
大菱鲆源副乳房链球菌的分离鉴定及其毒力基因型
2021-09-17 10:31王鹤李战军黄华徐晓莹贺加贝曹亚男赵强
大连海洋大学学报 2021年4期
王鹤,李战军,黄华,徐晓莹,贺加贝,曹亚男,赵强*
(1.烟台市海洋经济研究院,山东 烟台 264006;2.山东省渔业发展和资源养护总站,山东 烟台 264003)
近年来,鲆鲽类养殖成功助推了中国海水养殖第四次产业化浪潮的形成,是北方海水养殖的重要产业[1],而大菱鲆Scophthalmusmaximus是鲆鲽类养殖最主要的种类之一。近年来,随着养殖规模的扩大及种质资源的退化,大菱鲆养殖病害频发,严重制约了海水养殖业的绿色健康发展。
链球菌病是一种全球范围流行的主要细菌性疾病,在以色列、意大利、美国、日本、韩国等国家的水产养殖中均有报道[2-5]。1958年首次报道了链球菌病在鱼类(日本养殖虹鳟Oncorhynchusmykiss)中暴发,之后在牙鲆Paralichthysolivaceus、条纹鲈Dicentrarchuslabrax、美国红鱼Sciaenopsocellatus、黄条鲱Seriolaquinquerodiata等众多养殖及野生鱼类中均有不同程度的感染[6-9],链球菌病已成为危害范围广、感染种类多的严重疾病[10]。鱼类链球菌病是可由不同种属病原菌引起的具有中枢神经损伤的类似病征,主要临床症状为眼球突出和脑膜脑炎[11]。据报道,可引起温水性(水温高于15 ℃)链球菌病的病原菌有:格氏乳球菌Lactococcusgarvieae、鱼乳球菌Lactococcuspiscium、鲑漫游球菌Vagococcussalmoninarum、海豚链球菌Streptococcusiniae、无乳链球菌Streptococcusagalactiac、副乳房链球菌Streptococcusparauberis等[12]。副乳房链球菌隶属于链球菌科Streptococcaceae链球菌属Streptococcus,是一种球状革兰氏阳性菌。最初研究者认为副乳房链球菌是引起牛乳房炎的主要病原之一,1993年西班牙养殖大菱鲆暴发链球菌病,研究者分离到该菌,并将其认定为鱼类病原菌[13]。目前,副乳房链球菌被认为是引起鱼类链球菌病的主要病原菌之一,尤其是引起日本和韩国养殖鱼类链球菌病的主要病原[5,7]。
多年来,中国还未见由副乳房链球菌引起鱼类链球菌感染的报道。2019年在对山东省烟台市大菱鲆主产区常规病害监测中,发现某区周边养殖场有以眼球突出、下颌红肿、腹部膨胀为主要症状的死亡病例。本研究中,以自然发病大菱鲆为试验材料,分离纯化获得1株优势菌,通过临床症状观察、生理生化鉴定、16S rRNA序列分析、特异性引物扩增等,鉴定其为副乳房链球菌。这也是中国副乳房链球菌对养殖大菱鲆自然感染致病的首次报道,本研究中通过对分离菌株开展毒力基因检测、血清学鉴定、抗生素敏感性测试等,探究了大菱鲆感染副乳房链球菌的发病原因,以期为该病的有效防治与致病机制研究提供科学参考。
