
外源Mg2+对不同水稻品种苗期Cd2+吸收转运特性的影响
2021-09-25 05:01普天贇何腾兵付天岭高珍冉罗德强
耕作与栽培 2021年4期
普天贇, 何腾兵,, 滕 浪, 付天岭, 高珍冉, 秦 松, 罗德强
(1.贵州大学农学院, 贵阳 550025; 2.贵州大学新农村发展研究院, 贵阳 550025;3.贵州省农业科学院土壤肥料研究所, 贵阳 550025;4.贵州省农业科学院水稻研究所, 贵阳 550025)
水稻(OryzasativaL.)、小麦(TriticumaestivumL.)等禾谷类作物籽粒中易积累Cd,水稻在低Cd2+浓度下表现出比高Cd2+浓度下更高的Cd积累潜力[1],容易丧失食用价值,且水稻对Cd2+有较强的生理耐受能力和富集能力,因而水稻籽粒中的Cd积累量常常较高,成为“毒大米”。莫争等[2]研究发现,重金属中Cd2+在水稻中的转移能力最强,在水稻植株不同部位的积累分布依次为:根部>根基茎>主茎>穗>籽实叶部,而在Cd2+胁迫下会影响水稻对Ca2+、K+和Mg2+的吸收。许多离子如Mg2+、Ca2+、Mn2+、Fe2+、Cu2+和Zn+等可以抑制Cd2+在植物体内的转移[3-4],且外源添加N、Mg、P、S和Se能减轻Cd对某些酶和组织的有害影响[5],从而达到缓解Cd2+胁迫的作用;也有研究表明,Cd2+诱导的根毛细胞膜电位的去极化,不仅减弱了阳离子摄取的驱动力,还可部分阻断Ca2+和K+的内向电导,从而降低K+和Ca2+的摄取[6];但是否阻碍水稻对Mg2+的摄取需要证实。植物中Mg2+和Cd2+胁迫之间的关系复杂。研究表明,增加或减少水培植物的Mg2+供应可以减轻Cd的毒害[7];但在缺Mg2+条件下水稻幼苗的上部和根部Cd含量高于正常生长的水稻幼苗[8];也有研究认为,水稻种子在Cd2+胁迫下添加Mg2+诱导并促进植物生长使根系与芽中的Cd积累减少[9]。但从相关研究报道中很难得出关于Mg2+影响水稻吸收和转运Cd2+的一致性结论。
贵州属于高Cd地球化学背景区,土壤中Cd平均含量为0.659 mg·kg-1[11],且贵州大部分属喀斯特地区,土壤中富含Ca2+、Mg2+、CO32-/HCO3-、SO42-、NO2-等[12-14],特别是水中的Ca2+、Mg2+占主导地位[15-16]。田茂苑[17]研究发现,黄壤性水稻土和红壤性水稻土Cd污染风险格局与稻米Cd污染风险格局能够很好的重合,但石灰土性水稻土受Cd污染风险最大,而稻米受Cd污染的风险却较小,呈现出水稻土Cd污染风险格局与稻米Cd污染风险格局的不一致性。