
Characterization of metabolic landscape in hepatocellular carcinoma
2021-09-18 02:14:40JingWuRanXueRongTaoJiangQingHuaMeng
Jing Wu,Ran Xue,Rong-Tao Jiang,Qing-Hua Meng
Jing Wu,Qing-Hua Meng,Department of Medical Oncology,Beijing You-An Hospital,Capital Medical University,Beijing 100069,China
Ran Xue,Key Laboratory of Carcinogenesis and Translational Research(Ministry of Education),Department of Gastrointestinal Oncology,Peking University Cancer Hospital &Institute,Beijing 100036,China
Rong-Tao Jiang,National Laboratory of Pattern Recognition,Institute of Automation,Chinese Academy of Sciences,Beijing 100190,China
Abstract Hepatocellular carcinoma(HCC)is one of the most prevalent cancers worldwide,accounting for approximately 75%-85% of primary liver cancers.Metabolic alterations have been labeled as an emerging hallmark of tumors.Specially,the last decades have registered a significant improvement in our understanding of the role of metabolism in driving the carcinogenesis and progression of HCC.In this paper,we provide a review of recent studies that investigated the metabolic traits of HCC with a specific focus on three common metabolic alterations involving glycolysis,lipid metabolism,and glutamine addiction which have been gaining much attention in the field of HCC.Next,we describe some representative diagnostic markers or tools,and promising treatment agents that are proposed on the basis of the aforementioned metabolic alterations for HCC.Finally,we present some challenges and directions that may promisingly speed up the process of developing objective diagnostic markers and therapeutic options underlying HCC.Specifically,we recommend future investigations to carefully take into account the influence of heterogeneity,control for study-specific confounds,and invite the validation of existing biomarkers.
Key Words:Hepatocellular carcinoma;Metabolic reprogramming;Glycolysis;Lipid metabolism;Glutamine addiction;Diagnosis and treatment
INTRODUCTION
Liver cancer has been one of the most prevalent cancers worldwide,and is the third leading cause of cancer mortality following lung cancer and colorectal cancer,according to the GLOBOCAN estimates for 2020[1].Hepatocellular carcinoma(HCC)accounts for approximately 75%-85% of primary liver cancers.Chronic hepatitis B/C infection,alcohol abuse,aflatoxin contamination,and obesity stand out as the major risk factors for HCC[2].Due to the lack of symptoms and signs in early stages,most patients are diagnosed at an advanced stage without a satisfactory curative option.Although recent advances in the early diagnosis and treatment have increased the short-time survival of HCC patients,the long-time prognosis remains disappointing.Among all available treatment options for patients with advanced HCC,sorafenib has been the first-line systemic treatment for unresectable cases since 2005[3].However,drug resistance has significantly limited its clinical efficiency and availability.There is an urgent need to better understand the underlying mechanisms that drive the carcinogenesis and progression of HCC,and more importantly,to further explore effective and precise treatment for HCC patients.
Over the last decades,metabolic alterations have been labeled as an emerging hallmark of tumors,where tumor cells exhibit the propensity to consistently fuel glucose to produce lactate even in the presence of sufficient oxygen,a state named“aerobic glycolysis”[4,5].Cell-autonomous redirecting of metabolism could exert cancer-promoting functions,which meets the increased demands for energy and metabolic substrates of cancer cells to sustain rapid cellular proliferation.Recently,with the advent of DNA-sequencing technologies as well as proteomics and metabolomics analyses,the identification of disease-specific patterns of genetic mutations and metabolic profile becomes possible.Since the 1920s when aerobic glycolysis was first recognized in rat liver cancer,there has been an explosion of research aiming at exploring the metabolism of HCC,which is making rapid progress[5].Prospectively,a deep understanding of the metabolic transitions in HCC would confer advantages not only for early diagnosis but also for effective treatment,bringing us closer to the goal of developing objective diagnostic markers and therapeutic options.