1 材料与方法
1.1 材料
2019年7月,在山东省烟台市大菱鲆主产地的深井海水室内流水养殖车间,采集到患病大菱鲆样品。养殖水温为17 ℃,养殖密度约为20 kg/m2,患病个体体质量为195~535 g,平均体质量为408 g。健康大菱鲆购自烟台开发区天源水产有限公司(大菱鲆国家级良种场),平均体质量为351.1 g,平均体长为27.4 cm,养殖水温为(17±1)℃。
脑心浸液培养基(brian heart infusion,BHI)、水解酪蛋白琼脂培养基(muller hinton,MH)、革兰氏染色试剂盒均购自北京陆桥技术股份有限公司;生理生化鉴定管、药敏纸片购自杭州滨和微生物试剂有限公司;细菌基因组提取试剂盒、PCR扩增相关试剂购自宝生物(大连)有限公司;试验用引物及测序由生工生物工程(上海)股份有限公司完成。
1.2 方法
1.2.1 病原菌分离纯化 取有典型症状的濒死个体,用体积分数为75%的酒精擦拭腹部,解剖内脏,取其肝、肾、脾、脑、眼等组织,用无菌水冲洗、研磨后于BHI琼脂平板划线,28 ℃下倒置培养24~48 h,观察记录菌落形态特征,取优势菌纯化培养。经2~3次纯化后接种于体积分数为5%的羊血平板中观察溶血特性,同时用终体积分数为40%的甘油保种,于-80 ℃超低温冰箱中保存备用。
1.2.2 菌株鉴定 用革兰氏染色法对菌株染色观察,根据形态特征初步判断细菌种类。应用生理生化鉴定管测定菌株生理生化特性,进一步判断细菌种属。按照细菌基因组DNA提取试剂盒要求,用灭菌生理盐水将活化好的菌株调整至总密度约109CFU/mL,提取菌液DNA,使用16S rRNA通用引物(27F:5′AGAGTTTGATCMTGGCTCAG 3′,1492R: 5′GGTTACCTTGTTACGACTT 3′)扩增(表1),经测序后于NCBI网站进行Nucleotide Blast同源性分析,用MEGA 6.0 软件比对并构建系统发育树。选取基于23S rRNA、伴侣蛋白60(chaperonin 60, Cpn 60)和超氧化物歧化酶A(superoxide dismutase A, Sod A)基因片段设计的3对副乳房链球菌特异性引物进行扩增,PCR扩增产物片段大小与预期片段一致的,可进行测序验证。结合革兰氏染色、生理生化特性、16S rRNA系统进化关系及特异性引物扩增结果等综合判定分离菌株的种属。
