
蓝萼甲素通过cyclinB1/CDK1途径诱导DU145细胞G2/M期阻滞①
2021-09-25 10:07朱琳琳徐祉轩张明明新乡医学院医学检验学院河南省分子检验与医学检验技术协同创新中心河南省免疫与靶向药物重点实验室新乡453003
中国免疫学杂志 2021年18期
朱琳琳 徐祉轩 张明明(新乡医学院医学检验学院,河南省分子检验与医学检验技术协同创新中心,河南省免疫与靶向药物重点实验室,新乡453003)
前列腺癌(prostate cancer,PCa)发病率逐年升高,现已成为严重威胁亚洲男性健康的第二大恶性肿瘤,但相当一部分患者激素治疗不敏感或耐受,迫切需要寻找抗肿瘤效果明确但不良反应较小的药物。本研究选取的蓝萼甲素(glaucocalyxin A,GLA)是从传统中草药香茶菜中分离提取的活性成分,香茶菜作为中草药,用于清热解毒、抗菌消炎、抗肿瘤历史悠久。研究表明GLA具有良好的抗肿瘤作用,但机制尚不明确[1-4]。本研究检测GLA对人前列腺癌细胞系DU145细胞中周期蛋白B1(cy‐clinB1)、周期蛋白依赖性激酶1(cyclin-dependent ki‐nase1,CDK1)等细胞周期相关因子表达的影响,探讨GLA诱导细胞周期阻滞的作用机制,为GLA的临床应用提供实验依据。
1 材料与方法
1.1 材料人前列腺癌细胞系DU145细胞购自中科院上海细胞库。胎牛血清、RPMI1640培养液购自Gibco公司;GLA由新乡医学院药学院提供;Trizol试剂、逆转录酶(M-MLV)、实时荧光定量PCR试剂盒购自天根生化科技(北京)有限公司;所用引物均由上海生工生物工程(上海)股份有限公司合成;所用抗体均购自美国CST公司;BCA蛋白浓度检测试剂盒、流式细胞周期检测试剂盒购自江苏碧云天生物技术公司;流式凋亡检测试剂盒购自美国BD公司。
1.2方法
1.2.1 实验分组实验设空白对照组、1 μg/ml、2.5 μg/ml、5 μg/ml GLA组,4组细胞培养基均含10%胎牛血清、100 U/ml双抗(青链霉素),培养环境为37℃、5%CO2。空白对照组不加GLA,其他各组细胞贴壁后分别加入相应浓度GLA刺激培养。
1.2.2 细胞增殖4组细胞接种于96孔板中,每孔体积100 μl,约2 000个细胞,细胞贴壁后加药。各孔分别加入10 μl CCK-8溶液,96孔板置于37℃、5%CO2培养箱中孵育1 h,取出96孔板,置于酶标仪检测位,振荡混匀后,酶标仪450 nm测定各孔标本吸光度。
1.2.3 细胞周期细胞接种于6孔板中,约1×106个/孔,各孔细胞数量均匀。细胞贴壁后加药,加药后培养24 h,按试剂盒操作说明收集固定细胞,染色,上机检测分析。