In this review,we will focus on characterizing the metabolic traits of HCC.Specifically,we outline three common metabolic alterations driving the carcinogenesis and progression of HCC,including glycolysis,lipid metabolism,and glutamine addiction,among which,glycolysis and glutamine addiction have been extensively studied over the last decades,while lipid metabolism has grown in popularity in recent years.Then,we elaborate on the diagnostic markers or tools,as well as some promising treatment agents that are proposed on the basis of the aforementioned metabolic alterations for HCC.Finally,challenges related to the existence of great heterogeneity in HCC,the ignorance of study-related confounders,and a lack of validation of proposed markers are discussed.
METABOLIC ALTERATIONS IN HCC
Aerobic glycolysis in HCC
It has been reported that glucose is channeled into glycolysis rather than oxidative phosphorylation(OXPHOS)in many cancer types including HCC.The aerobic glycolysis constitutes a suitable mechanism that exerts beneficial effects on tumor proliferation,immune evasion,metastasis,angiogenesis,and drug resistance[6,7].Although ineffective in adenosine triphosphate(ATP)production,the process of aerobic glycolysis is more rapid and does provide a mass of metabolic intermediates that are building blocks for the synthesis of biomicromolecules[8].A study using nontargeted tissue metabolomics indicated that increased glycolysis constituted one of the core alterations of metabolic features of HCC,and the enriched glycolysis-related metabolites were related to energy supply[9].
The upregulation of key enzymes and transporters in aerobic glycolysis is the critical player in promoting glycolytic flux.Independent lines of research suggest that hexokinase-2(HK2),the first rate-limiting enzyme in aerobic glycolysis,is highly expressed in HCC,which predicts a higher risk of poor prognosis.Targeting HK2 could suppress glucose metabolism and reverse the malignant phenotype of HCC[10,11].Notably,recent studies suggested that HK2 depletion-induced glycolysis shutdown improved the sensitivity of HCC cells to metformin and facilitated the toxicity of sorafenib[12].Phosphofructokinase-1(PFK1)is the second rate-limiting enzyme catalyzing the formation of fructose-1,6-bisphosphate from the precursor fructose-6-phosphate in glycolysis.Specifically,a study conducted by Huet al[13]observed that metformin can moderate hepatoma cell proliferationviainhibiting glycolysis flux through the regulation of the hypoxia-inducible factor 1α(HIF-1α)/PFKFB3/PFK1 pathway in HCC,implying the potential utility of metformin as a cancer therapeutic drug[13].Generally,there are three isoforms of PFK1 involving PKFL(liver type),PKFM(mainly expressed in muscle),and PKFP(platelet isoform)[14],which are frequently upregulated in many tumor tissues including HCC.Growing evidence suggests that PKFL or PKFP functions as a downstream target of several genes that could affect the glycolytic flux to suppress HCC progression[15-17].Pyruvate kinase(PK)is the third rate-limiting enzyme catalyzing the last step of glycolysis.PK has four isoforms including PKL,PKR,PKM1,and PKM2,among which convergent findings suggest that PKM2 is highly expressed and related to decreased overall survival in HCC patients[18].PKM2 is targeted by other genes to restrain tumorigenicity and re-sensitize HCC cells to sorafenib[19].Epigenetic modification and posttranscriptional modification of PKM2 are the critical mechanisms through which upstream genes affect the stability and degradation of PKM2 to further affect the glucose metabolism[18,19].
Apart from the aforementioned enzymes,accumulating research has discovered the aberrant expression of the family of glucose transporters(GLUT1-4)driving the initial step of glycolysis in different cancers.Specifically,GLUT1 and GLUT2 are the most widely reported proteins overexpressed in HCC,and studies reported that patients with higher GLUT1 expression were associated with a worse prognosis,which are manifested by higher recurrence rates[20].In addition to these enzymes and transporters that directly participate in glycolytic flux,the lactate dehydrogenase and monocarboxylate transporters that are associated with the ongoing glycolytic flux are also found to promote the tumor growth and progression of HCC[21,22].