1.2.3 人工回归感染试验 将分离株于BHI平板涂布培养24~48 h,用灭菌生理盐水冲洗纯培养物制成菌悬液,平板计数。将菌悬液配制成6.3×107、6.3×108、6.3×109、6.3×1010CFU/mL 4个浓度,通过腹腔注射进行人工感染。选取与患病个体体质量接近的健康大菱鲆作为回归感染对象,在实验室内暂养一周后,随机分为5组,每组10尾,4个试验组每尾鱼注射梯度菌悬液0.1 mL,对照组鱼注射等体积的无菌生理盐水。人工感染后养殖水温为(18.0±0.5)℃,每天换水并记录发病死亡情况,及时剖检死亡个体,再次取其肝、脾、肾等典型病变组织进行病原检测。
1.2.4 毒力基因检测 检测分离菌株的C5a肽基因(scpB)、αC蛋白基因(bca)、βC蛋白基因(bac)、层粘连蛋白结合蛋白基因(lmb)、纤维蛋白结合蛋白A基因(fbsA)、CAMP基因(cfb)、荚膜多糖基因(cpsD)、黏附和毒力蛋白A基因(pavA)、细菌免疫原性黏附素基因(bibA)、表面免疫相关蛋白基因(sip)、β溶血素基因(cylE)及透明质酸酶基因(hylB)等12个毒力相关基因的携带情况,其中,sip基因和bibA基因的PCR扩增条件分别参考彭民毅[16]和常瑞祥[17]的方法,其余毒力基因引物及扩增条件均参考Godoy等[18]的方法。用15 g/L琼脂糖电泳检测,紫外凝胶成像拍照后,将与预期目的片段大小一致的PCR扩增产物进行测序验证。
1.2.5 血清型检测 按照Tu等[19]基于多糖聚合酶(WZY)基因设计的副乳房链球菌不同血清型引物(表2),对本研究中的分离株开展血清型鉴定。

表2 不同血清型的引物序列
1.2.6 药敏试验 按照美国临床检验标准委员会(CLSI)推荐的纸片扩散法(Kirby-Bauer, K-B)操作标准进行抗生素药物敏感性测试。用麦氏比浊法将分离菌株配制成浓度为1.0×108CFU/mL的菌悬液,均匀涂布于MH平板,贴药敏纸片,于28 ℃下培养24 h后用游标卡尺测量抑菌圈大小。测试用抗生素共计10类23种,每种药物设置4个重复。依据CLSI标准判定药物敏感性,标准未涵盖部分参照王巧煌[20]的判定标准。
2 结果与分析
2.1 临床症状与解剖观察
患病大菱鲆表现为食欲减退,体色发黑,游泳状态失衡、打转,具眼球突出、下颌红肿及腹部膨大等特征,个别鳍条基部出血、体表溃疡(图1)。剖检发现,肝呈花斑状充血,脾肿大、颜色暗黑,腹腔腹水。取样车间患该症状大菱鲆约占车间养殖总量的3%,10 d内患病个体死亡率约为75%。
2.2 表型特征及生理生化反应
试验中经病原菌分离纯化,获得1株优势菌,命名为BH2-4。该菌株在BHI平板上形成圆形、半透明、乳白色、微隆起菌落,菌落表面光滑湿润,边缘整齐,直径为0.5~0.8 mm。革兰氏染色后显微镜检为紫色、球形、链状细菌,在体积分数5%的羊血平板中不溶血(图2)。
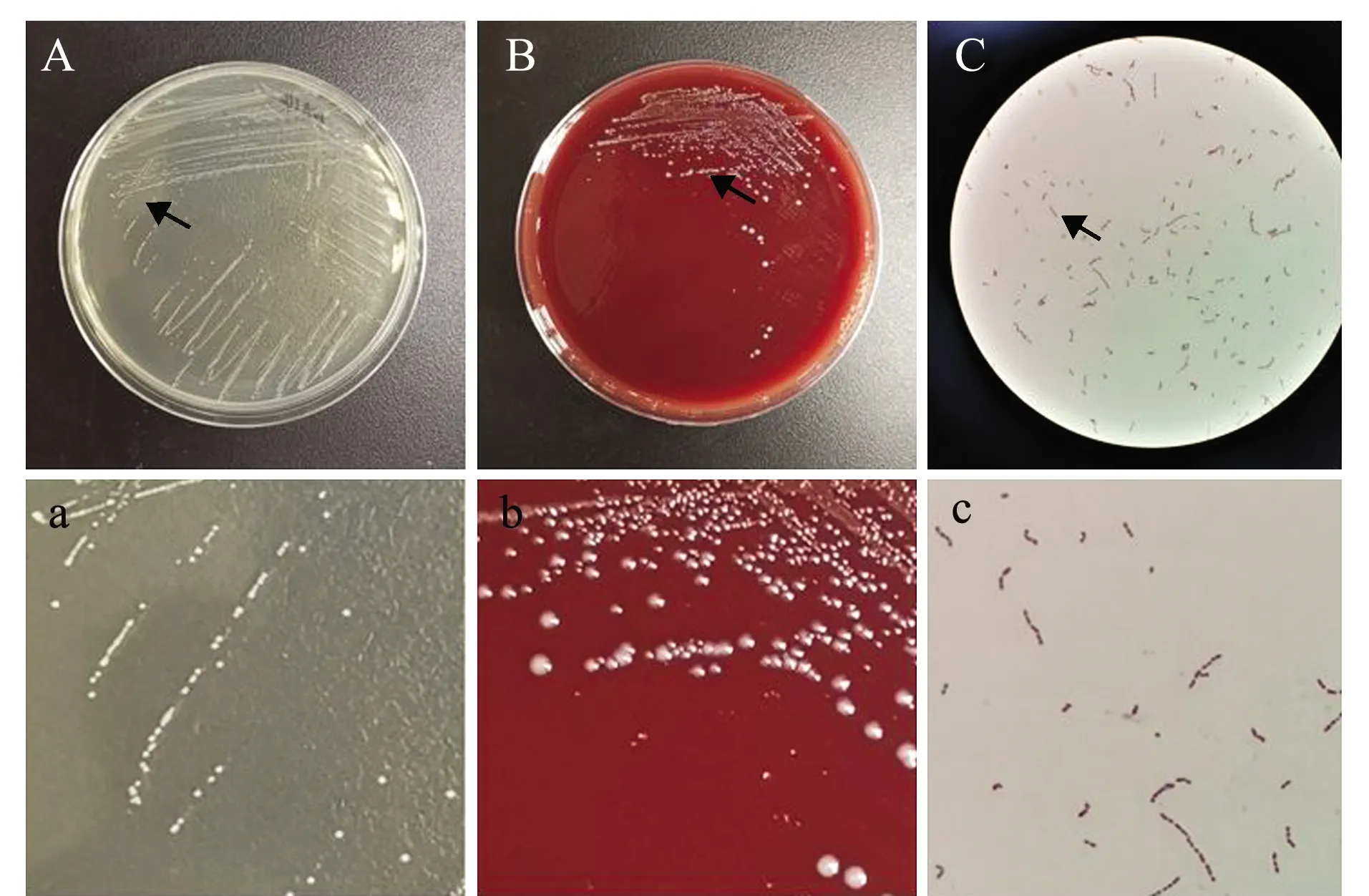