畅凯旋等[18]研究表明,喀斯特地区稻米Cd超标率较低,且喀斯特地区中性土壤Cd的有效态占比较低,从而降低水稻对Cd的吸收,但未指明喀斯特地区Mg2+对水稻Cd2+吸收的影响;也有研究表明,施用含Mg材料的调理剂与肥料能够降低水稻根系对Cd2+的吸收,可有效地降低水稻籽粒中Cd的含量[19-20]。因此本研究模拟喀斯特地区土壤与稻田灌溉水中不同浓度的Mg2+含量,进一步探讨Mg2+能否降低水稻苗期对Cd2+的吸收。目前研究大多集中在不同浓度Cd2+长期(15~42 d)胁迫下水稻Cd含量的差异[21-22],本研究采用本地常栽种的水稻品种[23]红优2号、黑糯72与杂交品种C两优华占、宜香优2115相对比,探究在短期胁迫(7 d)下水稻幼苗各部位Cd含量受不同浓度Mg2+影响的动态变化过程以及在两种Cd2+浓度胁迫下添加不同浓度Mg2+对水稻苗期根系Cd2+吸收及转运的影响,探讨在短期镉胁迫下添加外源Mg2+对水稻Cd2+吸收的机理,为非喀斯特地区施用含Mg2+材料修复治理不同程度Cd污染稻田土壤提供理论依据,也为贵州Cd地质高背景环境下筛选低富集Cd的水稻品种提供参考。
1 材料与方法
1.1 供试材料
供试水稻品种为籼稻:红优2号、宜香优2115、C两优华占;粳稻:黑糯72。其中红优2号与黑糯72为贵州本地选育品种,C两优华占为两系杂交品种,宜香优2115三系杂交品种;水稻种子与幼苗均由贵州省农业科学院水稻研究所提供。
1.2 试验设计
1.2.1水稻幼苗的培育
取籽粒饱满且大小相当的4个水稻品种种子若干,用10%的H2O2消毒浸泡15 min,用超纯水反复冲洗5~8遍后继续用超纯水浸泡24 h(期间换水4次),将种子均匀铺洒在滤纸上并放入28 ℃的恒温电热培育箱中避光催芽萌发,1 d后将露白萌发的种子转移至温度为25 ℃、相对湿度为95%的人工气候室内避光培养48 h。将水稻幼苗转移至装有1/4水培营养液[24]的水培箱中继续培养,幼苗间隔一定的距离,以保证有足够生长空间。待水稻幼苗长至3叶1心时选择出长势一致的水稻秧苗进行水培试验处理。
1.2.2水培试验