Despite these promising findings of the potential functions of these enzymes and transporters in HCC,a deeper investigation of these enzymes and their interplay with other genes is needed,which may raise the possibility of defining diagnostic biomarkers and therapeutic targets in the long run.
Lipid metabolism in HCC
Lipid metabolic abnormalities have been attracting increasing interest not only because the lipid plays a critical role in the survival of cancer cells but also because there are accumulating studies indicating that lipid metabolic alterations are a driving force in the development and progression of multiple cancers including HCC[9,23].Growing evidence suggests that increasedde novosynthesis of lipids and desaturation are responsible for HCC tumorigenesis and progression[24].As lipid metabolism is also a process driven by enzymatic reaction,the enzymes and metabolites involved in lipid metabolism are a major focus of research.Specifically,fatty acid(FA)synthase(FASN),ATP citrate lyase,acetyl CoA carboxylase,and stearoyl-Coenzyme A desaturase-1 have been found to be universally upregulated in cancers.Genetic or pharmacological inhibition of these proteins contributes to repression of HCC and mitigation of sorafenib resistance[25,26].
Relative to the consistence of studies investigating FA synthesis in HCC,findings about FA oxidation(FAO)are far from consistent.In most conditions,FAO is inhibited by downregulation of key enzymes to protect cells from oxidative damage and lipotoxic cell death(Figure 1A).For example,in steatohepatitic HCC or nonalcoholic steatohepatitis-driven HCC,carnitine palmitoyltransferase 2(CPT2),which can convert acylcarnitine to AcylCoA,is frequently downregulated and further inhibits the FAO by restraining the Src-mediated JNK activation to adapt to lipid-rich conditions[27].In line with the above findings,under hypoxia conditions,HIF-1α can facilitate HCC progression through HIF-1α-mediated FAO blocking[28].With regard to antiangiogenic drug(AAD)-induced hypoxic conditions,it is another scenario,where elevated frontal fibrosing alopecia and FAO can promote tumor proliferation and drug resistanceviaactivating the adenosine monophosphate-activated protein kinase(AMPK)pathway,while loss of CPT1 function could improve AAD therapeutic efficacy and its anti-tumor effects[29].In accordance with this finding,Luet al[30]found that CCAAT/enhancer binding protein α-mediated autophagy can trigger lipid catabolism upon glucose and glutamine double starvation regardless of normoxia or hypoxia[30].As is well known,25%–40% of HCCs harboring aberrant β-catenin activation exhibit a lower proliferation index but a considerably distinctive metabolic morphotype[31,32].Using a mouse model recapitulating β-catenin-mutated HCC,Senniet al[31]observed the overexpression of peroxisome proliferator activated receptor alpha(PPARα)and CPT2,and further uncovered a peculiar metabolic landscape that FAO rather than glycolysis was the predominant energy supplier(Figure 1B).Mechanistic studies implied that it is PPARα that orchestrates this metabolic rewiring and FAO is an indispensable driving force in β-catenin-activated HCC,highlighting the emerging strategies to intervene in PPARα and FAO for therapeutic benefit.Taken together,the discrepancy mentioned above reflects,to some extent,the complexity and heterogeneity of HCC,and detailed mechanisms of lipid metabolism underlying HCC progression remain to be further elucidated.
Glutamine metabolism in HCC
In addition to glucose,glutamine is another widely recognized nutrient being preferentially used by cancer cells to sustain rapid proliferation,which is known as glutamine addiction.Glutamine serves as a carbon and nitrogen donor for protein and nucleotides synthesis.In the perspective of energy metabolism,glutamine catabolism promotes ATP production by providing tricarboxylic acid cycle intermediates.In addition,the glutathione,one of the products of glutamine,confers selective advantage for tumor survival during the oxidative stress[33].All of the above aspects together constitute the phenotype of glutamine dependency.Consistent with this,transporters and enzymes involved in glutamine metabolism are frequently upregulated in multiple tumors including HCC[34].SLC1A5,also known as ASCT-2 that is responsible for glutamine uptake,is highly expressed in HCC tissues and implies a poor prognosis.Inhibiting SLC1A5 could impede HCC growth and increase the sensitivity of HCC cells to chemotherapeutic drugs[34].The most studied enzyme of glutamine metabolism is the glutaminase isoenzyme GLS1,which is also overexpressed in HCC and portends poor outcome[35].Intriguingly,the results from Jinet al[35]suggested that inhibiting GLS1 efficiently blocks glutamine consumption but is insufficient for tumor repression.In contrast,dual inhibition of GLS1 and SLC1A5 has a synergistic effect on growth arrest in HCC owning to lethal oxidative damage and apoptosis induction[35].
