
冷应激对放牧肉牛免疫功能和抗氧化功能的影响
2021-09-26 16:18格日乐其木格
当代畜禽养殖业 2021年4期
格日乐其木格
(内蒙古自治区赤峰市阿鲁科尔沁旗农牧局,内蒙古 赤峰 025500)
牛肉因富含微量元素且具有高蛋白、低胆固醇、低脂肪等优点而受到人们的喜爱,其在肉类市场所占份额也越来越大[1]。为了满足人们对牛肉的需求,就要求畜产品生产向集约化方向转变。但由于我国北方冬季寒冷,会使反刍动物产生冷应激,从而抑制动物的生长速度和增加幼畜的死亡率,故导致了畜牧业经济效益的下降[2]。冷应激在通常情况下可以分成两种,即急性冷应激和慢性冷应激,此可通过动物暴露在寒冷环境中的时间的长短来进行区分。若动物从急性冷应激中进入慢性冷应激,则在急性冷应激中所产生的反应会消失,动物机体代谢活动也随之发生相应的变化,但自身恒温性能不会改变[3]。
之前对奶牛冷应激的研究表明,北方寒冷的环境会降低奶牛的免疫功能、乳品质、采食量以及产奶量。有报道称,当温度为-6.8℃且无风时,欧洲牛也会产生冷应激,这表明动物产生冷应激不仅是因为环境温度低,还会受到风速等的影响。所以对于冷应激的评价方法会因为所采用的环境指标的不同而不同。在相同寒冷的环境条件下,冷应激会严重破坏西门塔尔母牛的免疫功能和抗氧化功能,而蒙古母牛则具有更强的耐寒冷性能[4]。研究还表明,当环境温度在5~25℃时,奶牛不会产生冷应激或热应激。用温湿度指数来判断是否产生冷应激或热应激的范围是:当THI小于38时奶牛产生冷应激,当THI大于72时奶牛产生热应激。而且一旦动物产生冷应激或者热应激,其机体会通过下丘脑-垂体-甲状腺轴(HPT)、交感-肾上腺髓质轴(SAM)以及下丘脑-垂体-肾上腺轴(HPA)来进行调节[5]。但这种调节不是没有极限的,当环境温度和湿度的变化超过动物自身调节的极限时,动物会无法适应环境的变化从而产生冷应激或热应激。之前对于冷应激的研究主要集中于奶牛及单胃动物上,而对肉牛冷应激研究较少。本研究通过探讨冷应激对放牧西门塔尔母牛免疫功能、抗氧化功能的影响,试图进一步了解冷应激对肉牛机体影响的生理机制,以便为我国北方地区在冬季应对放牧肉牛冷应激提供理论基础。
1 材料与方法
1.1 试验动物及饲养管理
将内蒙古赤峰市阿鲁科尔沁旗某天然草场提供的16头4岁,体重为500±20 kg且健康状况良好的放牧西门塔尔母牛作为试验动物。试验用西门塔尔母牛的饲养管理方式为全年放牧,自由采食牧草和饮水。
1.2 试验时间及地点
本试验在内蒙古赤峰市阿鲁科尔沁旗某天然草场进行。试验分为2个阶段:秋季试验(2019年9月15~18日),冬季试验(2020年 1月 10~13日)。
1.3 环境温湿度测定
由于试验是以在天然草场进行全年放牧的西门塔尔母牛为研究对象,所以所测的环境指标均为自然环境下的温度和相对湿度。试验期间,测定每天8∶00、14∶00以及20∶00的天然放牧草场的温度和相对湿度,并监测记录试验当月的平均气温和相对湿度。
1.4 血样采集
在试验期最后一天清晨对每头试验牛进行颈静脉采血。用非抗凝采血管采集10 mL血液,静置半小时后3000 r/min离心10 min,分离制备血清,置于-20℃冰箱中保存,用于血液生化指标的测定
1.5 血液指标测定
血清中 IgA、IgG、IgM、IL-2、IL-4、SOD、MDA、T-AOC含量测定采用酶联免疫分析法,试剂盒均购自南京建成生物工程研究所,测定方法遵照试剂盒说明书之内容。
1.6 数据分析
