
缺氮胁迫阶段及施氮时期对油用亚麻干物质及氮素积累的影响
2021-09-16 04:38:14郭丽琢高玉红牛俊义
干旱地区农业研究 2021年5期
郭丽琢,杨 波,高玉红,牛俊义
(1.甘肃农业大学农学院,甘肃 兰州 730070;2.甘肃省干旱生境作物学重点实验室,甘肃 兰州 730070;3.甘肃省陇南市农业技术推广总站,甘肃 武都 746000)
油用亚麻是世界性的特色油料作物[1]。油脂中高达45%~65%的α-亚麻酸含量[2],以及丰富的植物资源和低廉的价格,使其成为食疗获取α-亚麻酸的首选优质资源,亚麻油也在食用油消费结构升级的大潮中成为新宠;较强的干燥性又使亚麻油广泛应用于工业领域。籽粒富含的木酚素、多酚、植物甾醇等特色功能性生理活性物质,具有国际医学界和营养学界所公认的诸多显著生理功效[3]。功能性成分基础研究和高值化研究的深入,以及加工技术的日臻完善,突显了亚麻籽粒食用及化工生产之外的医用、保健、美容开发潜力,培育了新的经济增长点。广泛的用途及较高的经济附加值,扩大了亚麻籽粒的市场需求量。我国虽为世界第四大亚麻籽生产国[4],但近年来国内原料缺口逐年增大,对外依存度不断提高,目前已成为第二大亚麻籽进口国[4]。遵循“内外兼修”的发展思路[4],提高国内油用亚麻的产量、质量成为必需,以在助力地区经济发展的同时满足中国居民对高品质、多功能亚麻籽加工产品的消费需求。
我国油用亚麻的单产仅略高于世界平均水平[4],单产较低的原因之一是生产上的肥料运筹未得到优化。明晰主产区主推品种对养分供缺的响应,是制定合理施肥制度的前提和关键。相对而言,油料作物需要充足的氮素营养[5]。油用亚麻是需氮较多而又不耐高氮的作物,单位籽粒产量形成吸收的氮素较禾本科作物高30%~57%[6],适量适时供氮对保证其高产优质至关重要。亚麻氮素营养的研究,国内多集中于适宜的氮肥用量、氮磷钾配施以及水氮耦合等[7-8],揭示主推品种氮素营养特性及需氮规律的报道较少,关于其氮素缺乏及缺素阶段的影响,以及生长对施氮时期的响应更是鲜见报道。
国内少量的亚麻缺氮模拟研究均局限于苗期缺素[9-10]。现有的探索均表明,需氮较少的苗期[8]短期缺氮后,亚麻的生长发育及干物质积累即受到了显著影响,但即使同为营养生长阶段的缺氮胁迫,其生长发育的响应可截然不同。播种后至8叶间的缺氮胁迫下,干物质积累较完全营养液显著增加[9];而苗高8~10 cm时开始持续缺氮,3~20 d的生长均受抑、干物质量显著降低[10]。可见,缺氮胁迫的影响,因胁迫时期和历时而具有较大差异。作物的需氮规律与生育进程密切相关,油用亚麻苗期的氮素积累量仅占整个生育期总量的10%~20%[8]。需氮较少的营养生长阶段前期,缺氮的影响尚因缺素时期而差异较大;那需氮量较多的中后期缺氮,胁迫时期和历时的影响差异可能会更大。植物受到伤害后会产生一种积极的自我调节能力,即补偿效应[11]。而亚麻阶段性缺氮后,复氮的补偿效应鲜见报道,胁迫时期和补偿生长类型间的关系尚未明晰。系统探索缺氮时期及胁迫时长对亚麻生长的影响,明晰缺氮阶段影响的敏感程度及其补偿效应差异,据此确定氮肥施用的高效期和迟钝期,才能切实合理运筹氮素资源,促进亚麻高产稳产,助力农业节肥增效。