Apart from these transporters and enzymes,glutamine metabolism is also under oncogenic control,of which MYC and Wnt-β-catenin pathways have received the widest attention(Figure 2).As mentioned above,HCC cells bearing consistent βcatenin activation have distinct metabolic alterations.Generally,glutamine synthetase(GS)is exclusively expressed in hepatocytes around the central vein.In HCC with consistent β-catenin activation background,GS is supposed to be a target of β-catenin and is strikingly overexpressed,while in HCC without deregulated β-catenin signaling,GLS1 is commonly upregulated[36].Increased GS is correlated with mTORC1 activation and further contributes to glutamine accumulation[36](Figure 2B).Inhibiting GS could not only hinder tumor growth but also abrogate drug resistance.Thus,repressing glutamine-dependent mTORC1 activation may represent a novel therapeutic strategy for HCC patients[37].In MYC-driven liver tumors,glutamine is the dominant fuel for TCA,and GLS1 is overexpressed,accompanied by low expression of GS(Figure 2A).It is tempting to speculate that MYC may have a synthetic lethal effect with GLS1 inhibitor or glutamine depletion on growth inhibition[38].Of note,although glutamine addiction is a quasi-universal finding,some studies also indicated that it does not hold good in all cases,given the fact that certain cells including the well-differentiated HCC cell line Huh7 can proliferate independently of extracellular glutamine[39].Overall,targeting glutamine catabolism as a metabolic vulnerability holds great potentials in the treatment of HCC,and more research is needed to determine which tumors are most glutamine-dependent.