A—BHI培养基菌落形态;B—5%羊血培养基中菌落形态;C—革兰氏染色显微观察;a—图A箭头处放大;b—图B箭头处放大;c—图C箭头处放大。
生理生化反应结果显示:在10 ℃、45 ℃、pH 9.6及胆汁中不能生长,可在质量分数为6.5%的NaCl中生长;能水解七叶苷,不能水解精氨酸、赖氨酸;能利用蔗糖、甘露醇、葡萄糖、山梨糖、蕈糖产酸,不能利用枸橼酸、鼠李糖、蜜二糖、肌醇、阿拉伯糖、木糖;具碱性磷酸酶活性,不具β-半乳糖苷酶活性,不产生吲哚反应等(表3)。
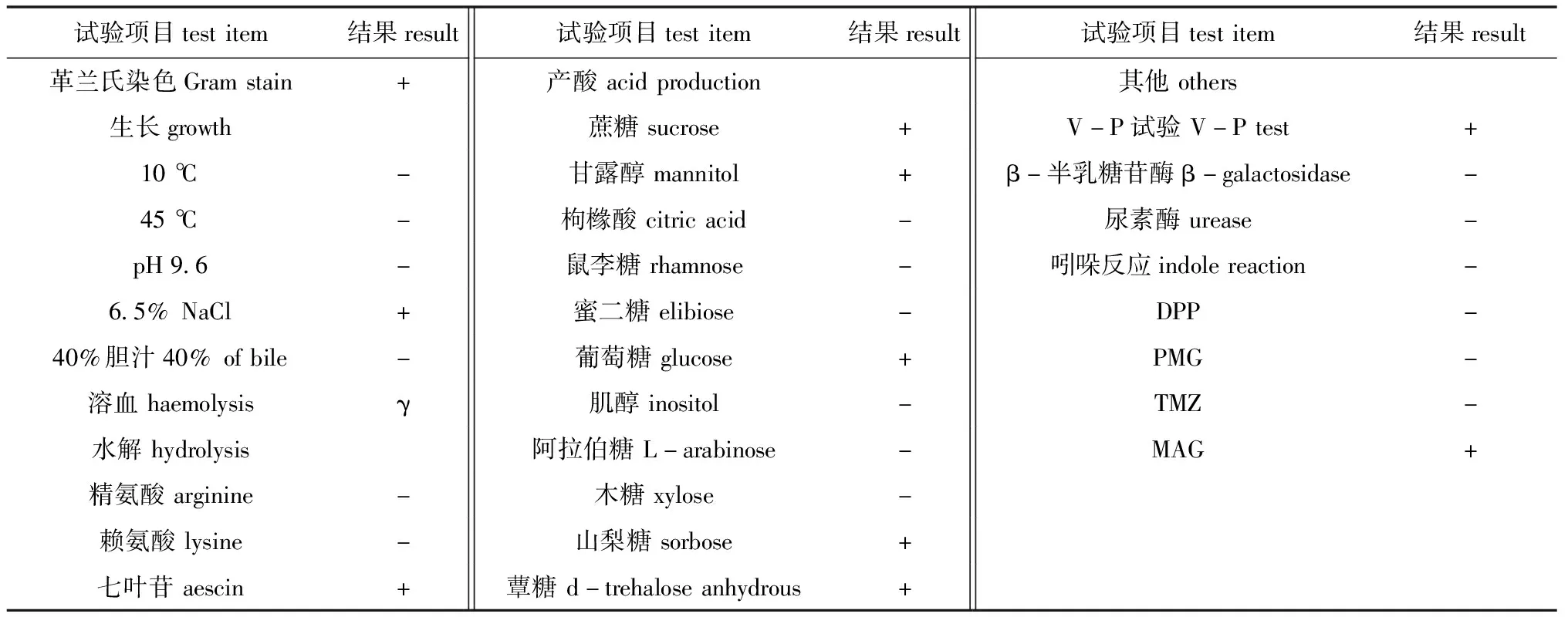
表3 菌株BH2-4的生理生化特性
2.3 分子特征鉴定
应用通用引物扩增BH2-4分离株的16S rRNA基因,获得约1 500 bp的片段,PCR产物纯化测序及NCBI数据库在线比对,结果显示,菌株BH2-4的16S rRNA与S.parauberisCIFT MFB10119、F21、RP25、DSM631、LMG14376株同源性最高(GenBank登录号分别为KP240952.1、KP137361.1、JQ988835.1、AY58447.1、AY942573.1),一致性均为100%。基于副乳房链球菌、乳房链球菌S.beris、海豚链球菌、无乳链球菌、停乳链球菌S.dysgactiae等菌株16S rRNA序列构建系统发育树(图3),结果显示,分离菌株与副乳房链球菌5条序列聚为一支后再与乳房链球菌聚为一支。

图3 基于16S rRNA序列构建的菌株BH2-4系统发育树
分离株BH2-4的23S rRNA、cpn60、SodA基因序列扩增,分别得到片段大小为718、400、254 bp的产物,琼脂糖凝胶电泳检测结果见图4。
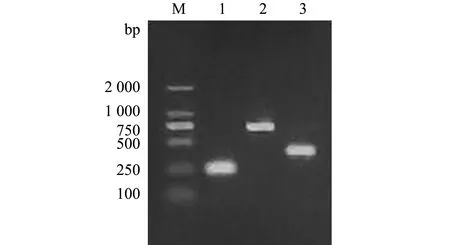
M—DNA分子标准;1—Sod A基因特异性引物扩增;2—23S rRNA基因特异性引物扩增;3—cpn 60基因特异性引物扩增。
2.4 人工回感试验
用不同浓度的菌液人工感染健康大菱鲆后,各组试验鱼伏于池底,对外界干扰反应迟缓,受强烈刺激后反应剧烈,出现无方向性窜游甚至撞壁现象。注射48 h后部分大菱鲆出现鳍条充血、眼球突出、下颌红肿、腹部膨大、内脏出血、脾肿大等症状,与自然感染发病鱼临床症状及剖解病变相似,各菌浓度组感染后症状详见表4和图5。

表4 不同浓度菌株人工感染健康大菱鲆后的临床特征
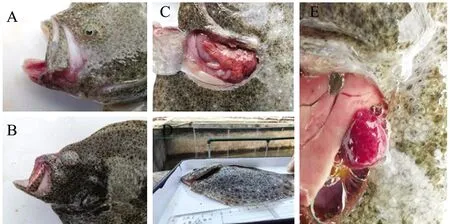
A—下颌红肿充血;B—眼球突出;C—肝脏出血;D—腹胀;E—脾脏肿大有白点。
从图6可见:6.3×1010CFU/mL最高菌浓度组在第6天开始出现死亡,死亡高峰出现在第10~11天,第13天时全部死亡;6.3×109CFU/mL菌浓度组第9天开始出现死亡,第15天时累计死亡率为90%。根据各浓度组死亡情况,采用改良寇氏法计算得到菌株BH2-4对受试鱼的LD50为6.9×108CFU/mL。从人工感染发病死亡鱼体的脾脏、肾脏等组织再次分离到回归感染所用菌株。