提高肥料利用率的关键是同步作物的养分需求与土壤、肥料的养分供给,即合理的基肥和追肥用量及比例。作物前期生长需氮少,加之1978—2008年间我国氮肥施用量3.58倍的增加导致的土壤供氮能力提高[12],降低基肥氮、提高追肥比例并推迟追肥时间的“氮肥后移”、全生育期多次追施成为许多作物进一步提高产量的途径[13-17]。油用亚麻生产中的氮肥以基肥为主,即使追肥,通常也是一次,时间多于现蕾或其之前,基肥∶追肥配比的随意性较高,缺乏根据营养规律和土壤环境特性的三宜施肥技术。根据亚麻缺氮阶段影响的敏感程度,设定基肥、追肥的用量及比例,进一步探索氮肥施用时期与干物质及氮素积累的关系,可为制定亚麻适宜的氮肥运筹方案提供理论依据。
本研究通过砂培和土培模拟试验,探讨了缺氮胁迫阶段和施氮时期对油用亚麻干物质及氮素积累的影响,以期揭示缺氮时期影响的敏感程度和复氮的补偿效应,探索氮肥施用时期的运筹效果,为优化增产节肥的氮素管理奠定理论基础。
1 材料与方法
1.1 试验材料
油用亚麻:陇亚杂1号品种。
供试土壤:采自兰州市安宁区,选用速效氮含量较低的土壤,有机质12.85 g·kg-1,全氮1.18 g·kg-1,碱解氮19.6 mg·kg-1,速效磷8.54 mg·kg-1,速效钾132 mg·kg-1。
1.2 试验设计
试验于2018年和2019年的3—8月进行。
试验一:阶段性缺氮胁迫试验,通过砂培模拟试验进行。采用单因素试验设计,以缺氮阶段为试验因素,各处理中除缺氮阶段外,其余阶段均正常供氮。试验以全生育期供氮为对照(CK),另设5个缺氮阶段水平,共6个处理(表1)。分别于枞形、现蕾、盛花和成熟期采样,每个处理均种植24盆,重复6次,共计144盆。
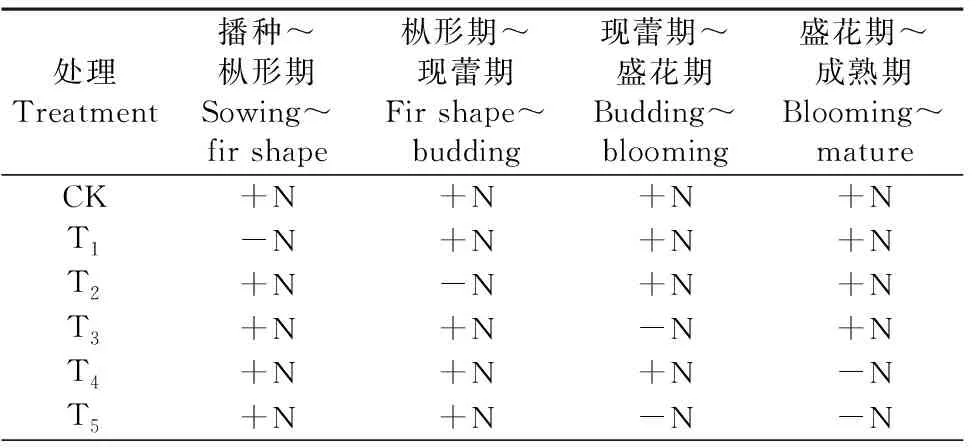
表1 氮素阶段性缺乏的处理方案Table 1 Nitrogen deficiency stage treatment scheme
模拟培养的营养液(表2),大量元素配方以Hoagland营养液(一)[18]及缺氮的不完全Hoagland营养液[18]为基准;除铁之外的其他微量元素,以Arnon微量元素混合液[18]为配方提供,1L营养液中加入1mL Arnon微量元素混合液;为保证铁盐的溶解性,用EDTA-Fe单独配制铁盐母液,母液Fe的浓度为0.018 mol·L-1,营养液中铁的浓度以Hoagland营养液(一)配方中的铁浓度为基准[18]。
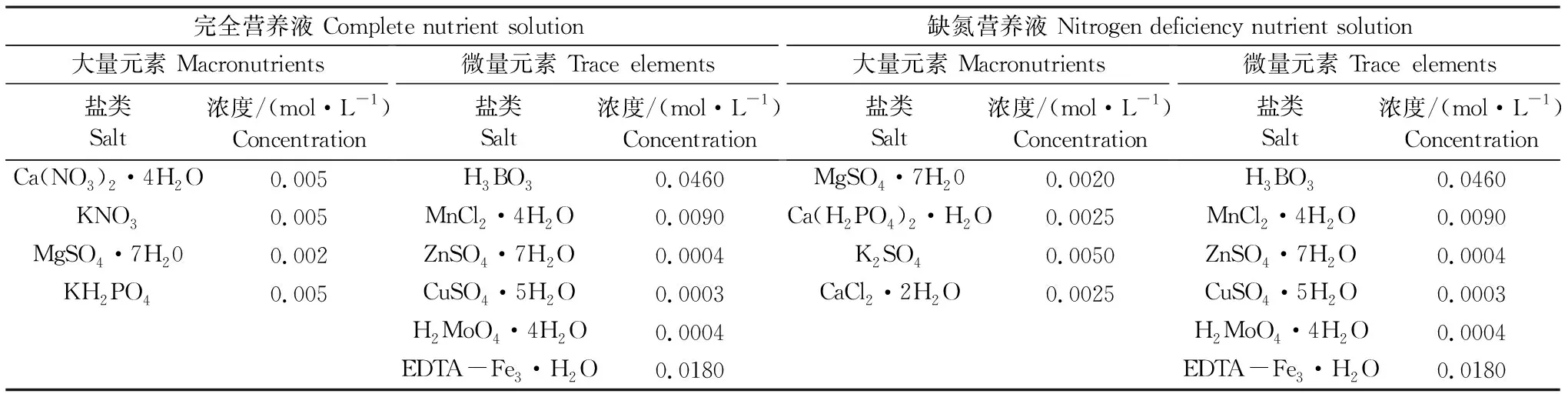
表2 营养液的成分Table 2 Composition of nutrient solution
选用直径2 mm及1 mm的石英砂,先用水浸泡洗涤,至水色呈无色透明后用3%盐酸溶液浸泡7 d,再用蒸馏水洗涤至没有Cl-为止。取直径2 mm和1 mm的石英砂等量混匀后装入内径15 cm、高13 cm的盆钵中,盆钵有排水孔和底座。装盆后,每盆均匀播种50粒种子,出苗后定苗至30株·盆-1。培养期间,采用间歇性浇灌补充营养液和水分。营养液采用砂面浇灌法,浇灌量及频率根据植株大小和环境温度调控至适宜程度。盆钵内的水分调控采用重量法,使水分含量为砂子最大持水量的70%~75%,水分的补给采用砂面浇灌和底部浇灌相结合。处理中涉及由完全营养液换用缺氮营养液时,先用去离子水充分冲洗待换盆钵,之后再浇灌缺氮营养液。植物培养于甘肃农业大学玻璃温室内。
