组织中糖原的合成和葡萄糖的摄取,而不完全补偿性地增加腹脂中葡萄糖的转运。
肥胖会导致多种并发症,包括2型糖尿病、脂肪肝等[34-35]。长期高能量的摄入会引起胰岛素抵抗,而低能量饮食能够改善机体胰岛素敏感性[36-38]。实验室前期研究发现,30%能量限饲显著降低了AA肉鸡的体重、腹脂重以及甘油三酯和葡萄糖水平[39],而其血清胰岛素水平没有明显的改变。本研究发现,30%能量限制产生了类似外源胰岛素的效应,显著降低了胸肌和肝中PPP1R3C的表达。然而15%能量限饲和15%蛋白限饲均未能显著改变鸡PPP1R3C的mRNA水平,显示了能量限饲对鸡PPP1R3C的效应具有剂量依赖性。另有研究表明,在高脂喂食的小鼠体内PPP1R3C的表达升高了两倍,禁食后再喂食小鼠体内PPP1R3C的表达也显著增加,另外高脂饲喂显著增加了小鼠mTORC1的活性,而敲除PPP1R3C后小鼠的胰岛素敏感性增加[40]。
4 结 论
本试验发现,鸡PPP1R3C呈现了明显的时空动态表达特性,PPP1R3C在所检测组织中广泛表达,在胸肌组织中表达量最高,其在胸肌组织中的表达量随个体发育逐渐上升。外源胰岛素显著下调了PPP1R3C在胸肌和肝的表达,而上调了PPP1R3C在脂肪组织中的早期表达。15%能量限饲和15%蛋白限饲均未能显著影响胸肌中PPP1R3C的表达,而30%能量限饲显著下调了胸肌和肝脏中PPP1R3C的表达,显示了能量限饲对PPP1R3C基因表达影响的剂量依赖性。
猜你喜欢
水产科学(2023年1期)2023-02-02 13:14:02
广西糖业(2022年5期)2022-11-24 11:14:10
肝博士(2021年1期)2021-03-29 02:32:08
今日农业(2020年18期)2020-12-14 19:08:44
中国生殖健康(2019年2期)2019-08-23 08:12:10
兽医导刊(2016年12期)2016-05-17 03:51:44
中国卫生标准管理(2015年1期)2016-01-14 03:41:27
少儿科学周刊·少年版(2015年11期)2015-12-17 23:47:17
少儿科学周刊·儿童版(2015年11期)2015-12-17 03:53:46
药学与临床研究(2015年4期)2015-06-05 11:35:51
