
小麦幼苗根系生理特性和解剖结构对根际pH的响应
2021-09-16 09:41王光涛冯素伟丁位华金立桥文昭普茹振钢
西北农业学报 2021年8期
王光涛,冯素伟,丁位华,金立桥,文昭普,茹振钢
(1.河南科技学院 小麦研究中心,河南新乡 453003;2.现代生物育种河南省协同创新中心,河南新乡 453003)
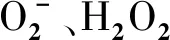
前人研究发现,紫穗槐[11]、水稻[12]、大豆[13]和小麦[14]在不同的胁迫处理下抗氧化酶活性及根系细胞结构表现不同的反应,但这些研究大多单一地集中在干旱胁迫、碱胁迫、铝胁迫及盐胁迫上。然而,关于不同pH 条件下的小麦抗氧化酶活性及根系微观结构的研究报道较少。为此,通过水培法模拟不同根际酸碱环境,研究不同pH 对小麦幼苗期生物量、根冠比、根系抗氧化酶活性、丙二醛(MDA)含量和根系解剖结构的影响,探讨小麦适应不同根际pH 的根系调控机制,以期为筛选耐酸碱品种,提升小麦产量提供理论 依据。
1 材料与方法
1.1 试验材料与设计
本试验选用黄淮麦区大面积栽培的两个半冬性小麦品种‘矮抗58’(AK58)和‘百农4199’(BN4199),均由河南科技学院小麦中心提供。
小麦种子经φ=1% H2O2表面消毒24 h,再用蒸馏水冲洗干净。将预先浸种的种子放入培养皿中。发芽3 d后,将大小均匀的小麦幼苗移栽到Hoagland营养液中进行水培,培养3 d后,营养液设置3个pH 水平:pH 4.0(酸胁迫)、pH 6.5(正常环境)和pH 9.0(碱胁迫),溶液pH 分别用1 mol/L HCl和1 mol/L NaOH调节。植物培养在光周期14 h(光照度20 000 lx)、昼夜温度22 ℃/19 ℃、相对湿度64%的光照培养箱内进行。每天随机移动培养箱以减少位置影响,定时更换营养液并调节至对应pH ,培养16 d。幼苗数量保证每个指标测10~15株,完全随机设计。取样小麦植株为三叶期幼苗。
1.2 测定项目及方法
1.2.1 干物质测定 蒸馏水冲洗根部,吸水纸吸干表面水分。将地上部和根部分开,105 ℃杀青30 min,于70 ℃烘至恒量,称干质量。
1.2.2 根系抗氧化酶及MDA含量测定 参照郝再彬等[15]的方法测定抗氧化酶活性及丙二醛含量。超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)法,过氧化物酶(POD)活性采用愈创木酚显色法,过氧化氢酶(CAT)活性采用紫外分光光度法,丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法。
1.2.3 根系解剖结构 参照李和平[16]的方法制作石蜡切片观察小麦幼苗根系的解剖结构。先用去离子水冲洗幼苗根部,然后切取根尖0~1 cm,迅速固定于FAA(90 mL 50%醇+5 mL甲醛+ 5 mL冰乙酸)固定液中,抽气,24 h后梯度酒精脱水,制作石蜡切片,对根尖材料纵切,苏木精染色,OlympusCX41镜检照相。
1.3 数据处理
采用Microsoft Excel 2016进行数据处理和作图,用SPSS 19.0进行数据统计分析。
2 结果与分析
2.1 不同pH对植株生物量积累的影响
如图1所示,不同pH 条件下两个品种间根系干质量差异显著,而地上部干质量差异不显著。两个品种地上部及根系生物量积累均表现为pH 6.5>pH 9.0>pH 4.0,说明酸碱胁迫会抑制生物量的积累,酸胁迫对植物的伤害大于碱胁迫。BN4199在不同pH 条件下根系干质量显著高于AK58,地上部干质量高于AK58但差异不显著。两个品种根冠比均表现为pH 4.0 由图2所示,不同根际pH 对小麦根系抗氧化酶活性有显著影响。不同根际pH 下2个品种的根系SOD、POD和CAT活性表现为pH 4.0 如图3所示,不同pH 条件下,根系MDA含量有较大的差异,2个品种均表现为pH 6.5 如图4所示,2个品种的根系解剖结构在不同pH 条件下有显著变化。pH 6.5条件下2个品种根系结构无明显差异,根系细胞排列整齐、紧密,表皮细胞完好。