试验二:氮肥施用时期试验,通过土培试验于甘肃农业大学温室进行。共设6个处理:F0:不施氮肥;F1:氮肥0.2 g·kg-1土壤,全部基施;F2:氮肥0.2 g·kg-1土壤,2/3基肥+1/3现蕾期追肥;F3:氮肥0.2 g·kg-1土壤,1/2基肥+1/2现蕾期追肥;F4:氮肥0.2 g·kg-1土壤,1/6基肥+5/6枞形期追肥;F5:氮肥0.2 g·kg-1土壤,1/6基肥+1/3枞形期追肥+1/2现蕾期追肥。氮肥为尿素,用量以纯N计;各处理均施磷肥(P2O50.2 g·kg-1土壤)和钾肥(K2O 0.15 g·kg-1土壤),肥料品种分别为过磷酸钙和硫酸钾,均全部作为基肥施入。
过2 mm筛的土壤13 kg,与基肥充分混匀后装入内径32 cm、高24 cm的盆钵中。装盆后灌足水分,待水分含量下降至适宜水平时进行播种,出苗后定苗至46株·盆-1。培养期间控制土壤水分含量为田间持水量的75%~80%,定期调换盆钵的位置。分别于枞形、现蕾、青果和成熟期采样,每个处理均种植24盆,重复6次,共计144盆。
1.3 测定指标与方法
干物质:烘干法。
全氮:H2SO4-H202消煮,半微量凯氏定氮法测定[19]。
籽粒产量及其构成因素:成熟期收获计产。
1.4 数据处理与统计分析
采用Microsoft Excel 2003、SPSS 17.0统计软件进行数据处理和统计分析。
2 结果与分析
2.1 阶段性缺氮胁迫对亚麻干物质积累及籽粒产量的影响
2.1.1 阶段性缺氮胁迫对亚麻干物质积累的影响 缺氮胁迫对亚麻干物质积累的影响因胁迫时期而具有较大差异(图1,图2)。
T1处理下,枞形期各器官及整株的干物质量均与CK无显著差异,至现蕾期,根干质量较CK增加68.6%,而茎、叶、花蕾的干物质量较CK分别降低31.4%、20.7%、35.2%,表明幼苗时期缺氮胁迫的影响因苗龄小、需氮少而具有一定的滞后性,在胁迫结束的枞形期并未体现出来,但花蕾显现时已突显,因枞形前后是根系建成的关键时期,缺氮使较多的光合产物被根系利用,地上部分的分配较少,故而根干物质量显著增加而茎、叶等干物质积累量下降,根冠比显著增加,是CK的2.3倍。因现蕾时光合源茎、叶较CK的劣势,至盛花期,T1的根、茎、叶及花蕾干物质量分别比CK降低了23.8 %、43.8%、17.7%、27.3%,但器官上的差异除茎秆外呈现比现蕾期缩小的趋势,根冠比仅是CK的1.1倍,远低于现蕾期的差异。至成熟期,T1除根系、蒴果的干物质量较CK降低了15.5%和18.1%之外,茎、叶及整株与CK间均无显著差异。可见,枞形期之前的缺氮胁迫对干物质积累的影响表现滞后,且其抑制作用随枞形之后持续的复氮而逐渐缩小,虽复氮的前、中期未能消除这种负面影响,但最终使地上的营养器官产生了等量补偿。
T2处理和CK相比,从胁迫结束的现蕾期开始,缺氮对各器官的生长抑制均显著显现,至成熟期也未能恢复到对照的水平。其根、茎、叶和生殖器官干物质积累量较CK的现蕾期分别降低了11.5%、22.0%、50.2%、77.0%,盛花期分别降低了40.4%、34.5%、49.5%、68.8%,成熟期分别降低了48.1%、9.8%、30.3%、26.6%。可见,因枞形期后植株开始快速生长,氮素营养需求增加,枞形期~现蕾期缺氮胁迫对生长的抑制作用反应快速;复氮初期,其对根、茎的抑制效应仍在加剧,盛花期根、茎干物质积累量的降低幅度大于现蕾期;随着复氮的持续进行,至成熟期,地上部分的抑制幅度较盛花期降低,但对根系生长的抑制幅度仍和盛花期近似。枞形期~现蕾期的缺氮胁迫对各器官均产生了严重的不足补偿[11],即不能完全补偿缺氮造成的干物质积累量损失。
