
毛翠雀花花序内的性分配和繁殖成功
2021-09-13 05:01张新安宇梦史长莉米兆荣张婵
广西植物 2021年8期
张新 安宇梦 史长莉 米兆荣 张婵

摘 要:兩性花植物花序内不同位置的性分配和繁殖成功一般存在差异,通常认为资源竞争、结构效应和交配环境是形成这种差异的主要原因。为了研究雄性和雌性繁殖资源在花序内不同位置间的最优分配问题,该文以青藏高原高寒草甸典型高山植物毛翠雀花为材料,通过比较花序内不同位置的花部特征和种子性状,对其花序内的性分配模式和雌性繁殖成功进行研究,并通过观察传粉者运动特点以及人工去花和补授花粉实验,探讨花序内资源竞争和交配环境对繁殖资源分配的影响。结果表明:(1)不同位置间的雄蕊数、雄蕊鲜重/雌蕊鲜重、花粉数及花粉胚珠比从花序基部到上部显著增加,而雌蕊鲜重和胚珠数逐渐减少,表现出上部花偏雄的性分配;上部花的结籽率显著低于基部花和中部花,不同位置间的发育种子数/果实和发育种子重/果实随着花位置的升高而显著降低,说明基部花具有更佳的雌性繁殖成效。(2)去花处理后,剩余果实的单个种子重/果实显著增加,但发育种子数/果实没有显著增加;而给上部花人工补授异花花粉后,位置间结籽率的差异消失,说明传粉限制而非资源竞争导致了花序内位置依赖的种子生产模式。(3)毛翠雀花雄性先熟的开花特征,以及传粉者苏氏熊蜂从花序基部到上部的定向访花行为,导致了花序内交配环境的变化。综上结果表明,毛翠雀花花序内的性分配和繁殖成功差异是对交配环境适应的结果,对其在高山环境中实现雌雄适合度最优化具有重要意义。
关键词:毛翠雀花,花序内差异,性分配,繁殖成功,交配环境
中图分类号:Q945.5
文献标识码:A
文章编号:1000-3142(2021)08-1324-09
Abstract: Intra-inflorescence variation in floral sex allocation and reproductive success are common in hermaphroditic plants, and several non-exclusive hypotheses, including resource competition, architectural effect and the mating environment, have been formulated to explain this phenomenon. In order to investigate the optimal allocation mode of male and female reproductive resources between different positions in the inflorescence, through comparison of floral and seed traits, the floral sex allocation and female reproductive success within inflorescences of Delphinium trichophorum (Ranunculaceae), a typical alpine plant of Qinghai-Tibet Plateau was investigated. Observation of pollinator movements, artificial flower-removal experiment, and hand-supplemental pollination were also used to investigate the effects of resource competition and mating environment on allocation of reproductive resource in the inflorescence. The results were as follows: (1) From bottom to upper positions of inflorescence, stamen number, stamen fresh mass/pistil fresh mass, pollen number and pollen/ovule ratio all increased significantly, while pistil fresh mass and ovule number declined gradually, which showed upper flowers performed male-biased function. Seed set ratio of upper flowers were significantly lower than those of bottom and intermediate flowers. The number and mass of mature seed per fruit decreased significantly with the increase of flower position, indicating greater female reproductive success of bottom flowers. (2) The flower-removal experiment improved single seed mass of the remaining fruits, but did not increase the number of mature seeds per fruit. There were no differences of seed set ratio among positions after hand-supplemental pollination of upper flowers, which suggested that the position-dependent pattern of seed reproduction within inflorescences was attributed to pollen limitation rather than resource competition. (3) The protandry of D. trichophorum and the directional visiting behaviors (from bottom to upper) of pollinator Bombus sushikini led to variations of mating environment within inflorescences. All the above results indicate that intra-inflorescence variation in floral sex allocation and reproductive success of D. trichophorum are results of adaptation to mating environment, and this strategy is of great importance to optimize the male and female fitness for the species.