2.5 分离菌株的毒力基因型
分离菌株BH2-4的毒力基因型为scpB--bca--bac--lmb--fbsA--cfb--cpsD--pavA--bibA+-sip+-cylE--hylB+。本研究中选取了12个与链球菌相关的毒力基因,仅检测出sip、hylB和bibA3种,其余基因检测均为阴性(图7)。
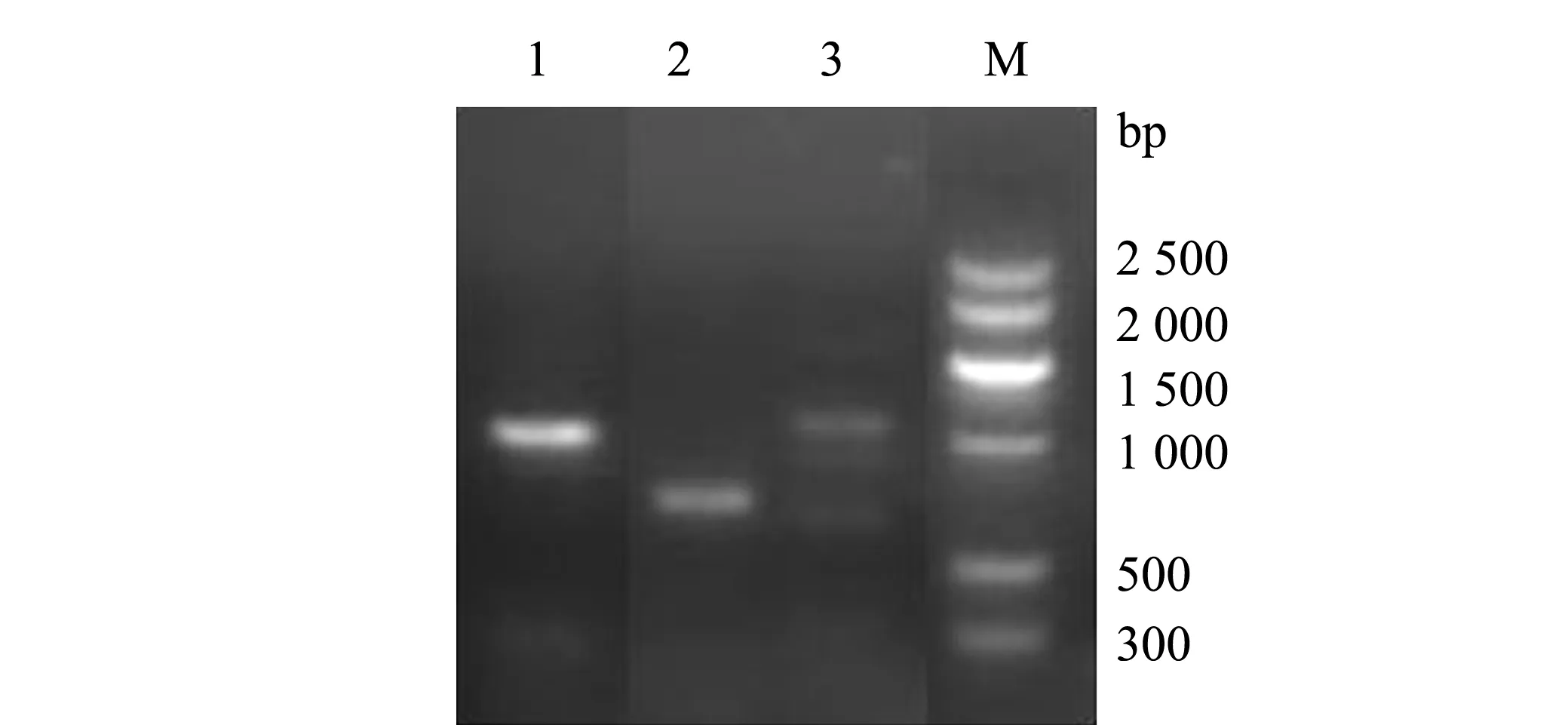
M—DNA分子量标准;1—sip基因;2—hyl B基因;3—bib A基因。
2.6 血清学检测结果
根据日本学者的研究结果,副乳房链球菌血清型可分为Ⅰ型、Ⅱ型和未分型(non-typed, NT)3种,其中,血清型Ⅰ又可分为Ⅰa、Ⅰb、Ⅰc 3种亚型[21]。应用WZY基因引物进行特异性扩增,可有效鉴别Ⅰa、Ⅰb、Ⅰc和Ⅱ型[19]。本研究中分离株BH2-4应用上述引物进行血清型扩增并未得到明显PCR产物,无法鉴别其血清型。
2.7 药敏试验结果
从表5可见:菌株BH2-4对10类23种测试抗生素的敏感性存在差异,对β-内酰胺类5种、喹诺酮类4种、四环素类1种、大环内酯类1种、多肽类1种、利福霉素类1种、氯霉素类1种在内的7类14种抗生素类药物敏感;对包括氨基糖苷类3种、磺胺类2种及呋喃类1种在内的3类6种抗生素耐药;对诺氟沙星、强力霉素、多粘菌素B中度敏感。

表5 菌株BH2-4对10类23种抗生素的敏感性(n=4)
3 讨论
3.1 副乳房链球菌的鉴定与流行病学
副乳房链球菌最初被认为是乳房链球菌Ⅱ型,具有与乳房链球菌相似的生理生化特性,直到1990年Williams等[22]通过反转录酶序列分析,发现二者在系统发育上截然不同,由此提出新物种——副乳房链球菌。本研究中的生理生化鉴定结果与Williams等[22]对副乳房链球菌新物种中的描述一致。尽管本研究中分离株BH2-4表现为γ溶血,与近年来报道的副乳房链球菌溶血特性不一致,但与物种鉴定描述中的“γ溶血或微弱的α溶血”吻合。Nho等[7]研究牙鲆源副乳房链球菌时发现,从有典型链球菌症状患病牙鲆中分离到的菌株具α溶血特性,但回归感染后再次从濒死或死亡牙鲆分离到的菌株则表现为不溶血。有研究表明,海豚源链球菌的溶血特性与血平板中添加的血液类型及基础培养基有关[23]。海豚源链球菌在添加牛血或人血的琼脂平板中呈α溶血或部分α溶血,而在添加羊血的平板中则呈β溶血;在体积分数为3%的马血心浸液血平板中呈β溶血,而在添加同种同等比例血液的Todd-Hewitt平板中呈α溶血[24]。