和CK相比,生殖生长阶段不同生育时期的缺氮胁迫均造成各器官生物量的显著降低,复氮亦未能消除其对生长的抑制作用;成熟期时,根、茎、叶及蒴果的干物质积累量T3分别较CK降低了43.4%、25.8%、19.2%、35.1%,T4分别降低了41.4%、17.1%、38.3%、49.7%,T5分别降低了49.1%、25.5%、44.8%、58.8%。T3、T4处理下两个时段的胁迫对器官生长均造成了显著的影响,对叶片和蒴果生长的抑制花前胁迫小于花后胁迫,对茎秆则刚好相反;二者地上部分干物质积累的降低幅度均呈现蒴果大于茎、叶的趋势,差值均在10%以上;但两种胁迫处理下的整株生物量无显著差异。T5的胁迫历时为T3、T4之和,但其茎秆的干物质量与T3无显著差异、叶片的干物质量与T4无显著差异,表明生殖生长阶段茎秆的干物质积累量与氮源供给的关联程度花后低于花前,而叶片则相反。
缺氮胁迫期间干物质积累量对胁迫阶段响应的敏感性见图2。T1、T2、T3、T4分别为播种~枞形期、枞形期~现蕾期、现蕾期~盛花期、盛花期~成熟期缺氮,T1的播种~枞形期干物质积累量与CK间无显著差异;T2的枞形期~现蕾期干物质积累量显著小于CK;T3的现蕾期~盛花期、T4的盛花期~成熟期干物质积累量均因缺素导致叶片等的衰亡而为负值,且T4的盛花期~成熟期干物质积累量降低程度远大于T3的现蕾期~盛花期。这表明油用亚麻对枞形期之前的缺氮不敏感,对枞形期之后氮素亏缺阶段敏感程度为盛花期~成熟期>现蕾期~盛花期>枞形期~现蕾期。
2.1.2 阶段性缺氮胁迫对亚麻籽粒产量的影响 氮素缺乏阶段影响亚麻的产量及其构成因素,缺氮胁迫时各产量指标均呈下降趋势(图3),且随胁迫时期的后延,单株蒴果数、单果籽粒数及单株籽粒产量的降低幅度均呈增加趋势。与CK相比,T1处理的蒴果及籽粒形成未受到显著抑制,籽粒产量降低了18.02%;而T2、T3、T4、T5的单株蒴果数分别降低了20.00%、36.67%、46.67%、43.33%,单果籽粒数分别降低了21.37%、38.46%、49.57%、50.43%,单株籽粒产量分别降低了26.54%、35.06%、38.83%、48.00%。上述3个指标中,T1、T2均显著大于T3、T4、T5,T1均显著大于T2,表明缺氮胁迫对产量及其构成因素的影响为营养生长时期小于生殖生长时期,营养生长阶段枞形前期小于枞形后期。T3与T4的单株蒴果数无显著差异,单果籽粒数后者比前者显著降低了18.06%,但单株籽粒产量亦无显著差异;而亚麻开花早晚对同一蒴果内籽粒数及粒重均有显著影响,一般早开花比晚开花者籽粒多、千粒质量高[20],根据此亚麻产量形成特性,结合前述T3、T4单株蒴果数、单果籽粒数和单株籽粒产量的差异显著性表现,说明盛花期~成熟期的氮素供给对提高籽粒的形成能力及其充实度至关重要,特别是后者。
各缺氮阶段处理中,复氮对籽粒产量均未产生等量补偿,但不足补偿的不足程度以T1最轻,缺氮减产的敏感程度以生殖生长阶段中花前或花后缺素最为严重。
2.2 氮肥施用时期对亚麻氮素及干物质积累的影响
2.2.1 氮肥施用时期对植株氮素积累量的影响 氮肥施用时期影响亚麻植株体内的氮素积累量(表3),且处理间的差异幅度为生殖生长期高于营养生长期,生殖生长的前中期高于后期。