Key words: Delphinium trichophorum, intra-inflorescence variation, sex allocation, reproductive success, mating environment
自然界中,大约90%的开花植物是雌雄同体(Ainsworth, 2000),其有性繁殖的资源分配可以分为两个部分:雌性功能和雄性功能。两性功能之间存在一种权衡关系,即对一种性别投入的增加会导致对另一种性别投入的减少(Charnov, 1982)。性分配(sex allocation)理论主要是从雌雄功能的角度考虑个体的适合度,研究繁殖资源在两性功能之间的最优分配问题(赵志刚等,2005)。性分配模式在个体间、个体内甚至是同一花序的不同花之间均可能存在差异(Kudo et al., 2001),通常花序内雌性和雄性器官大小、数量或重量的变异都被认为是性别表达的改变(樊宝丽等,2008)。常见的变化模式是顶向开花的植物,其基部较上部具有较大的花,较大或较多的胚珠、果实和种子以及较低的花粉胚珠比等,即基部位置在资源投资上偏向于雌性,上部位置偏向于雄性(Ashman & Hitchens, 2000;Buide, 2004, 2008)。而基向开花的植物,一般呈相反的模式。
造成花序内性状表达和性别分配差异的原因是多方面的,主要有以下几种解释。(1)资源竞争(resource competition):早期开放或处于优势位置的花(靠近营养来源和光合产物来源)对资源的吸收具有更大的竞争优势(Thomson, 1989;Guitián et al., 2001)。资源竞争所造成的不同位置间的资源差异是花序内雌雄功能分化的重要原因,因为资源有效性对雌雄功能的影响不同,相对于雌性功能,维持雄性功能的成本较低(Torices et al., 2019)。因此,理论预测在资源充足的花序位置具有偏雌的性分配,而资源限制的位置具有偏雄的性分配(Brunet & Charlesworth, 1995)。(2)结构效应(architectural effect):花序内不同位置间花的形态和繁殖潜力的差异是由花序轴本身固有的结构特征造成的,不随植株资源状况的變化而变化(Diggle, 1995, 2003)。结构效应认为,在植物生长发育过程中支撑结构直径的变化(Midgley & Bond, 1989)或远端维管结构的萎缩(Wolfe & Denton, 2001)会导致母体资源运输效率从花序基部到顶部逐渐降低,从而成为花部特征以及果实和种子繁育特征的调控机制。(3)交配环境(mating environment):开花时间上的差异或传粉者的定向运动可能导致花序内不同位置间柱头上落置的花粉数量或质量有所不同(Brunet & Charlesworth, 1995;Wiggam & Ferguson, 2005)。Brunet & Charlesworth(1995)认为,植物为了获得最佳的雌雄适合度,较早被访问的花应该对雌性的繁殖投入更多,而较晚被访问的花应该对雄性的繁殖投入更多。