水培试验于2019年6月12日至2019年6月19日进行,水稻秧苗在贵州省农业科学院水稻研究所温室大棚中进行水培,植株在昼夜温度为35 ℃和20 ℃,相对湿度为60%~70%的避雨环境下生长。采用国际水稻研究所提供的水培营养配方培养,处理所用试剂为分析纯氯化镉(CdCl2)、氯化镁(MgCl2)。水培容器为1 000 mL棕色广口瓶,瓶口用定植棉固定。选择长势一致的3叶1心的水稻秧苗,用蒸馏水清洗根系后移栽至装有1 L营养液的广口瓶中,每瓶有4株幼苗,每2 d用NaOH与HCl调节pH值至5.75,设置2个Cd2+浓度(1 mg·kg-1、2 mg·kg-1)(简称为:Cd 1、Cd 2),4个Mg2+浓度(0、0.5、1、2 g·kg-1)(简称为:Mg 0、Mg 1、Mg 2、Mg 3),在2个Cd离子水平下添加4种不同浓度Mg2+培养,1个空白对照(Cd 0 Mg 0),共计9个处理(Cd 0 Mg 0,Cd 1 Mg 0,Cd 1 Mg 1,Cd 1 Mg 2,Cd 1 Mg 3,Cd 2 Mg 0,Cd 2 Mg 1,Cd 2 Mg 2,Cd 2 Mg 3),每个处理重复3次。每2 d更换一次营养液,每次更换营养液添加相应Mg2+与Cd2+处理,共换3次营养液,培养7 d。
图1 不同水稻品种幼苗胁迫第3天长势情况
1.3 样品采集与分析
在水稻苗期取水稻植株样品(每瓶取2株),先用自来水小心清洗根系,再将水稻根系用0.02 mol·L-1的EDTA溶液浸泡15 min,除去根表Cd2+,用超纯水冲洗整个水稻植株3次,吸干水分,然后将水稻根系、叶鞘、叶片分开,称取鲜重后在110 ℃下杀青再在70 ℃下烘干至恒重,粉碎并过100目筛。植物中Cd含量用HNO3-H2O2消解,用原子吸收分光光度计(ICE-3500)测定Cd含量,每个样品重复3次,同时质控样为芹菜标准物质(GBW 10048)。试验过程中均使用优级纯药品与超纯水。
1.4 数据处理
叶鞘-根系转移系数(TF叶鞘-根)=叶鞘中Cd含量/根系Cd含量;
叶片-叶鞘转移系数(TF叶片-叶鞘)=叶片中Cd含量/叶鞘Cd含量;
分配系数=(各器官Cd的累积量/整个植株Cd累积量)×100%。
试验数据为3次重复平均值,采用Microsoft Excel 2010软件和SPSS 22.0软件进行统计分析。
2 结果与分析
2.1 Mg对苗期水稻根系、叶鞘及叶片Cd2+吸收的影响
由图2可知,在Cd 1与Cd 2胁迫下,各品种水稻根系对Cd2+的吸收能力为:贵州本地水稻品种<杂交水稻品种,添加Mg2+对红优2号和C两优华占根系Cd2+吸收C两优在Cd 2中呈下降趋势,而黑糯72和宜香优2115根系对Cd2+的吸收则呈降低趋势,且黑糯72的根系对Cd2+的吸收能力比其他品种弱。