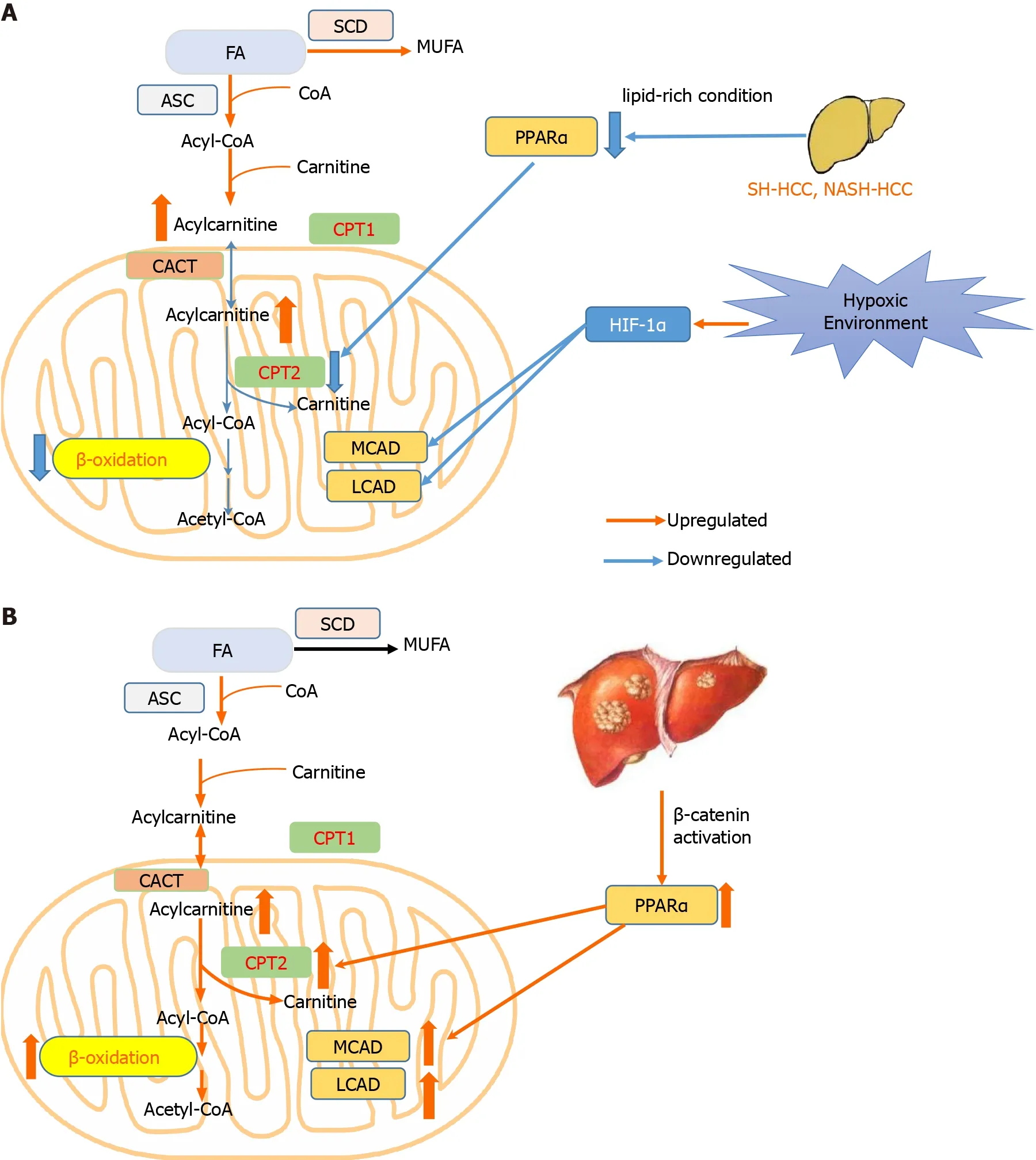
Figure 1 Hepatocellular carcinoma develops two distinct lipid metabolism patterns depending on the environment or genetic background of tumor cells.A:In most conditions including hypoxia environment,and steatohepatitic or nonalcoholic steato-hepatitis-driven hepatocellular carcinoma(HCC),βoxidation is repressed to avoid excessive oxidative damage;B:In HCC harboring β-catenin mutation,β-oxidation is promoted to trigger HCC.CACT:Carnitine–acylcarnitine translocase;CPT1:Carnitine palmitoyltransferase 1;CPT2:Carnitine palmitoyltransferase 2;FA:Fatty acid;HCC:Hepatocellular carcinoma;HIF-1α:Hypoxia inducible factor 1 subunit alpha;LCAD:Long-chain acyl-CoA dehydrogenases;MCAD:Medium-chain acyl-CoA dehydrogenases;MUFA:Monounsaturated fatty acid;PPARα:Peroxisome proliferator activated receptor alpha;SCD:Stearoyl-CoA desaturase.

Figure 2 Two distinct principles governing glutamine metabolism in hepatocellular carcinoma.A:Glutaminolysis in MYC-induced hepatocellular carcinoma(HCC)is enhanced via increasing transport and catabolism to fuel Krebs cycle;B:In HCC bearing β-catenin mutation,glutamine accumulation in response to glutamine synthetase upregulation further activates phosphorylated mammalian target of rapamycin.GLS1:Kidney-type glutaminase;GLS2:Liver type glutaminase;Gln:Glutamine;GS:Glutamine synthetase;SLC1A5:Solute-linked carrier family A1 member 5;p-4EBP1:Phosphorylated 4E-binding protein 1;pmTOR:Phosphorylated mammalian target of rapamycin.