表3 氮肥施用时期对植株氮素积累量的影响/(mg·株-1)Table 3 Effect of nitrogen application stage on nitrogen accumulation of oil flax plant/(mg·plant-1)
枞形期,植株氮素积累量占生育期积累总量的比例除F0达到30.44%之外,其余5个处理为19.29%~23.64%,即20%左右,表明不施氮肥显著提高了播种~枞形期氮素吸收量占生育期的比例。虽然该时期氮素积累比例较低,但基肥比重高低的影响显而易见。F1处理的氮素积累量除与F2处理差异不显著之外,较F0降低了11.52%,较F3、F4和F5分别降低了14.67%~15.79%,表明氮肥全部作基肥不利于枞形期之前氮素的吸收利用,甚至低于不施氮肥者对土壤环境氮的汲取能力;基肥比例控制在2/3及以下时,比例的高低不影响枞形期的氮素积累。
现蕾期时,0.2 g·kg-1的氮肥用量中,施入量占比为F1100%、F22/3,F31/2、F4100%、F51/2,而5个施氮处理植株的氮素积累量呈现F4=F5>F3>F1>F2的趋势。F4、F5较其他3个处理的氮素积累量增加了24.33%~51.49%,即便F5的氮肥已施入量尚低于F1和F2;同等施入量条件下,F4比F1的积累量增加了33.33%,F5比F3的积累量增加了26.47%,表明枞形期追肥较不追肥显著促进了枞形期~现蕾期的氮素吸收量,即使施入总量较低的情况下,播种及枞形时两次施入的氮素吸收效果仍优于播种时一次施入,可使枞形期~现蕾期的氮素积累量增加40.06%~87.31%。
所有施肥处理的总肥料量已于青果期全部施入土壤,但F2、F3、F5有现蕾期追肥而F1和F4没有。青果期及成熟期的氮素积累量,均呈现F5、F3、F4>F1=F2的趋势,青果期F3和F5之间无显著差异但较F4增加了8.04%~10.92%,成熟期F3和F4之间无显著差异但较F5降低了15.78%。现蕾期~青果期的氮素积累量,F1、F2、F3、F4、F5分别为3.07、3.82、5.25、3.07、4.05 mg·株-1,现蕾期追肥的F2、F3、F5高于不追肥的F1和F424.43%~31.92%。F3和F4之间,青果期前者比后者增加了8.04%而成熟期二者无显著差异,F3和F5之间与之相反,成熟期后者比前者增加了18.74%。以上表明,基肥占比达到1/2以上时不利于终末植株体内的氮素积累,基肥≤1/2、剩余肥料于枞形或现蕾期一次或分两次施入,可使成熟期的植株氮素吸收量增加10.71%~37.50%;现蕾期是改善植物氮素营养的关键追肥时期,枞形期追肥也具有较好的促进氮素积累作用。
2.2.2 氮肥施用时期对亚麻不同阶段氮素积累比例的影响 氮肥施用时期不仅影响植株体内氮素的最高积累量(表3),也影响不同阶段氮素积累比例的高低(图4)。
F0的吸氮比例,播种~枞形期、青果期~成熟期显著高于其他处理,而枞形期~现蕾期、现蕾期~青果期显著低于其他处理,致使F0的吸氮比例在4个时段间的差异远小于其余5个处理,即不施氮肥时,生育期内氮素的吸收比例较为均衡。