在Cd 1与Cd 2浓度胁迫下,添加不同浓度Mg2+处理水稻根系对Cd2+吸收的差异均到达显著水平(p<0.05),在Cd 1 Mg 3处理时各品种水稻根系中的Cd含量大小为:C两优华占>红优2号>宜香优2115>黑糯72,与Cd 1 Mg 0相比分别下降25.66%、40.81%、69.97%及70.17%;在Cd 2 Mg 3处理下各品种水稻根系中Cd含量大小为:C两优华占>红优2号>黑糯72>宜香优2115,与Cd 2 Mg 0相比分别下降43.91%、51.27%、68.19%及76.65%。综上所述,Cd2+胁迫下添加Mg2+能够降低根系对Cd2+的吸收,且在高Cd2+浓度(Cd 2)水平下,相同Mg2+浓度处理,水稻根系中Cd2+含量下降幅度要比低Cd2+浓度水平(Cd 1)下的要大,即高Cd浓度胁迫下添加Mg2+使水稻根系中Cd含量下降效果最好。综上,Cd2+胁迫下添加Mg2+能够降低水稻幼苗根系对Cd2+的吸收。
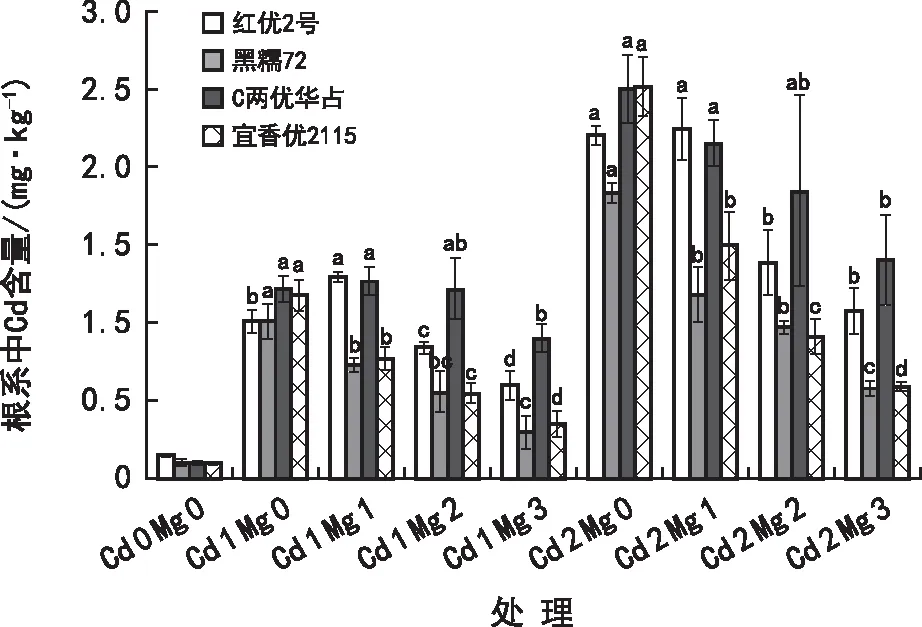
图2 在Cd2+胁迫下Mg2+对水稻幼苗根系Cd2+吸收的影响
由图3可知,在Cd 1和Cd 2胁迫下,各品种水稻叶鞘对Cd2+的吸收能力为:贵州本地水稻品种<杂交水稻品种,在Cd 1胁迫下,红优2号叶鞘中Cd含量随着Mg2+浓度的增加呈先增后降的趋势,其他品种Cd含量则随着Mg2+浓度的增加而逐渐减少。在Cd 1 Mg 3处理下各水稻叶鞘Cd含量大小为:C两优华占>宜香优2115>红优2号>黑糯72,与Cd 1 Ca 0相比分别降低47.37%、52.97%、33.30%、58.67%,在Cd 2 Mg 3处理下各水稻叶鞘Cd含量大小为:黑糯72>红优2号>宜香优2115>C两优华占,与Cd 2 Ca 0相比分别降低48.16%、48.13%、60.78%、75.51%。综上,Cd2+胁迫下添加Mg2+能够降低水稻叶鞘中Cd的含量,且在高Cd2+浓度水平(Cd 2)下,相同Mg2+浓度处理水稻叶鞘中Cd含量下降幅度要比低Cd2+浓度水平(Cd 1)下的大,即高Cd2+浓度胁迫下添加Mg2+水稻叶鞘中Cd含量下降效果最好。
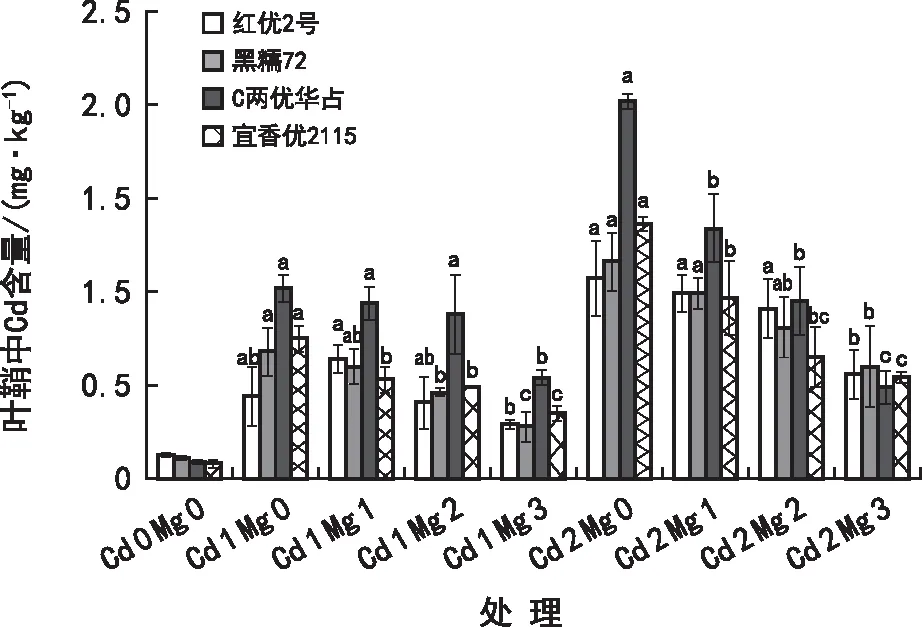
图3 在Cd2+胁迫下Mg2+对水稻幼苗叶鞘Cd2+吸收的影响