METABOLISM-BASED DIAGNOSIS AND TREATMENT
Metabolic rewiring has been a perpetual topic of interest for decades.As the field continues to progress,tremendous effort has been devoted to identifying predictive biomarkers and agents that hold great promise for early tumor diagnosis and treatment(Table 1 and Supplementary Table 1).We now review these diagnostic tools or markers and treatment agents.
Diagnosis
The limitations of traditional tools[alpha-fetoprotein(AFP)and ultrasound]for HCC diagnosis significantly restrict their diagnostic ability,thereby entailing other tools or biomarkers with high specificity and sensitivity.In recent years,there is an emerging trend in using certain metabolic radiotracers in combination with positron emission tomography/computed tomography(PET/CT)to image tumors and provide prognostic information(Table 1).For example,F-18 fluorodeoxyglucose(18F-FDG,an indicator for glycolysis)PET/CT images can be used to non-invasively monitor the development of HCC in a rat model[40].Moreover,they are also commonly studied as predictors of overall survival and tumor recurrence after related treatments in HCC[41,42].However,the fact that normal cells in the heart,brain,etc.also consume a large amount of glucose may inevitably increase the false-positive results.Simultaneously,the false-negative detection also increases in virtue of low FDG uptake by well-differentiated HCC.Alternatively,other investigational PET agents exert a supplementary role to18F-FDG.Specifically,C-11 acetate(a surrogate for FA synthesis)PET/CT may help detect glycolysis-independent HCC and metastatic sites[43,44].Likewise,(4S)-4-(3-[18F]fluoropropyl)-L-glutamate,namely,[18F]FSPG,has been investigated both in animal and human studies[45].In HCC,[18F]FSPG has a detection rate of 100%,compared with 60% of18F-FDG.Typically,the accumulation of[18F]FSPG is significantly lower than that of18F-FDG in heathy liver[46],indicating the potential diagnostic value of[18F]FSPG in HCC.
Apart from the above imaging-based tools,some metabolites also show some promise as diagnostic biomarkers[47].Metabolomics is emerging as a convenient and potent tool that opens interesting cues to systematically explore diagnostic biomarker candidates and reveal the underlying mechanisms of tumorgenicity.To date,many studies have applied serum,urine,and feces for assessing metabolite profile in HCC[48-50].As different metabolic activities can coexist simultaneously in complex physiopathologic contexts,studies always find out a combination of a wide repertoire of metabolites that may have diagnostic value in clinical practice[51,52](Table 1).Specifically,metabolomics analysis of HCC tissues and serum specimens discovered that combining betaine and propionyl-carnitine together can differentiate HCC from cirrhosis and chronic hepatitis[9],which concurs with prior evidence provided by Zhouet al[53].Another study identified a panel of 11 metabolites and three clinical covariates with LASSO regression,and diagnostic model constructed using these features and support vector machines achieved a high classification accuracy in distinguishing HCC from liver cirrhosis[52].Likely,Wanget al[54]also acquired 13metabolites using partial least-squares-latent structure discriminate analysis,which can discriminate HCC from healthy controls and liver cirrhosis[54].In spite of the above exciting results,these potential biomarkers have not been integrated into clinical realm.One major reason is that there is still a scarcity of large population and multicentral-based validation of these biomarkers.Consequently,many efforts are required before these biomarkers can be translated into clinical utility.