对于不同植物,以上各影响因素的作用存在巨大差异,特别是生长在青藏高原高寒草甸上的典型高山植物会采取怎样的资源分配策略使其在极端环境中实现雌雄适合度最优化是值得深入探讨的问题。根据本课题组前期观察发现,毛翠雀花(Delphinium trichophorum)花序内不同位置的花在花冠大小等花部特征上有所不同,推测该物种花序内不同位置间的繁殖资源分配可能存在差异。因此,本文对毛翠雀花花序内不同位置的性分配模式、传粉者运动特点及繁殖成功进行了研究,拟解决以下3个问题:(1)毛翠雀花雄性和雌性繁殖资源在花序内不同位置间的分配是否存在显著差异;(2)繁殖资源在花序内的分配对其繁殖成功有何影响;(3)毛翠雀花雄性先熟的开花特征,以及传粉者在花序上的运动与其雄性和雌性繁殖资源的分配有何联系。
1 材料与方法
1.1 研究地概况
本研究于2016年7—9月在中国科学院海北高寒草地生态系统研究站(以下简称海北站)(101°12′—101°23′ E、37°29′—37°45′ N)附近开展。海北站位于青藏高原东北隅的青海海北藏族自治州门源县境内,祁连山北支冷龙岭东段南麓坡地的大通河河谷西段。该站地形开阔,海拔为3 200~3 600 m(实验样地海拔高度为3 220 m),四面被祁连山环绕,是典型的高原大陆性气候特征。海北站无明显四季之分,仅有冷暖二季之别,冷季漫长而寒冷,暖季为5—9月。最暖的7月月平均气温为10.1 ℃,最冷的1月为-15.0 ℃,年平均气温-1.6 ℃。年平均降雨量560 mm,暖季的降雨量占全年降雨量的79%。植被类型为青藏高原典型的地带性植被:金露梅(Potentilla fruticosa)为建群种的高寒灌丛草甸和以嵩草属(Kobresia)植物为建群种的高寒嵩草草甸(李英年等,2004)。
1.2 研究材料
毛翠雀花是毛茛科翠雀属的多年生草本植物(图1)。总状花序狭长,长6~30 cm,密生多花,开花顺序为顶向式;萼片5,灰白色、淡蓝色或紫色,花瓣2,退化雄蕊2,雄蕊多数,心皮3。毛翠雀花自交亲和,花期集中在8—10月。
1.3 研究方法
首先根据花序轴的位置选择大小相似的三个区域分为基部、中部和上部(Kudo et al., 2001)。
1.3.1 花序内不同位置的花部特征 在种群开花早期,随机标记大小相似、花数量接近的20个植株。在花即将开放且未散粉时,依次取其主花序上所有花蕾并保存在FAA固定液中(福尔马林∶乙酸∶无水乙醇体积比=5∶5∶90)(Zhang et al., 2012),室内统计花粉数和胚珠数。(1)依据Dafni(1992)的方法,用玻璃棒将花药充分研磨,使花粉粒完全脱离花粉囊,加入少许洗涤剂和50%乙醇定容到5 mL试管中,充分摇匀后用微型移液器取10 μL滴于载玻片上,在40倍光学显微镜下统计花药中的花粉数,重复10次,最后求得每朵花的总花粉数。(2)在载玻片上用镊子解剖每个花蕾的心皮,在体式显微镜下统计每个花蕾的胚珠数。最后用每朵花的花粉数除以其对应的胚珠数,得到花粉胚珠比(Cruden, 1977)。