由图4可知,水稻叶片中Cd含量在Mg 1处理下本地品种低于杂交品种,但在Mg 2、Mg 3处理下各个水稻品种叶片中的Cd含量相差不大。在添加Mg2+后,各处理水稻叶片中Cd的含量与Cd 1 Mg 0、Cd 2 Mg 0相比均显著下降,且随着Mg2+浓度的增加而降低,分别在Cd 1 Mg 3与Cd 2 Mg 3时,下降幅度最大。在Cd 1 Mg 3处理下水稻叶片Cd含量:黑糯72>宜香优2115>C两优华占>红优2号,与Cd 1 Mg 0相比分别下降36.08%、60.59%、72.85%、53.22%;在Cd 2 Mg 3处理下水稻叶片Cd含量:黑糯72>宜香优2115>红优2号>C两优华占,与Cd 2 Mg 0相比分别下降56.31%、63.35%、52.67%、75.52%。综上,在Cd2+胁迫下Mg2+能够降低水稻叶片中Cd含量。在高浓度Cd2+(Cd 2)胁迫下添加Mg2+使水稻叶鞘中Cd含量下降效果最好。
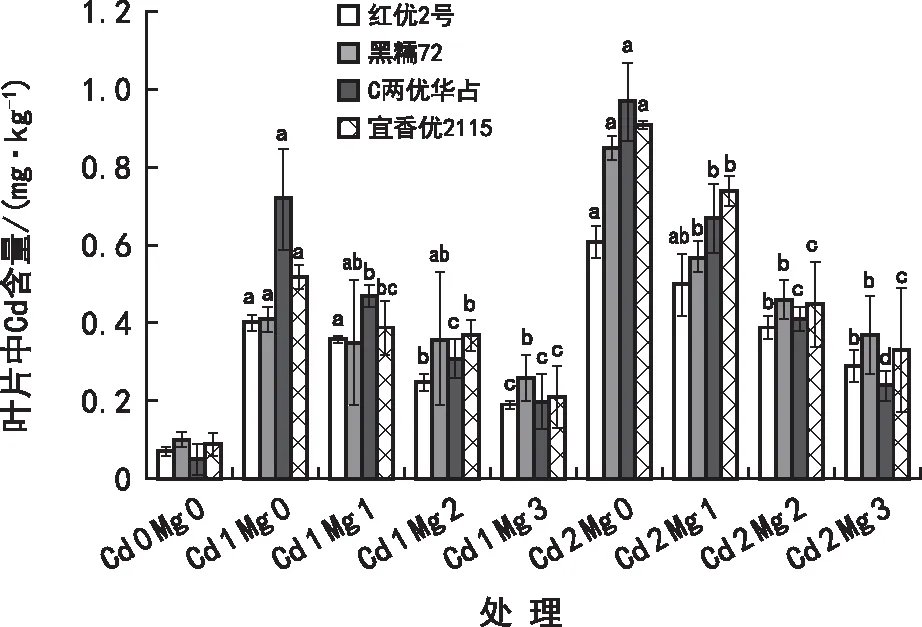
图4 在Cd2+胁迫下Mg2+对水稻幼苗叶片Cd2+吸收的影响
2.2 Mg对苗期水稻各器官Cd分配差异的影响
由图5可知,在两种不同浓度Cd2+的胁迫下,所有品种水稻苗期各器官Cd的分配系数表现为根>叶鞘>叶片,说明水稻植株吸收的Cd主要分配于根中(>40%),其次为叶鞘、叶片(<15%)。在Cd 1、Cd 2的胁迫下,随着Mg2+浓度的升高,水稻品种C两优华占的根部Cd分配比也逐渐升高,而地上部分的Cd的分配系数却呈下降趋势,说明植株内的Cd更多的积累于地下部;水稻品种红优2号在Cd 1胁迫下随着Mg2+浓度的升高,其根部Cd分配系数变化较为稳定,且在Cd 2胁迫下其根部Cd分配系数随着Mg2+浓度的升高变化不大,说明Mg2+对红优2号植株体内Cd的分配影响很小;但通过观察水稻品种黑糯72的Cd分配比例可以发现,在Cd 1、Cd 2的胁迫下,随着Mg2+浓度的升高,水稻根部的Cd分配比例呈下降趋势,说明植株内的Cd更多的积累于地上部。
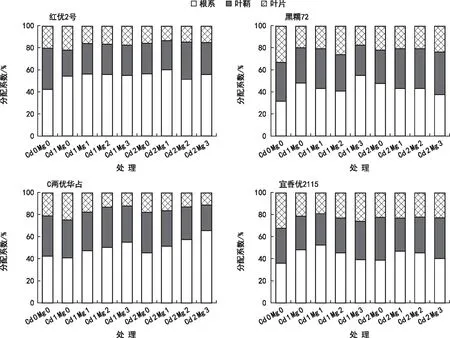
图5 4个水稻品种不同器官Cd的分配系数
本研究发现,在Cd胁迫下,黑糯72、宜香优2115和C两优华占根部Cd分配系数明显低于红优2号,但黑糯72叶片的Cd分配系数明显高于其他水稻品种叶片的Cd分配系数。不同水稻品种苗期所吸收并积累在植株中的Cd在各个器官的分配比例存在显著差异,产生这样的结果可能是不同的水稻品种对Cd胁迫的耐受度存在差异,即表现在水稻通过调节Cd在其各器官的分布以降低Cd对根部胁迫压力的能力差异上。
2.3 Mg对Cd在水稻苗期各器官转移系数的影响