Table 1 Summary of metabolism-based diagnostic tools or markers and treatment agents
Treatment
To date,there have been some existing and investigational metabolism-targeted drugs(Table 1).After being phosphorylated by HK2,2-deoxy-D-glucose(2-DG),a glucose analog,could noncompetitively repress HK2 and thus decrease the glycolytic cascade.Moreover,2-DG is able to hinder tumor progression by inducing apoptosis[55].Combining 2-DG with other genes or proteins can not only inhibit tumor growth but also reverse sorafenib resistance in HCC[56,57].In the ClinicalTrials.gov results database,there exist some completed clinical trials(such as NCT00096707 and NCT00633087)focusing on the safety and efficiency of 2-DG in advanced cancers,which would provide valuable cues for its clinical utilization.3-bromopyruvate(3-BP)is another antiglycolytic agent that functionsviadirectly restraining HK2.3-BP was first reported to have potential therapeutic value in cancers by Koet al[58].A study conducted in rabbits recapitulated that 3-BP was lethal to both primary liver tumor and metastatic tumors arising from the lungs,but without any toxic side effects.Other investigations attributed the tumor inhibition of 3-BP to its functions in inducing apoptosis and destructing cellular antioxidative defense[58].In addition,there has been evidence suggesting that 3-BP is a potent inhibitor of several other molecules including PK and ribonuclease A,confirming the multi-faceted properties of 3-BP in mediating toxicity in cancer cells[59].Taking the above meaningful results into consideration,3-BP was eventually approved by the Food and Drug Administration.
Metformin has been used as a first-line anti-diabetic drug;however,its anti-cancer effect has attracted numerous attention recently.In mitochondria,metformin can inhibit complex I and result in ATP reduction,which further activates AMPK,which is known as an energy stress sensor.Moreover,metformin participates in various biological processes including inflammatory response,insulin regulation,immune response,mTORC1 activation,and folate metabolism,all of which are associated with its anti-tumor action[60,61].Similarly,searching on clinicaltrials.gov with “Metformin”and “Tumor” or “Neoplasm” led to the retrieval of hundreds of completed or ongoing trials about the anti-tumor effect of metformin(some of the trials are shown in Supplementary Table 1).Other agents like TVB-2640,a reversible inhibitor of FASN,have also progressed to human clinical trials[62].These existing preclinical experiments are promising in optimizing the clinical response in HCC treatment.
Apart from tumor diagnosis and treatment,the metabolism can also be used to classify HCCs into subtypes.A study proposed three subclasses of HCC with active,intermediate,or exhausted metabolic activities,which not only verified the heterogeneity of HCC from the metabolic landscape,but also implied possible prognosis and potential response toward metabolic therapies[63].
CHALLENGES AND FUTURE DIRECTIONS
The advent of proteomics and metabolomics facilitates our understanding of metabolic landscape and mechanisms underlying tumorigenesis,which would generate great optimism in cancer therapy based on metabolic vulnerability.The studies surveyed above do imply that we have made preliminary progress in revealing the metabolism perturbation and providing opportunities for the diagnosis and treatment of HCC.Despite such success,some crucial issues should be discussed and circumvented in future investigations.Here we elucidate some challenges and provide some potential future directions.
Great metabolic heterogeneity of HCC
There is a growing consensus that HCC is one of the most heterogeneous cancers that is characterized by morphological multiformity,varied signal transduction network perturbation,and microenvironmental discrepancies[64].An abundance of research has been devoted to investigating the metabolism profile of cancers,and there is ample evidence suggesting that there exists great metabolism heterogeneity in HCC[65].Indeed,the heterogeneity of HCC could at least partially account for the inconsistent findings in metabolic profiles,such as the difference of metabolic phenotype between HCC patients bearing β-catenin mutation and those without this mutation[31,32].This is of extraordinary importance for selecting certain patients for a specific therapeutic option.As a concrete example,a randomized,double-blind,phase 3 study assessing the efficacy of Everolimus(an mTOR inhibitor)vsplacebo in advanced HCC patients yielded negative results,partially owing to the fact that this study enrolled in “all comers” without selecting susceptible patients with mTOR-addicted tumors[66].
In addition,the metabolic zonation of the liver can impose further complexity and heterogeneity when designing a metabolic study[67].For example,in parallel with the fact that 50% of liver genes are significantly zonated,certain metabolic processes are also distributed zonally with lipogenesis being much higher in the central side while gluconeogenesis and FAO dominating in the portal side[68].Finally,asin vitrostudies are indispensable for providing preliminary cues for translational studies,the selection of distinct experimental models is also a significant contributor to discordant results.Specifically,for HCC,poorly-differentiated cell lines like HLE most resemble the clinically-relevant HCC metabolism,while well-differentiated cell lines including Huh7 may resemble that existing in the early stage of liver cancer[39].Consequently,all the above factors may plague various forms of experimental results,and the influence of heterogeneity should be taken into account before conducting a metabolic study.