1.2.4 细胞凋亡细胞接种于6孔板中,约1×106个/孔。细胞贴壁后加药培养24 h,收集细胞。预冷PBS洗涤细胞,Binding Buffer重悬细胞,Annex‐in V-FITC标记后避光、室温孵育15 min,PI染色后上机检测分析。
1.2.5 Real-time PCR Trizol一步法提取细胞总RNA,异丙醇法浓缩RNA,对RNA浓度和质量检测后进行逆转录,并按试剂盒操作说明完成qRTPCR。qRT-PCR反应条件为95℃预变性10 min,95℃变性15 s与60℃退火/延伸60 s进行40个循环,最后分析熔解曲线,采用2-ΔΔCt法(其中Ct值为循环阈值)分析mRNA的相对表达水平。所用引物序列见表1。
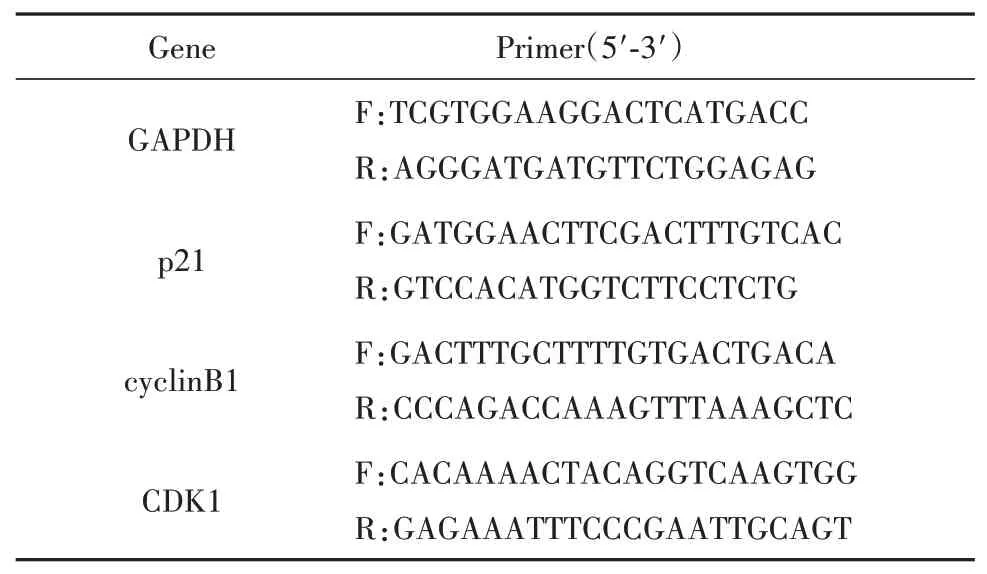
表1 Real-time PCR引物序列Tab.1 Primers used in real-time PCR
1.2.6 Western blot提取细胞总蛋白,BCA法测定蛋白浓度后进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳,电泳结束后依次转膜、室温封闭、孵育一抗(p21、cyclinB1、CDK1为1∶1 000稀释)4℃过夜、孵育HRP-二抗(1∶5 000稀释)室温摇床1 h,洗膜后曝光扫描,以目的蛋白与β-actin蛋白条带灰度值的比值,作为其相对表达量。
1.2.7 免疫荧光采用爬片:在24孔板中培养DU145细胞,细胞贴壁后加药刺激24 h,PBS洗涤细胞后4%多聚甲醛室温固定30 min,0.5%TritonX-100室温通透15 min,室温封闭90 min,一抗孵育(cyclinB1、CDK1分别为1∶400、1∶100稀释),4℃过夜,荧光二抗(1∶400稀释)室温避光孵育1 h,DAPI染色,应用荧光显微镜检测蛋白在细胞中的定位分布。
2 结果
2.1 GLA抑制DU145细胞增殖各组细胞按相应药物浓度分别刺激培养1 h、6 h、12 h、24 h、48 h,检测细胞增殖情况。结果显示,与对照组相比,1 μg/ml GLA对细胞增殖的影响无统计学差异(P>0.05);2.5 μg/ml与5 μg/ml GLA能够抑制细胞增殖,5 μg/ml GLA的抑制效果更为显著(P<0.01),见图1。可见GLA抑制DU145细胞增殖的作用呈现剂量依赖效应。
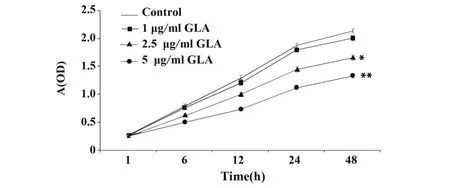