施氮的5个处理中,播种~枞形期、青果期~成熟期两个时段的吸氮比例分别低于24%和10%,而枞形期~现蕾期、现蕾期~青果期两时段的比例均为30%~48%,表明枞形期~青果期是亚麻氮素积累的关键期,60%~80%左右的氮素于此期吸收,氮肥的施用时期不影响此吸收比例趋势。但基肥、追肥比例及追肥时期则显著影响氮素积累主要时段内的氮素积累比例。F4处理下枞形期~现蕾期的积累比例较F5增加12.18%、较F2及F3分别增加了59.69%和51.39%,F3处理下现蕾期~青果期的积累比例较F2、F5分别增加了12.01%和38.83%,而较F4增加了58.13%,即枞形期追肥增加了枞形期~现蕾期的吸氮比例,现蕾期追肥增加了现蕾期~青果期的吸氮比例,表明当期追肥对增加氮素吸收比例的效果立竿见影,且其增效随追肥比例的增加而增加。
2.2.3 氮肥施用时期对亚麻干物质积累的影响 不同施肥时期处理下干物质的积累量见图5。枞形期亚麻根、茎、叶及整株的干物质积累量均呈现F0≥F4、F5≥F3≥F2>F1的趋势。根干物质积累量表现为F2、F3较F0、F4、F5显著降低了13.26%~19.71%,F1又较F2、F3显著降低了14.93%~18.31%;茎干物质积累量表现为F3较F0、F2较F3、F1较F2处理均显著降低了10%左右;叶干物质积累量表现为F1较其他4个施肥处理显著降低了8.84%~12.42%。F0、F4、F5的根冠比为1.09~1.15,而F3、F2、F1分别为0.94、0.93和0.90。可见,随基肥用量的增加,器官干物质积累量呈降低趋势,对根的影响大于茎和叶,表明较高的基肥用量抑制了干物质的积累,特别是不利于根系的生长,使得根冠比显著降低。
现蕾期已施入肥料量相等的情况下,器官干物质积累量呈现F4>F1和F5>F3的趋势,整株的干物质积累量表现为F4较F1、F5较F3处理分别提高了31.70%和16.57%,表明枞形期追施氮肥具有显著的促生作用。F5的根、茎、叶、整株干物质积累量较F4分别提高了22.79%、13.66%、8.62%、15.17%,表明同等小剂量的基肥条件下,枞形期追肥的促生作用小剂量高于大剂量。
各器官及整株干物质积累量于青果期及成熟期表现为F1、F2间差异不显著,F3、F5大于F1,且F5大于F2的趋势,表明现蕾期追肥具有促生作用,且追肥比例1/2时促生效果优于1/3;F5的植株干物质积累量较其他4个施肥处理分别增加了11.18%~25.53%和7.89%~22.02%。
2.2.4 氮肥施用时期对亚麻籽粒产量的影响 从表4可以看出,F1、F2的籽粒产量、蒴果数及有效蒴果数、千粒重低于F3、F5,表明基肥用量高不利于提高产量,主要是不利于蒴果的形成和籽粒的增重。F4的产量低于F3和F510.92%~18.71%,F5的产量高于F3和F49.59%~23.02%,表明现蕾期追肥对籽粒产量的提高至关重要,而枞形期小剂量的追肥给现蕾期追肥增产潜能的发挥奠定了坚实的基础。
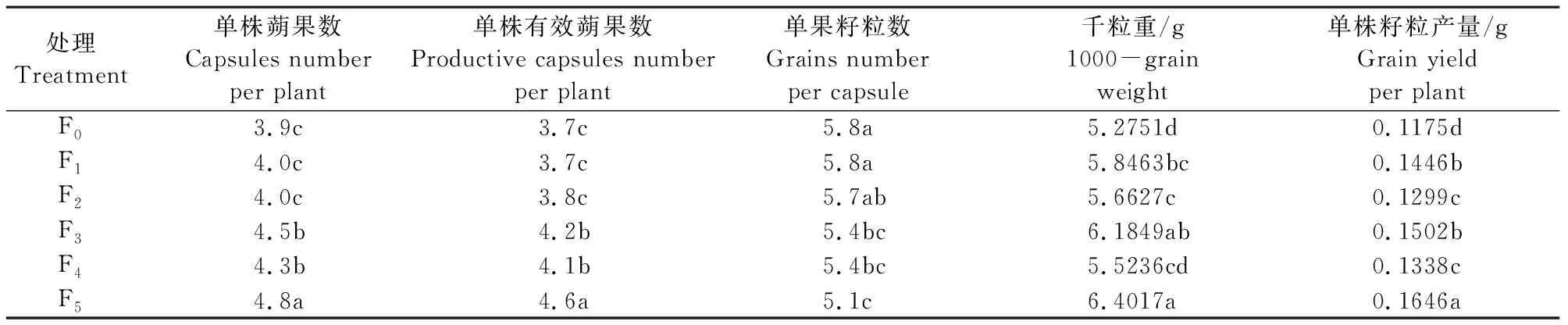
表4 氮肥施用时期对亚麻产量的影响Table 4 Effect of nitrogen application stage on yield of oil flax
3 讨 论
氮是核酸、蛋白质、酶、叶绿素、维生素和激素等有机物质的主要组成成分[21],直接或间接影响植物的光合作用、呼吸作用、蛋白质合成以及脂肪代谢[22],是限制植物生长发育和产量形成的首要矿质营养元素。油料作物种子富含油脂和蛋白质,对氮的需求较高,致使氮肥成为油料作物种植中主要的能量利用和输入消费物质[23]。在氮、磷、钾三要素中,以缺氮胁迫对油菜生长发育及产量形成的影响最为显著[24]。亚麻籽含有35%~45%的油脂和20%左右的蛋白质[1-2],氮素营养的盈亏在籽粒产量形成和品质调控中举足轻重[6-8,20]。