鱼类链球菌感染泛指以败血症、脑膜炎等为主要症状的相似但又不尽相同的病症,据统计至少6种不同的病原菌均可诱发链球菌感染[14]。链球菌种类的多样性较好地解释了仅通过表型鉴定链球菌病的难点所在,受生长速度、接种水平和潜伏期等因素影响,基于生理生化、抗原特性的病原菌鉴别较难精确鉴定到种属水平[11,25]。在分子鉴定方法中,候选基因测序非常适合细菌的鉴定,分子鉴定是优于传统理化方法的最佳选择,并广泛应用于链球菌中[11-12,14]。本研究中应用Mata等[14]、Alber等[15]基于23S rRNA、伴侣蛋白60(cpn60)和超氧化物歧化酶A(SodA)基因片段设计的副乳房链球菌特异性引物扩增得到了预期目的片段大小的条带,与他人应用该片段扩增的结果一致[7,26-27]。尤其23S rRNA基因退火温度从50 ℃[26]至60 ℃[27]均能得到稳定的扩增结果,是鉴定副乳房链球菌特异性强且稳定性好的理想基因。
副乳房链球菌致病范围较广,曾有牛、猪、鱼等感染致病的报道[28-30]。以前中国未见鱼源副乳房链球菌致病报道,但日本和韩国养殖的牙鲆链球菌感染多由该菌引起,曾造成了严重的经济损失[19,21]。副乳房链球菌感染分急性感染和慢性感染两种,急性感染3~7 d内死亡率超过50%,慢性感染数周内只有少量死亡[30]。鱼类感染该链球菌后,一般表现为眼球突出、腹水腹胀、脾脏肾脏肿胀或苍白、内源性或外源性出血等症状[13,31-32]。本研究中,BH2-4株回归感染亦出现类似病症,且与分离到该菌株的自然发病症状相似。研究表明,副乳房链球菌人工感染的死亡率与注射部位、水温密切相关,Mori等[33]试验结果表明,背鳍基部注射死亡率高于腹腔注射;Kim等[31]报道,23~24 ℃水温下副乳房链球菌感染牙鲆的LD50为1×102~1×103CFU/mL,Han等[34]报道,21 ℃时不同血清型副乳房链球菌感染的半致死浓度分别为107、108、109CFU/mL,水温在26 ℃时的感染死亡率显著高于22 ℃[32]。本研究中,人工感染水温为(18.0±0.5)℃,腹腔注射菌株BH2-4对大菱鲆的LD50为6.9×108CFU/mL,与Han等[34]的研究结果相近。此外,土耳其、北美洲亦有从健康鱼体中分离到副乳房链球菌的报道,但用从健康条纹鲈鱼中分离到的副乳房链球菌人工感染却并未导致发病,由此分析,分离得到病原菌的机体可能是致病菌携带者或曾患病痊愈者,也可能条纹鲈不是副乳房链球菌的敏感宿主[11,26]。
3.2 副乳房链球菌毒力基因与血清型