图1 DU145细胞增殖曲线Fig.1 Proliferation curve of DU145 cells
2.2 GLA阻滞DU145细胞周期与对照组相比,1 μg/ml GLA对细胞周期的影响并无统计学差异。随着浓度增加,GLA显现出阻滞细胞周期的作用,当5 μg/ml GLA刺激培养DU145细胞24 h后,细胞周期明显停滞于G2/M期(P<0.01),见图2。GLA阻滞DU145细胞周期的作用呈现出明显剂量依赖效应。
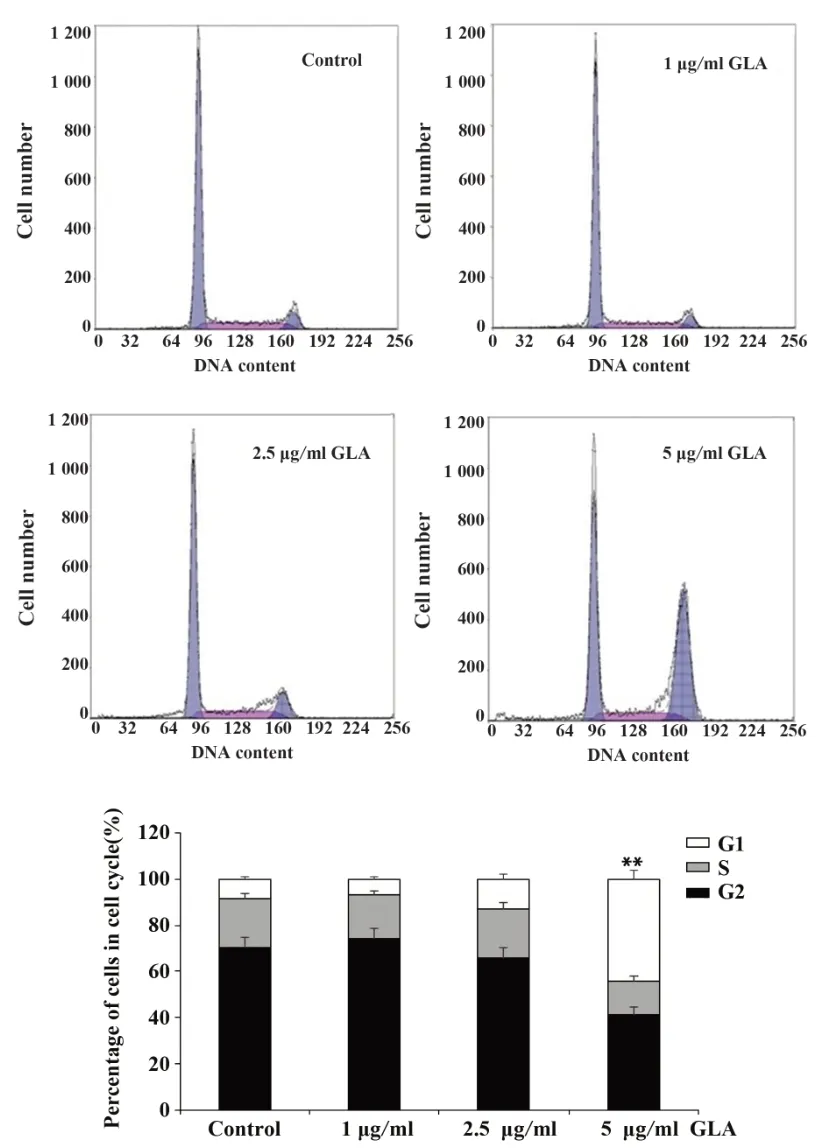
图2 GLA引发DU145细胞G2/M期阻滞Fig.2 GLA induces G2/M cell cycle arrest in DU145 cells
2.3 GLA调节DU145细胞周期蛋白的表达1 μg/ml GLA刺激培养细胞24 h,与对照组相比,p21、cy‐clinB1、CDK1 mRNA与蛋白的表达均无统计学差异。但随着浓度的增加,2.5 μg/ml GLA能够促进p21的表达,并抑制cyclinB1与CDK1的表达;5 μg/ml GLA的作用效果更为明显,在显著促进p21表达的同时,更加抑制了cyclinB1与CDK1的表达,且qRTPCR与Western blot的结果基本一致,见图3。可见GLA调节DU145细胞周期相关因子的表达,在mRNA与蛋白水平上均表现出剂量效应。
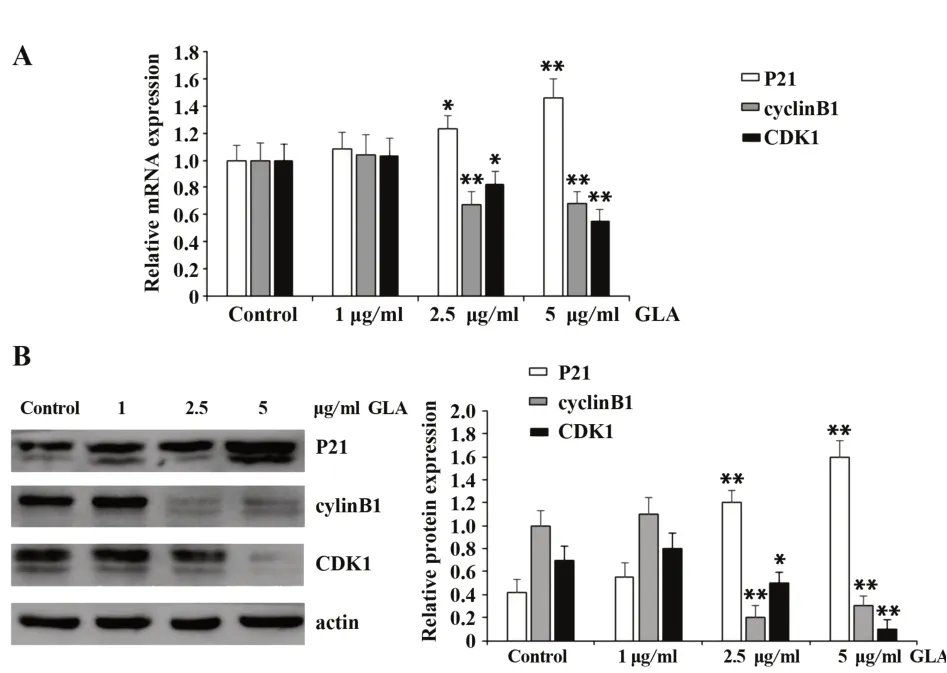
图3 GLA调节DU145细胞周期相关蛋白的表达Fig.3 GLA regulate cell cycle relative protein expression in DU145 cells
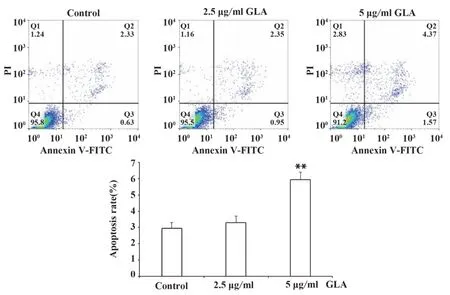
图5 GLA促进DU145细胞凋亡Fig.5 GLA promote DU145 cells apoptosis
2.4 免疫荧光验证DU145细胞周期蛋白的定位表达免疫荧光结果进一步验证了Western blot的结果,细胞周期蛋白cyclinB1、CDK1主要表达于细胞质,5 μg/ml GLA能够显著抑制cyclinB1、CDK1的表达(见图4)。GLA引起DU145细胞周期阻滞与其抑制cyclinB1、CDK1表达的作用效果及剂量效应完全一致,因此推测GLA极有可能通过抑制cyclinB1、CDK1的表达而导致细胞周期阻滞于G2/M期。