采用Excel 2003对所有试验数据进行初步整理后,使用SPSS17.0中的单因素方差分析 (one-way ANOVA)进行差异显著性分析,结果用平均值±标准差方式表示,以P<0.05作为差异显著性判断标准。
2 结果与分析
2.1 肉牛放牧草场的温湿度
由表1可知,在不同的试验阶段,环境温湿度是不同的,所以对试验牛所造成的影响也是不同的。秋季试验期放牧草场的最低温度为5℃,最低湿度为30%。冬季试验期放牧草场的最低温度为-28℃,最低湿度为21%。根据试验当月温度和湿度的监测结果发现,秋季放牧草场的月平均气温为16℃,月平均湿度为45%;冬季放牧草场的月平均气温为-17.5℃,月平均相对湿度为30%。
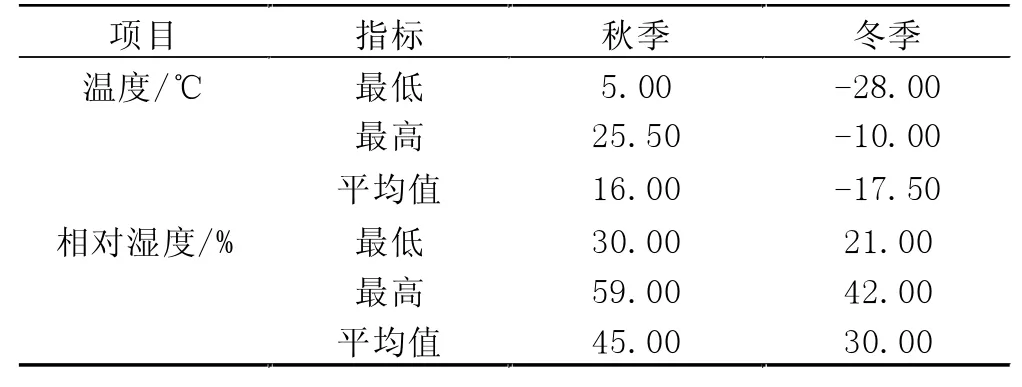
表1 不同季节平均气温和湿度
2.2 不同季节西门塔尔母牛血清免疫指标变化情况
通过比较秋季和冬季放牧西门塔尔母牛的血清免疫指标结果可知,冷应激使放牧西门塔尔母牛血清中 IgG、IL-2 的含量显著下降(P<0.05),与秋季血清免疫指标相比,IgA、IgM、IL-4含量有下降的趋势,但差异不显著(P>0.05)。

表2 不同季节西门塔尔母牛血清免疫指标测定结果
2.3 不同季节西门塔尔母牛抗氧化指标变化情况
通过比较秋季和冬季放牧西门塔尔母牛的血清抗氧化指标结果可知,天然草场的冷应激使放牧西门塔尔母牛SOD含量出现增加的趋势,但差异不显著(P>0.05)。天然草场的冷应激使放牧西门塔尔母牛血清中MDA的含量显著上升(P<0.05)。天然草场的冷应激使放牧西门塔尔母牛血清中的T-AOC显著下降(P<0.05)。

表3 不同季节西门塔尔母牛血清抗氧化指标测定结果
3 讨论
3.1 不同季节的温度和湿度指数对放牧西门塔尔母牛的影响
在本试验中,冬季平均温度最低可达-17.5℃,平均相对湿度最低为30%,因此冬季的平均温湿度指数也最低。冬季的平均温湿度指数为22.64,说明本试验中的放牧西门塔尔母牛在冬季处于冷应激状态中。通过研究发现,温湿度指数THI>38为无冷应激,25<THI≤38为轻度冷应激,8<THI≤25为中度冷应激,-12<THI≤8 为重度冷应激,-25<THI≤-12 为极端冷应激[6],因此天然草场的冬季使放牧西门塔尔母牛产生了轻度的冷应激。而在秋季,天然草场的平均温度为16℃,平均相对湿度为45%,平均温湿度指数为59.95,因此通过温湿度指数可知,放牧西门塔尔母牛在秋季处于无冷应激状态。又因为THI≤72时,牛无热应激反应;72<THI≤79 时,牛处于轻度热应激;79<THI≤88时,牛处于中度热应激;THI>88时,牛处于重度热应激[7],所以在秋季放牧西门塔尔母牛也不产生热应激。
3.2 冷应激对放牧西门塔尔母牛免疫指标的影响
研究发现,无论高温还是低温都会增加牛的死亡率,而死亡率最低时的温度条件为15~20℃。通过对极端高温进行相对风险估计了解到,其风险范围为1.3~2.5,不仅温度强度会影响牛的死亡率,暴露在高温或者低温中的时间长短也会对牛的死亡率产生一定的影响,而且它们与牛的死亡率成正比[8]。在环境温度为-30℃时,绵羊的免疫球蛋白含量会极显著地下降,这说明重度冷应激会使绵羊的免疫功能受到抑制[9]。动物淋巴结中的浆细胞和脾脏能合成和分泌免疫球蛋白IgM、IgG以及IgA,这些免疫球蛋白会在动物的特异性免疫中发挥重要作用[10]。其中IgG占血清总免疫球蛋白的比例约为75%。IgG在动物体液免疫中发挥着重要作用,IgG还可用作为血清学诊断的抗体[11]。