透明质酸酶(hyaluronidase)属碱性糖蛋白类蛋白水解酶,可以特异性地分解细胞外基质透明质酸(hyaluronidase acid),为细菌生长繁殖提供必需的碳源和能量[35],是细菌致病的重要毒力因子,尤其在链球菌等革兰氏阳性菌的致病机制中发挥重要作用[36]。研究表明,透明质酸酶B(hyl B)是无乳链球菌重要的毒力因子之一,且不同来源的无乳链球菌hylB基因具有高度的保守性,通过对其蛋白表达及抗原性分析可为疫苗的研制提供良好基础[37]。表面免疫原性蛋白(surface immunogenic protein,Sip),是暴露于菌体表面的一类免疫相关蛋白,与细菌对宿主的黏附、定植有关[38],因其具有高度的保守性,广泛分布于人源和牛源的多个血清型无乳链球菌菌株中,是研究疫苗的良好候选靶抗原[39]。B族链球菌细胞表面可产生免疫原性黏附蛋白(GBS immunogenic bacterial adhesion, Bib A),能促进B族链球菌吸附到宿主细胞表面[17],与C4结合蛋白特异性结合,对其抵抗机体吞噬及毒理作用发挥起到关键作用,缺失bibA基因的菌株存活及抗吞噬作用明显降低[40]。本研究中测定了分离株BH2-4对链球菌12种常见毒力基因的携带情况,结果只检测出上述3种基因(sip、hylB、bibA),其余孔毒素(cfb、cylE)、促进免疫逃避因子(cpsD、scpB)或黏附因子(fbsA、lmb、bca、bac、pavA)均未检出。有研究表明,链球菌的致病力取决于细菌在宿主免疫细胞中的存活及避免被宿主细胞杀死而建立感染、诱导细胞凋亡的能力[41-42]。海豚链球菌的致病力完全取决于其毒力因子,对其菌株致病力的判断可以通过检测毒力因子加以辨别[43],而副乳房链球菌的相关研究报道较少,其致病机理与毒力基因关联度有待进一步研究。
遗传变异源于碱基的突变、插入或缺失,这些变异可能会影响进化趋势,或者说菌株的血清型与毒力间可能存在联系[42]。日本学者[21,44]将本土牙鲆源副乳房链球菌血清型进行了详细分类,可分为3大类5种不同类型。Tu等[19]基于多糖聚合酶基因(WZY)设计的引物可有效区分血清型Ⅰ型(Ⅰa、Ⅰb、Ⅰc)和Ⅱ型。应用该引物扩增本研究中分离株并不能得到明显的PCR产物,表明BH2-4株与日本牙鲆源副乳房链球菌的WZY基因可能存在差异。韩国学者在兔抗血清凝集试验中发现本土牙鲆源副乳房链球菌血清型亦可分为Ⅰ、Ⅱ两型,Ⅰ型毒力强于Ⅱ型,其毒力差异可能与细胞表面包膜厚度不同有关[44]。在对血清学相关研究中发现,副乳房链球菌荚膜机体抗血清杀伤活性和抗巨噬细胞吞噬能力有关[45]。荚膜多糖参与血清学反应,荚膜多糖结构的变异可能是导致血清学差异的原因之一[46],未分型副乳房链球菌未能成功分型的原因可能与其荚膜结构缺失有关。
3.3 药物敏感性