重金属转移系数可以体现重金属从植物根部向地上部的转移特性,即反映出植株向地上各个器官运输重金属的能力,转移系数越大,说明重金属在植株中的转移能力越强。如表1、表2所示,在两种浓度Cd2+胁迫下,随着加入的Mg2+浓度增加,品种红优2号、黑糯72以及宜香优2115的叶鞘-根系转移系数呈递增趋势,在Cd 1胁迫下转移系数分别增加了13.63%、62.30%和57.81%;在Cd 2胁迫下转移系数分别增加了4.08%、66.7%和68.52%,而C两优华占则相反,各器官转移系数都在逐渐降低,在Cd 1和Cd 2胁迫下分别下降了40%和56.79%;在两种浓度Cd2+胁迫下,随着加入的Mg2+浓度增加,品种红优2号、C两优华占以及宜香优2115的叶片-叶鞘系转移系数呈递减趋势,在Cd 1胁迫下转移系数分别降低了35.35%、50.00%和14.29%;在Cd 2胁迫下转移系数分别降低了0.90%、12.00%和4.55%,而黑糯72在Cd 1胁迫下叶片-叶鞘系转移系数下增加了62.30%,在Cd 2胁迫下减少14.86%。
从表1、表2可以看出,各品种水稻幼苗的叶鞘-根系转移系数均大于叶片-叶鞘转移系数,说明Cd在水稻植株的转移中,Cd主要集中分布在水稻根部,在向地上部转移的过程当中,转移到水稻叶片部位的Cd要比转移到叶鞘中的少,且4个水稻品种中,黑糯72对Cd的转移能力最强,红优2号对Cd的转移能力最弱,也验证了上一节中Mg2+对红优2号植株体内Cd的分配影响很小,而黑糯72内的Cd更多的积累于地上部的结论。但Mg2+对水稻C两优华占Cd的转移影响最大,抑制了Cd2+由根系向地上部的转移。
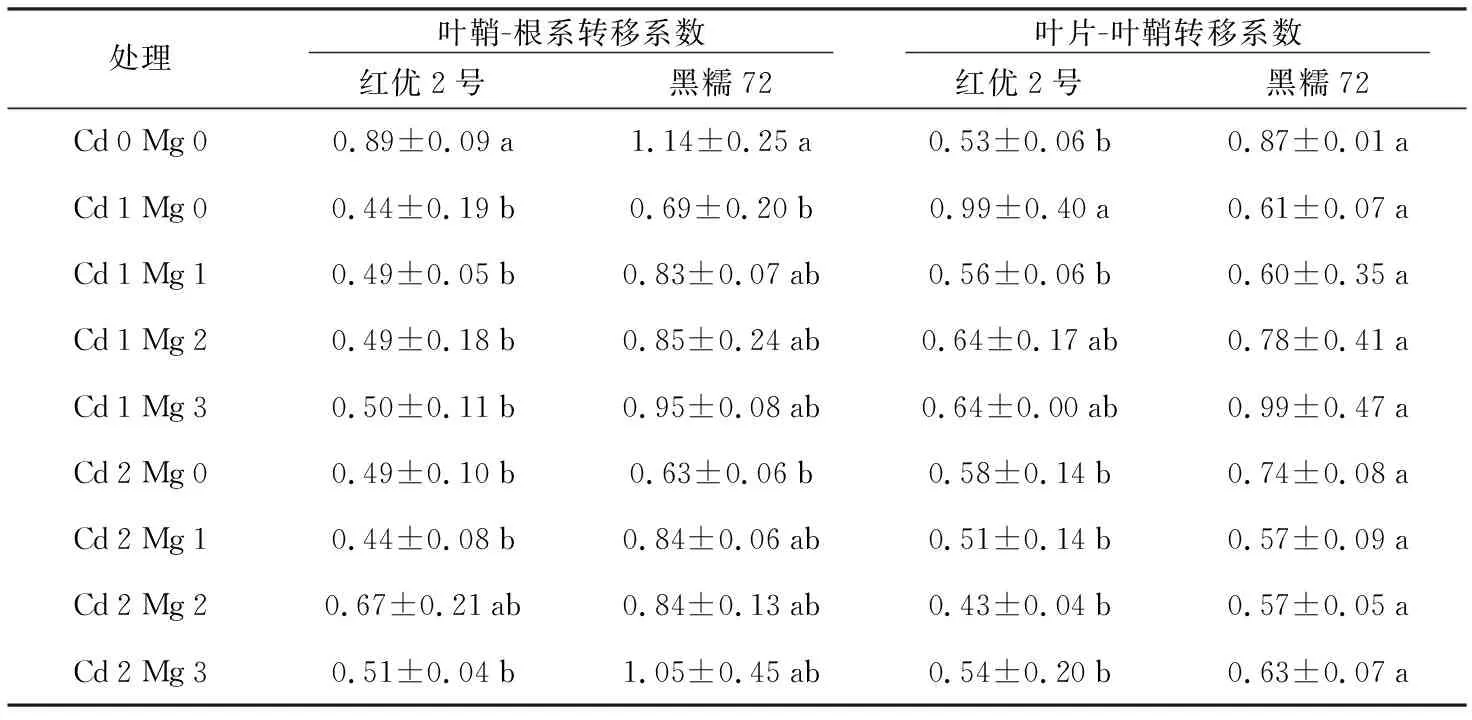
表1 红优2号与黑糯72不同器官Cd的转移系数
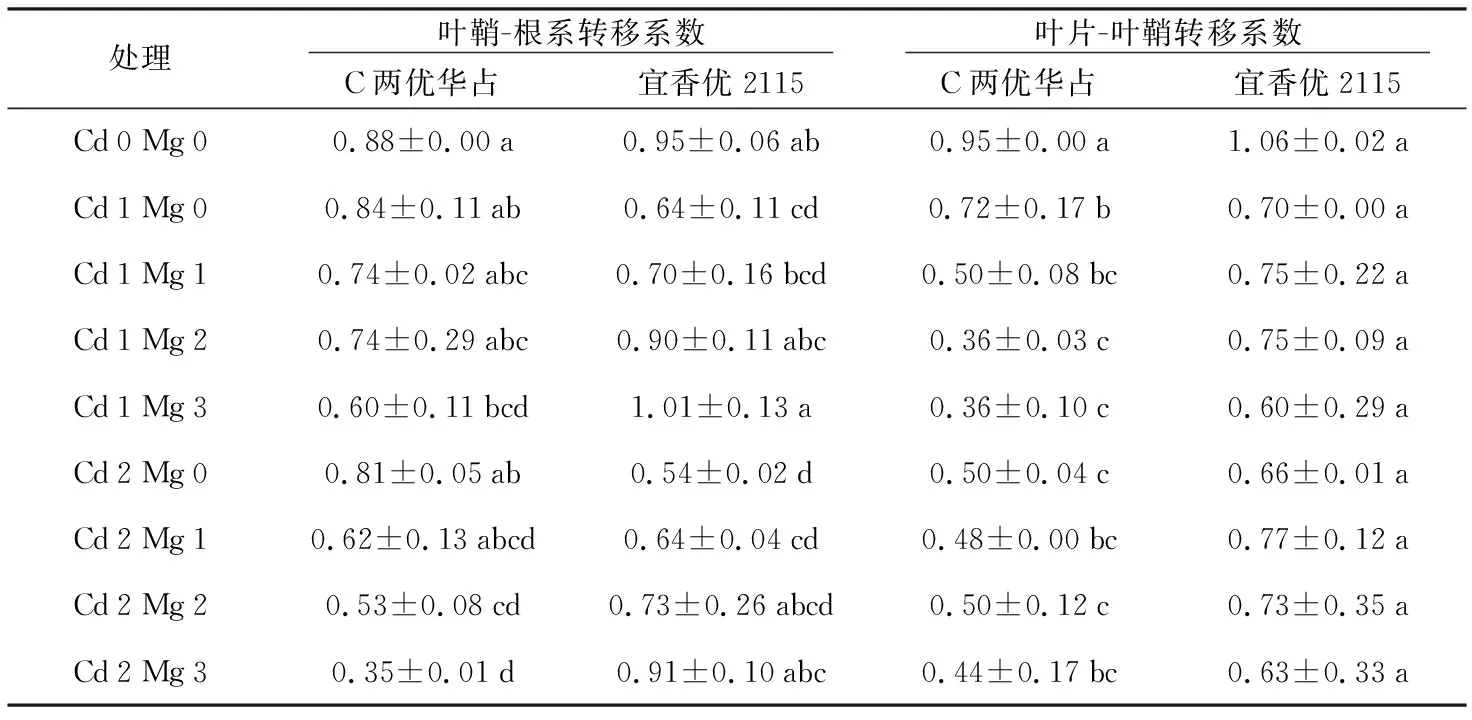
表2 C两优华占与宜香优2115不同器官Cd的转移系数
由表3可知,添加不同浓度的Cd2+,水稻根系与叶鞘中Cd积累量呈显著正相关,不同水稻品种间相关性大小关系为宜香优2115>黑糯72>红优2号>C两优华占,其中宜香优2115根系与叶鞘的相关系数最大(R2=0.966 9),说明添加Mg2+能有效促进宜香优2115根系中的Cd2+向叶鞘中转移,但对C两优华占(R2=0.806 9)根系中Cd2+向叶鞘中转移的促进作用最小。

表3 不同水稻品种根部与叶鞘中Cd含量相关性分析
2.4 Mg对苗期不同水稻品种各器官中Cd含量相关性