3.1 油用亚麻的氮素吸收与积累
枞形期之前,油用亚麻地上部分的生长较慢,氮素吸收量较少。枞形期之后营养体的大量增殖使得氮素吸收显著增加。现蕾期前后是油用亚麻氮素吸收强度较大的阶段[25],积累量主要集中在生殖生长阶段[25],盛花期~成熟期的氮素吸收量可高达全生育期积累量的67.67%~81.24 %[8]。本研究也例证了生殖生长阶段占比30%~50%以上的积累量。生殖生长阶段是油用亚麻氮素营养的关键期,保证该期的氮素供给对提高籽粒产量至关重要。
砂培试验中现蕾期~盛花期和盛花期~成熟期缺氮时,其干物质积累量均为负值,充分说明了保证生殖生长期氮素营养的重要性。以此为基础设计的土培追施氮肥试验进一步验证了现蕾期追肥具有显著的干物质及籽粒产量补偿效应。
3.2 作物缺氮胁迫阶段的影响及复氮的生长补偿
土壤养分亏缺是作物生长发育过程中较为常见的一种逆境胁迫,作物受到胁迫伤害后会产生一种积极的自我调节能力,即补偿效应,Maschinski和Whitham[26]按照补偿程度将其分为超补偿、等量补偿和不足补偿。缺氮后恢复供氮,作物生长趋向正常,但表型性状及干物质积累量等的恢复程度因作物和胁迫类型而异。
大多数幼苗期的缺氮胁迫,当胁迫解除后根系甚至植株生长通常能恢复或超越正常水平。油菜幼苗缺氮胁迫后,以及香蕉苗缺氮后复氮时,茎叶干物质量虽低于对照,但根干物质量产生了等量补偿,根冠比增加[24,27];菘蓝幼苗的缺氮及复氮处理,根、叶干物质量均产生了等量补偿[28];亚麻播种后至8叶间的缺氮胁迫,幼苗干物质积累较完全营养液增加了151.3%[9];本研究中,亚麻枞形期之前的幼苗期缺氮,短期复氮后,虽茎、叶和整株干物质量恢复不到正常处理水平,但根系干物质量及根冠比显著高于对照,产生了超越补偿;连续复氮后,成熟期的营养器官干物质量产生了等量补偿,印证了幼苗期缺氮胁迫通常伤害较小、生长容易恢复的观点。幼苗期是根系建成和生长的关键时期,缺氮下的生存策略使较多的光合产物分配到根系,致其生长较快,根冠比增大,形成较大的根系扩大吸收能力,应对氮素亏缺[29]。
作物生育期内氮素亏缺的敏感期及不同时期缺素的影响程度因作物而差异较大。双季稻生产中的早稻及晚稻均以分蘖期和穗分化期为氮素亏缺的敏感期,比生育期内常量供氮的对照减产24.4%~34.2%;抽穗期及乳熟期胁迫时产量和对照间无显著差异[30]。冬小麦的阶段氮素亏缺,越冬期亏缺造成的干物质量降低最大,较全生育正常供氮的对照降低44.3%,复氮也未能消除这种影响,致使籽粒产量降低了10.3%;拔节期亏缺亦显著降低了生物学产量和籽粒产量,而抽穗期和成熟期亏缺者影响不显著[31]。虽然水稻和冬小麦的生殖生长期缺氮未造成显著的减产,但菊花生殖期缺氮可减产29.8%,虽然对其营养器官的生物量影响不大[32]。油用亚麻属于“双库”的油料作物[33],对氮素营养的依赖性较强,苗高8~10cm时开始持续缺氮20 d,即可造成器官及植株的干、鲜质量降低,根、茎、叶、整株的干物质量分别显著降低了33.3%、49.7%、100.0%、45.0%[10];本研究也表明枞形期~现蕾期的缺氮具有显著的干物质积累抑制和籽粒产量降低作用,恢复供氮后直至终末,亦未能消除此影响。本研究中,生殖生长阶段某一生育时期缺氮和生殖期缺氮,均造成了植株生物量及籽粒产量的显著降低,且二者的减产均显著高于枞形期~现蕾期缺氮者,同菊花上生殖期缺氮减产严重的研究结果相同[32]。