图4 DU145细胞周期蛋白细胞内表达(×400)Fig.4 Localization of cell cycle relative protein in DU145 cells(×400)
2.5 GLA诱导DU145细胞凋亡GLA阻滞细胞周期后极有可能引发细胞生物学行为发生变化。流式检测结果显示:2.5 μg/ml GLA作用于细胞,虽然能引起凋亡增加,但与对照组相比并无统计学差异(P>0.05),5 μg/ml GLA才显 示 出促 凋 亡 效 果(P<0.01)。
3 讨论
前列腺癌为常见恶性肿瘤,近年来随着经济水平的不断提高,人口老龄化进一步明显,我国前列腺癌患者呈逐年上升趋势[5-6]。大多数前列腺癌发病隐匿、进程缓慢,就诊时往往已是中晚期,临床多用雄激素治疗,但很多患者治疗后期出现激素不敏感或耐受,最终肿瘤转移扩散[7]。因此迫切需要寻找抗肿瘤效果明确但不良反应少的药物。
传统中草药一直以其活性强、毒性小备受关注,其中香茶菜在民间应用历史悠久[1-4],已用于临床的冬凌草就是从香茶菜中提取的活性成分,本研究选取的GLA则是香茶菜的另一种活性成分。前期研究发现,小剂量GLA能够通过IL-6激活Stat3信号通路,调节细胞炎症反应,促进肿瘤细胞增殖与侵袭迁移[8-9]。中草药往往对细胞功能具有双向调节作用,研究也表明GLA具有抗炎、抗肿瘤、免疫调节等作用[3-4]。因此,本研究采用较高浓度GLA作用于前列腺癌DU145细胞,探讨GLA抗肿瘤的作用机制。
增殖实验结果显示,2.5、5 μg/ml GLA作用于DU145细胞24 h后,可以抑制DU145细胞的增殖,尤以5 μg/ml GLA的作用效果更为显著。与前期实验结果一致的是,与对照组相比,1 μg/ml GLA对细胞增殖的影响作用并无统计学意义。这些结果均提示GLA抑制细胞增殖的作用呈剂量效应及时间依赖性。增殖与细胞周期关系密切,周期紊乱可以引起增殖异常。
细胞周期结果显示,与对照组相比,1 μg/ml GLA对细胞周期基本无影响。随着浓度增加,2.5 μg/ml GLA表现出阻滞作用,5 μg/ml GLA的阻滞作用尤为显著,G2期细胞达44.3%,整体上凸显G2/M期阻滞。G1期到S期、G2期到M期是复杂活跃的分子水平变化时期,容易受环境条件影响。已有研究发现,白藜芦醇能够抑制DU145细胞从G0/G1期向S期转变,从而抑制其增殖[10]。本研究结果提示GLA能够显著影响DU145细胞从G2期向M期的转变,进而阻滞细胞周期,抑制其增殖。