由表4可知,添加不同浓度的Cd2+,4个水稻品种的叶鞘与叶片中的Cd含量呈显著正相关,与水稻根系与叶鞘中Cd积累量呈显著正相关有一致性。不同水稻品种间的相关性强弱为宜香优2115(R2=0.978 7)>黑糯72(R2=0.911 6)>C两优华占(R2=0.895 9)>红优2号(R2=0.840 7),说明Mg2+对宜香优2115叶鞘中Cd向叶片中转移的促进作用最强,对C红优2号的促进作用最弱。由此得出,Mg2+能够增加宜香优2115叶鞘中Cd2+向叶片转移的移动性,增加水稻叶片中Cd的含量,Mg2+能有效促进宜香优2115中的Cd2+向叶片中移动性,增加了水稻地上部Cd风险,与其他水稻品种相比,Mg2+对C两优华占Cd2+移动的促进性最弱,使得C两优华占的大部分Cd积累在根系中,叶鞘与叶片中的Cd风险较小,也验证了红优2号对Cd2+的转移能力最弱的结论。

表4 不同水稻品种叶鞘与叶片中Cd含量相关性分析
3 讨 论
水稻对Cd2+的吸收主要通过质外体途径与共质体途径进入根系维管柱中,其中质外体途径属于被动运输,随着Cd2+浓度差异的增加,被动吸收越多[19-20],本试验结果与之一致;而共质体途径则属于主动运输,通过能量代谢使钙载体蛋白进入植物根系,最后通过胞间连丝进入根系维管柱中,需要消耗大量能量[21]。李延等[22]研究发现,水稻根、茎、叶不同部位的含Mg量与供Mg2+浓度呈正相关关系;Ding等[23]还发现,Mg2+具有较高的水化半径,Mg2+和其他阳离子如K+、Ca2+、Mn2+、和Al3+等有拮抗作用而产生阳离子竞争关系,可以推测Mg2+与Cd2+半径相近且电性相同,共同竞争水稻根系的离子吸附位点,从而降低水稻对Cd2+的吸收。
本试验中,在2种Cd2+浓度胁迫下通过外源增加Mg2+可显著降低Cd2+在水稻幼苗根系和地上部器官中的含量,因此推测在Cd胁迫下Mg2+降低水稻对Cd2+的吸收能力主要有3个原因,第一是Cd2+与Mg2+相互竞争能量,Cd2+和Mg2+在主动运输过程中消耗了大量能量从而减少了对镉的吸收;第二是Cd2+与Mg2+竞争离子吸附点,Mg2+与Cd2+因半径相近电性相同,共同竞争水稻根系离子吸附位点,降低水稻对Cd2+的吸收;第三是Cd与Mg竞争非选择性阳离子通道,减少对Cd2+的吸收并且减弱Cd在水稻各器官中的转移能力[30]。
在2种Cd2+浓度胁迫下,苗期中4个品种水稻的各器官对Cd的富集量的大小表现为:根系>叶鞘>叶片,这与赵步洪[25],范中亮等[26]的研究结果相一致,且水稻根系中所积累的Cd含量占水稻所吸收Cd2+总量的45%~90%[27],但不同浓度Cd2+处理下,水稻各器官中Cd含量在品种间存在显著差异[34]。