生长补偿效应的形成需要一定的时间积累[34],且生物量积累以养分吸收为基础。亚麻氮素营养具有生殖生长阶段吸收比重高的特点,因此,生殖生长阶段一定时长的缺氮胁迫造成减产程度较高,尤以盛花期~成熟期缺氮对植株的干物质积累及产量抑制的敏感程度最高。
3.3 作物氮肥追施的效果
随着施氮水平的增加甚至过量施用[12],我国大田作物的供氮能力普遍增强[35],氮肥施用上总量控制、基肥比例适当下调并分次追肥成为许多作物进一步增产的有效途径[35]。改全部基施的习惯方式为氮肥分次施用并适当后移,显著提高了红壤旱地花生的生物量、产量及氮肥利用率,50%基施+30%花针期追施+20%结荚期追施较传统施肥增产9.8%[36]。相同氮肥用量下,夏玉米[37]、糜子[38]的产量随施氮次数的增加不断提高,谷子三次施肥较两次施肥增产15.07%[39]。沿黄灌区的油用亚麻施氮量达180 kg·hm-2时,氮肥用作基肥的比重越大,籽粒产量越低;基追比为3∶7时于枞形、现蕾和盛花期分三次按5∶3∶2 的比例追肥,产量较全部基施提高25.23%,较基追比7∶3者提高22.47%[40]。旱区的油用亚麻,施肥量150 kg·hm-2时,全部基施、2/3基肥+1/3现蕾期追肥、1/2基肥+1/2现蕾期追肥、1/3基肥+2/3现蕾期追肥、1/3基肥+1/3分茎期追肥+1/3现蕾期追肥5种方式中,现蕾、盛花、青果、成熟各时期的植株干物质及氮素积累量、籽粒产量均以三次施肥者最高,成熟期较其他处理提高4.1%~16.3%、13.4%~52.3%、5.2%~16.20%[41]。本研究中,1/6基肥+1/3枞形期追肥+1/2现蕾期追肥的方式,较其他施肥方式的氮素及干物质最大积累量、籽粒产量提高7.17%~37.5%、7.89%~22.02%、9.59%~26.71%。综上,油用亚麻应适当降低基肥的用量,加强盛花前的追肥。基肥≤1/3、剩余肥料于枞形(或分茎)、现蕾期分两次施入,并使现蕾期追肥占1/2左右,是油用亚麻较为合理的氮肥运筹方式。
4 结 论
1)油用亚麻的氮素积累主要集中于枞形期~青果期,枞形期之前和青果期之后的吸收较少;追施氮肥对产量形成至关重要。
2)只有枞形期之前的缺氮胁迫,复氮后油用亚麻的植株干物质积累量产生了等量补偿;对植株的干物质积累及产量,盛花期~成熟期缺氮抑制的敏感程度最高。
3)降低基肥用量至1/2以下,在枞形期小剂量追肥的基础上,重视蕾花期的追肥,可同步油用亚麻的氮素需求与土壤、肥料的养分供给,提高干物质积累及籽粒产量。
猜你喜欢
山东农业科学(2024年3期)2024-06-11 20:39:45
特种经济动植物(2023年10期)2023-10-25 07:54:36
纺织科学研究(2023年9期)2023-10-23 11:18:12
南方农业·上旬(2023年7期)2023-10-09 08:48:44
江苏农业学报(2023年2期)2023-06-08 23:29:47
中国农业大学学报(2022年3期)2022-05-19 00:40:44
土壤学报(2022年1期)2022-03-08 08:30:40
中国麻业科学(2018年6期)2018-04-09 11:22:12
纺织科学研究(2017年4期)2017-05-17 04:00:02
纺织服装流行趋势展望(2016年6期)2016-05-04 03:52:43