参与细胞周期调控的蛋白主要有S、M、G1期蛋白与CDK激酶。cyclin B为M期周期蛋白,从S期开始表达,在G2/M期到达峰值,后期消失。CDK1的活化依赖于cyclinB1,cyclinB1与CDK1结合并激活CDK1可以促进细胞由G2期进入M期[11]。p21则是CDK激酶的抑制剂,能够抑制cyclinB1/CDK的激活,阻滞细胞G2/M期转变[12]。鉴于GLA引发G2/M期阻滞,因此本研究进一步对p21、cyclinB1、CDK1进行检测。结果显示,5 μg/ml GLA作用DU145细胞24 h,促进p21表达的同时,显著抑制cyclinB1、CDK1的 表 达。GLA对p21与cyclinB1/CDK1的 作用效果正好相反。与此一致的是,LIN等[13]研究发现,GLA能够通过促进/抑制p21、Cdc25C等周期蛋白的表达,阻滞膀胱癌UMUC3的细胞周期。免疫荧光进一步印证了Western blot结果。与对照组相比,5 μg/ml GLA刺激培养24 h,DU145细胞质中cyclinB1与CDK1的表达明显增多。因此GLA极有可能通过抑制周期蛋白cyclinB1的表达,影响CDK1激酶的活化,进而导致G2/M期阻滞。
细胞周期紊乱会诱发凋亡,细胞周期长时间停滞也会诱发凋亡。流式结果就显示出5 μg/ml GLA能够引起DU145细胞凋亡,提示GLA引起周期阻滞的同时,也诱发了凋亡。已有研究发现,GLA能够抑制PI3K/Akt信号通路,导致p21、Bax蛋白累积,继而激活下游信号通路,引起细胞周期阻滞和凋亡[13-15]。本研究结果也显示GLA诱发G2/M期阻滞与p21蛋白增多相关,提示GLA极有可能通过p21蛋白信号通路诱导DU145细胞凋亡,具体作用机制在后续实验中进一步探索。
综上所述,本研究发现GLA能够诱导DU145细胞周期阻滞于G2/M期,这一作用极有可能是通过抑制细胞周期蛋白cyclinB1、CDK1的表达而诱发,cyclinB1、CDK1表达升高进一步激活凋亡信号通路,促进细胞凋亡。因而,控制GLA的作用浓度及作用时间,有效发挥其阻滞细胞周期的作用,对前列腺癌防治有新的意义和价值。
猜你喜欢
核科学与工程(2022年3期)2022-10-18
广州化工(2020年6期)2020-04-18
国际呼吸杂志(2019年4期)2019-03-12
现代矿业(2018年9期)2018-10-16
中成药(2017年9期)2017-12-19
中成药(2017年5期)2017-06-13
中华老年多器官疾病杂志(2016年7期)2016-04-28
癌症进展(2016年10期)2016-03-20
华南农业大学学报(2015年5期)2015-12-04
医学研究杂志(2015年5期)2015-06-10