目前,对链球菌的防治主要以抗菌药物为主,药物的敏感性试验对抗菌药物的选择具有重要的临床指导意义。本研究中,分离得到的菌株BH2-4菌除对链霉素、庆大霉素、卡那霉素3种氨基糖苷类药物,磺胺异恶唑、复方新诺明2种磺胺类药物及呋喃类的呋喃唑酮耐药外,对β-内酰胺类、喹诺酮类、四环素类、大环内酯类、多肽类、利福霉素类的18种测试药物均敏感(其中包括4种中度敏感),在耐药基因携带方面,检测到BH2-4株携带磺胺类sul1和喹诺酮类gyrA两种耐药基因(检测结果未列出)。Lee等[47]研究韩国牙鲆源副乳房链球菌时发现,其分离株对氨苄西林、阿莫西林、多西环素、土霉素、庆大霉素、红霉素、复方新诺明等药物存在多重耐药现象;土耳其虹鳟源副乳房链球菌分离株对万古霉素、红霉素敏感,对依诺沙星、土霉素和氯霉素耐药[11];野生条纹鲈分离株对复方新诺明耐药,对磺胺类药物普遍中度敏感[26];86株韩国牙鲆源副乳房链球菌中,63%的菌株对四环素耐药,58%的菌株对红霉素耐药,16%的菌株对青霉素耐药[30];370株韩国黄尾鰤源γ溶血链球菌中,17%的菌株对大环内酯、利霉素、四环素、氯霉素、红霉素耐药[4]。本研究结果与上述研究结论相符,分离的菌株BH2-4对药物比较敏感,并未出现严重的多重耐药。尽管应用β-内酰胺类、喹诺酮类、大环内酯类、利福霉素类药物均能起到较好的治疗效果,但其中的头孢曲松、头孢拉啶、头孢哌酮、诺氟沙星、氧氟沙星、洛美沙星、红霉素等敏感药物已在水产中明令禁止使用;在其他药物的选择及使用中应遵循水产用药相关规定或参照最新版兽药典。
4 结论
1)本研究中从以突眼、腹水、红肿为主要症状的患病大菱鲆中获得的优势菌株BH2-4为γ溶血的革兰氏阳性菌,具有与副乳房链球菌一致的生理生化特性。
2)经16S rRNA通用引物扩增、Blast在线比对,链球菌特异性引物扩增等,综合生理生化结果,鉴定优势株BH2-4为副乳房链球菌Streptococcusparauberis。
3)BH2-4的毒力基因型为scpB--bca--bac--lmb--fbsA--cfb--cpsD--pavA--bibA+-sip+-cylE--hylB+。应用WZY基因引物无法有效鉴别BH2-4的血清型。
4)腹腔注射健康大菱鲆,半致死浓度LD50为6.9×108CFU/mL。
5)根据药物敏感性测试结果,在副乳房链球菌引起的大菱鲆链球菌病防治中,推荐使用恩诺沙星粉(水产用),并执行500度日的休药期。
猜你喜欢
黑龙江水产(2022年6期)2022-12-03
农药科学与管理(2019年6期)2019-11-23
中国水产(2019年3期)2019-03-25
海洋科学(2017年10期)2017-02-09
华南农业大学学报(2015年5期)2015-12-04
华南农业大学学报(2015年5期)2015-12-04
中国兽医杂志(2015年8期)2015-08-08
医学综述(2014年14期)2014-03-27
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28
现代检验医学杂志(2014年5期)2014-02-02