在有抗原刺激的体液免疫中最先产生免疫应答的免疫球蛋白是IgM,其在动物的早期免疫防御中发挥着重要作用。动物血液中免疫细胞因子的含量能够反映动物免疫功能的高低[12]。细胞的生长因子包括IL-2,免疫系统中白血球的细胞活性能够通过IL-2调控,IL-2同时还可以参与抗体反应,在这个过程中IL-2的主要作用是增强巨噬细胞以及T淋巴细胞的活性[13]。对动物体液免疫起到调节作用的细胞生长因子还有IL-4,IL-4的作用是诱导B细胞的成熟和刺激B细胞成熟免疫球蛋白,从而参与动物的免疫反应[14]。通过在寒冷的冬季对三河牛的研究发现,血清中的IL-2、IL-4以及IgG的含量均会有所下降,但与春季非应激期相比下降不显著[15]。有研究发现,肉牛在慢性冷应激中会使血清中的免疫球蛋白的主要成分IgG、细胞生长因子中的IL-4、IL-2的含量有所降低,这说明慢性冷应激不仅会使肉牛的免疫功能受到抑制,还会使肉牛产生炎症反应[16]。而且随着冷应激时间的延长,会使动物机体内血清免疫抑制因子的含量升高从而抑制IL-2的产生,进而造成动物机体免疫功能的下降[17]。对奶牛的研究发现,冷应激会降低其免疫功能[18]。在本试验中,IgG、IL-2显著下降,这说明冷应激可抑制免疫球蛋白的合成,使放牧西门塔尔母牛的免疫功能受损。
3.3 冷应激对放牧西门塔尔母牛抗氧化指标的影响
动物在冷应激的状态下,其氧化平衡会被破坏从而导致自由基的大量产生,而过量的自由基会导致脂质过氧化物大量产生,增强脂质过氧化的作用,进而导致动物产生氧化应激损伤[19]。动物机体内的抗氧化酶可以清除自由基,保护机体免受氧化应激的威胁。冬季寒冷的天气可能会造成动物产生氧化应激,使SOD的含量增加[20]。脂质过氧化的最终产物是MDA,MDA含量的高低反映氧化应激的剧烈程度,是细胞氧化损伤程度的指标之一[21]。SOD在动物机体内的主要作用是清除超氧阴离子自由基,防止细胞损伤,而且SOD活性的高低能够间接反映机体清除氧自由基的能力。机体的总抗氧化能力通过T-AOC来衡量[22]。冷应激会使肉牛血清中的T-AOC和MDA含量显著升高,而后者升高的原因有可能是因为肉牛体内积累了大量的自由基。通过对西门塔尔杂交犊牛的研究发现,犊牛血清中的MDA和T-AOC的含量与环境温度的高低成反比。不仅如此,在对奶牛的研究中也发现,SOD的含量与非应激期相比会显著降低,而MDA的含量则会显著升高。通过对西门塔尔犊牛在冷应激中的抗氧化指标的研究发现,在环境温度为-40℃时,MDA含量显著升高,而SOD 的活性显著降低[23]。造成这种现象的原因可能是因为动物在初冬时遭受了冷应激,所以产生了代偿效应,造成SOD的活性上升。但随着冷应激时间的延长,SOD的活性降低。本试验的研究结果为,T-AOC的含量显著下降,MDA的含量显著上升,而SOD的含量虽然上升但不显著,这表明冷应激使放牧西门塔尔母牛的抗氧化功能降低。
猜你喜欢
海峡姐妹(2021年7期)2021-11-15
辽河(2021年8期)2021-08-30
乡村地理(2021年4期)2021-03-28
长江文艺(2019年5期)2019-08-07
文苑·经典美文(2018年8期)2018-08-16
润·文摘(2018年4期)2018-05-14
儿童故事画报·智力大王(2016年5期)2016-07-04
少儿科学周刊·儿童版(2015年12期)2016-05-16
小哥白尼·野生动物画报(2009年5期)2009-06-18
长城(2009年2期)2009-04-09