Liu J等[35]研究表明,籼稻茎叶和籽粒中Cd含量普遍高于粳稻,其中糙米中Cd含量呈现籼型杂交稻>粳型杂交稻>常规籼稻>常规粳稻[36]。说明水稻品种对Cd2+的吸收能力为杂交稻>常规稻,籼稻>粳稻,本研究的结果与之一致,说明不同水稻品种对Cd的耐受性存在差异。本研究还发现,外地杂交水稻品种在相同Cd2+浓度胁迫下比贵州本地水稻品种各部位Cd含量高,本地品种水稻根系Cd含量为红优2号>黑糯72,但黑糯72叶鞘与叶片中的Cd含量均为最高,说明在黑糯72根系中Cd易向地上部位转移,杂交品种根系、叶鞘及叶片Cd含量均显示C宜香优2115>C两优华占,说明各水稻品种之间Cd的吸收分配特点有差异。此外,还发现了一个相关联的因果关系:Mg2+对杂交水稻C两优华占Cd2+移动的促进性最弱,且在Cd2+胁迫下添加Mg2+后根系的分配比例提高,使得大部分Cd积累在根系中,也导致Cd2+从根系到地上部的转移能力减弱,即叶鞘-根系、叶片-叶鞘转移系数在逐渐下降,从而该品种水稻叶鞘与叶片中的Cd含量相较于其他三个水稻品种最少,地上部Cd风险也最小。本研究还发现,水稻各器官Cd转移系数与Mg处理浓度无显著相关性,这与龙小林等[37]的结论相一致,说明Cd向叶片的转运能力主要是由水稻品种的基因型决定,受Cd2+胁迫等环境因素影响较小。
4 结 论
在Cd2+胁迫下,Mg2+能够显著降低水稻根系、叶鞘及叶片中Cd含量,且在添加2 mg·kg-1浓度的Mg2+处理下降低Cd含量的效果最为显著。在Cd2+胁迫下各水稻品种对Cd的吸收能力有明显差异,吸收强弱关系为:C两优华占>红优2号>宜香优2115>黑糯72,杂交品种比本地品种对Cd的吸收能力强。水稻根系、叶鞘及叶片中Cd含量关系为:根系>叶鞘>叶片,且根系对叶鞘及叶片中Cd与Mg含量均有显著影响。
猜你喜欢
现代园艺(2022年7期)2022-11-19
蔬菜(2022年8期)2022-08-25
今日农业(2021年10期)2021-11-27
今日农业(2021年9期)2021-11-26
诗潮(2021年1期)2021-03-15
湖北农业科学(2019年22期)2019-12-23
农家致富顾问·下半月(2019年7期)2019-09-28
农村百事通(2019年8期)2019-05-13
现代农业研究(2009年9期)2009-10-21
作文新天地(高中版)(2009年2期)2009-01-22