pH 9.0条件下,2个品种根系结构较对照受损较小,根系细胞排列较为整齐,细胞形态规则,表皮及根冠区细胞较为完整。pH 4.0条件下较对照根系受损较大,表皮细胞破损,细胞排列疏松紊乱、细胞形态不规则,根冠区细胞受损严重。2个品种在pH 9.0条件下细胞形态及排列、表皮和根冠区细胞好于pH 4.0条件,说明酸性条件对根系结构的损害程度大于碱性条件。‘BN4199’根系结构在pH4.0条件和pH9.0条件下根系细胞的排列及形状、根冠区细胞的完整性好于‘AK58’。 如表1所示,地上部和根系干物质积累量与SOD、POD、CAT活性呈显著正相关,与根系MDA含量呈极显著负相关,说明保持较高的根系抗氧化酶活性对保护根系膜结构和植物生物量的积累有着重要的作用。 表1 不同测定指标的相关性Table 1 Relevance of different parameters 根际pH 的变化会改变土壤中的养分有效性,还可以影响微生物的活性以及矿物养分的吸收[17],从而影响植物的生长发育。徐呈祥[18]研究认为生物量的积累准确地反映植物对胁迫的响应。本试验发现,酸碱胁迫下不利于小麦植株干物质积累,说明酸碱胁迫抑制养分的吸收,使生物量积累降低,而碱胁迫较酸胁迫有更强的营养吸收能力。根冠比体现同化物在地上和地下器官之间的分配,根系结构对环境变化的响应表现很大的可塑性,并且对作物在胁迫条件下的发育至关重要[19]。本试验表明碱胁迫下植株干物质积累及根冠比大于酸胁迫,李菊艳等[20]研究认为提高根冠比可以增强耐盐性,可见酸碱条件下根冠比的提高有利于提高抗逆能力及干物质积累。 当ROS不完全还原时会氧化生物分子,导致细胞氧化破坏,积累许多有害的过氧化产物,如MDA等[21]。植物体内的某些酶和抗氧化剂可以降低和抑制活性氧对植物细胞的伤害[22-23]。本试验发现,酸碱胁迫抑制小麦根系抗氧化酶活性,提高根系膜脂过氧化程度,说明酸碱胁迫抑制根系的生理活动。根系解剖结构是根系发育水平的体现,与根系生理密切相关,对逆境供水和保证植物活力具有重要作用[24]。Choudury等[25]研究认为在各种胁迫下过量的ROS积累是细胞损伤的主要原因。本试验发现碱性条件下生物量积累、根冠比、抗氧化酶活性均高于酸性条件,而酸性条件对根系结构的损害程度大于碱性条件。说明酸碱胁迫下保持较高的抗氧化酶活性有利于根系生长,提高根冠比可以适应不良的根际酸碱环境。相关分析表明保持较高的根系抗氧化酶活性对保护根系膜结构和植株生物量的积累有重要作用。酸碱胁迫下‘BN4199’地上部和根系生物量积累、根系抗氧化酶活性高于‘AK58’,根系结构好于‘AK58’,而根系MDA含量小于‘AK58’,说明抗酸碱胁迫强的品种,膜脂过氧化程度较低,根系结构受损较小。Terletskaya等[26]研究认为耐性较强的小麦品种的根冠比随胁迫而增加,而敏感品种的根冠比则下降。‘BN4199’在酸碱条件下根冠比显著高于‘AK58’,说明‘BN4199’具有较强的耐酸碱能力,根冠比可作为小麦苗期筛选耐酸、碱品种的参考指标。 综上表明,不同小麦品种耐酸碱能力存在较大差异,酸胁迫对小麦生长的抑制程度大于碱胁迫。酸碱条件下,保持较高的抗氧化酶活性有利于稳定根系生长,以适应不良根际环境。与‘AK58’相比,‘BN4199’具有较强的耐酸碱能力。2.2 不同pH对小麦幼苗根系抗氧化酶活性的影响
2.3 不同pH对小麦幼苗根系MDA含量的影响
2.4 不同pH对小麦幼苗根系解剖结构的影响
2.5 地上部和根系各指标之间的相关性分析

3 讨 论
4 结 论
猜你喜欢
中国交通信息化(2022年7期)2022-10-27农业工程学报(2022年10期)2022-08-22中国土壤与肥料(2022年4期)2022-06-14林业调查规划(2022年3期)2022-06-13福建轻纺(2022年4期)2022-06-01林业科学(2022年2期)2022-05-11四川农业科技(2021年7期)2021-12-01中国生态农业学报(中英文)(2021年9期)2021-09-10中老年保健(2021年7期)2021-08-22安徽农学通报(2020年7期)2020-05-26