Study-related confounders
First of all,many of the basic metabolic studies focus on the expression and activities of pivotal enzymes of certain metabolism,which frequently suggest that intervention of these related enzymes would impede tumor growth.Indeed,there are limited beneficial effects because the altered metabolism may just be a consequence of HCC genetic lesion.Therefore,intervening related genes is the root to prevent tumor initiation and progression.A case in point was that in c-Myc-mediated energy alterations,the combination of sorafenib or regorafenib and c-Myc inhibitor demonstrated a significant tumor inhibition effect,compared with the combination with 2-DG or BPTES(a GLS1 inhibitor)[69].Meanwhile,as different metabolic types frequently coexist simultaneously in the complex physiopathologic contexts of HCC(such as glycolysis and glutaminolysis),a comprehensive understanding of metabolic perturbation is fundamental for an efficient treatment option.
Second,for clinical-related investigations,heterogeneity of sample properties between compared groups is a confusing factor and propensity matching of the irrelevant variables(e.g.,gender,age,body mass index)that may affect the result should be ensured in order to avoid misleading results[70].Importantly,obesity and diabetes are two common factors that are remarkably related to personal metabolism,incorporation of which inevitably confuses researchers.Third,heterogeneity across studies is another considerable confounder.Specifically,these studies are usually varied across a series of key aspects including tumor stage,degree of cirrhosis or fibrosis,etiology,etc.As a prominent example,the metabolic phenotype of HCC infected with hepatitis B or hepatitis C virus significantly differs from HCC cases with alcoholic cirrhosis,consequently incorporation of HCC patients with a mixture etiology of hepatitis B and alcoholic cirrhosis can bias results[50,71].
Finally,the field also calls for studies performed in large-sample cohorts since small samples cannot fully represent the entire spectrum of a population although they are flexible in data analyses.Moreover,studies leveraging small samples are in danger of yielding false-positive results[72].
Lack of validation of proposed markers
To date,a number of metabolic studies have been devoted to identifying specific biomarkers for early HCC diagnosis.These biomarkers,however,have not been translated into concrete improvements in real-world settings.One of the main reasons comes from the fact that most of these studies have focused on discovering new findings instead of validating them.Consequently,to what extent the markers developed from experiments can be effectively applied in clinical remains largely unknown.The next phase of cancer metabolism research should shift form marker discovery to robust validation[47],which can promisingly speed up the process of applying the proposed markers in clinical practice[73].Furthermore,instead of focusing on using traditional group-level comparisons to detect whether significant difference exists between HCC and healthy controls or other liver diseases for specific metabolites,advanced machine learning models can be built to identify metabolites that can diagnose HCC at the individual level[70].
CONCLUSION
In this paper,we review the recent advances in the field of metabolism of HCC with a focus on aerobic glycolysis,lipid metabolism,and glutamine addiction.Furthermore,we present some promising strategies for diagnosis and treatment based on tumor metabolism.This review facilitates to depict the metabolism landscape of HCC,thus providing therapeutic approaches for the prevention and treatment of HCC.
 World Journal of Gastrointestinal Oncology2021年9期
World Journal of Gastrointestinal Oncology2021年9期
- World Journal of Gastrointestinal Oncology的其它文章
- Use of liquid biopsies in gastrointestinal cancers
- Neoadjuvant chemotherapy without radiation as a potential alternative treatment for locally advanced rectal cancer:A metaanalysis
- Prognostic value of modified Lauren classification in gastric cancer
- Scoparone inhibits pancreatic cancer through PI3K/Akt signaling pathway
- Effect of oncometabolic surgery on gastric cancer:The remission of hypertension,type 2 diabetes mellitus,and beyond
- Immune aspects of hepatocellular carcinoma:From immune markers for early detection to immunotherapy