在种群开花早期,随机标记大小相似,花数量接近的30个植株,按主花序的开放次序,每朵花在花中期(花开放后有一半花药开裂时)取样。将每朵花的雄蕊和雌蕊分解,统计雄蕊数,并用万分之一电子天平称重,获得不同位置花的雄蕊鲜重和雌蕊鲜重。
1.3.2 花序内不同位置的种子生产模式 随机选取大小相近、花数目接近的30个植株,按主花序从基部到上部的次序,首先当每朵花萎蔫后果实开始发育时套网,然后依次按果实成熟次序采集所有成熟果实,最后自然通风条件下干燥3个月。毛翠雀花的发育种子黑棕色,呈大而饱满的四面体形,而其败育胚珠会萎缩成小而干瘪的黄褐色颗粒,根据以上特征将两者区分开来,并分别计数。然后称量发育种子重/果实,并计算单个种子平均重和结籽率,结籽率=发育种子数/(发育种子数+败育胚珠数)。
1.3.3 传粉者运动 在种群盛花期,选择晴朗无风的天气,每天随机标记种群内不同植株上的10个花序,从9:00到17:00之间连续观察并记录访问选定花的所有传粉者及其在花序上的移动方向。
1.3.4 去花对种子性状的影响 随机选取20个植株作为对照组,每一株选择与其匹配的一个植株(大小相似、花数目接近),作为处理组。20个处理植株分别在花后期(花瓣萎蔫)去花,只保留第10、11、12、13、14朵花,比较最终每朵花与对照植株相应的第10、11、12、13、14朵花,得出发育种子数及其重量、单个种子平均重和结籽率的差异。
1.3.5 人工补授异花花粉 随机选取大小相似、花数目接近的30个植株,其主花序“上部”位置的每朵花在柱头张开后人工补授花粉(10 m以外植株的异花花粉)。果实成熟后,统计花序内不同位置果实的结籽率。
1.3.6 数据分析 所有数据用SPSS13.0软件进行统计分析,采用协方差分析比较花序内不同位置间花部特征和种子性状的差异以及补授异花花粉对结籽率的影响;采用双因素方差分析检验去花处理和花序内不同位置及其交互作用对单个种子平均重、发育种子数/果实、发育种子重/果实和结籽率的影响。
2 结果与分析
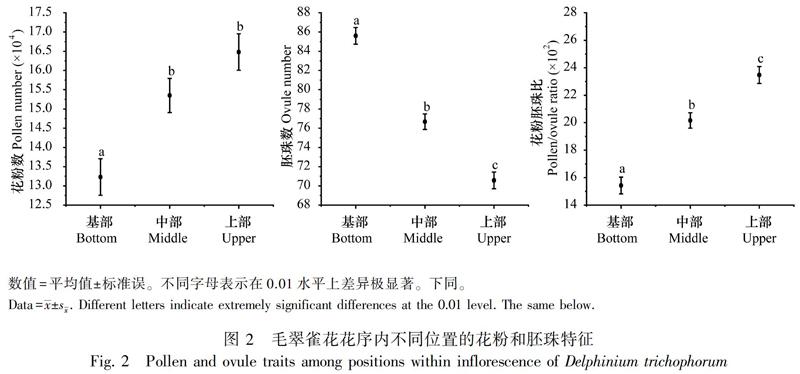
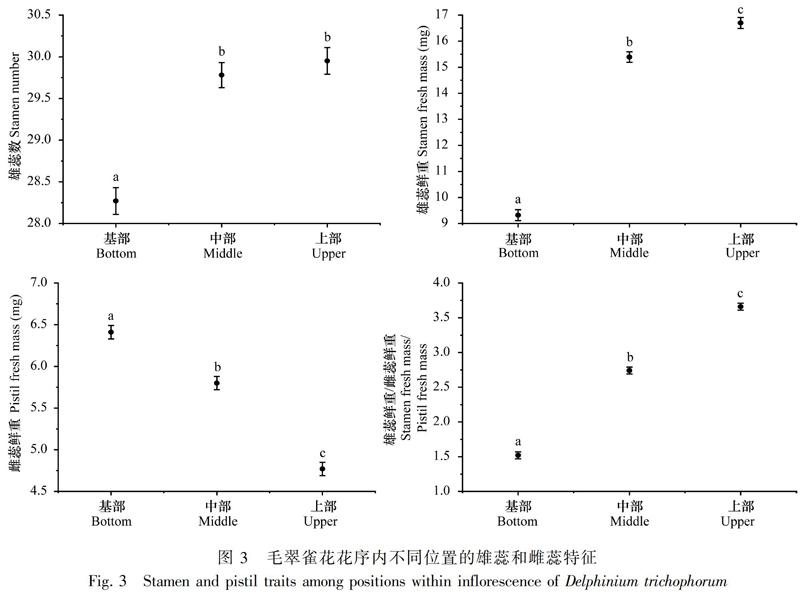
2.1 花序内不同位置的花部特征
毛翠雀花的花部特征在花序内不同位置间存在显著差异。从花序基部到上部,每朵花中的花粉数(P<0.001)和花粉胚珠比(P<0.001)显著增加,而胚珠数(P<0.001)显著减少(图2)。基部花的雌蕊鲜重(P<0.001)较高,而上部花的雄蕊数(P<0.001)、雄蕊鲜重(P<0.001)以及雄蕊鲜重/雌蕊鲜重(P<0.001)较高(图3)。以上结果表明,毛翠雀花花序内不同位置间雌雄繁殖投入存在显著差异,雌性繁殖投入从基部花到上部花显著减少,而雄性繁殖投入显著增加,因此整个花序内表现出上部花偏雄的性分配。
2.2 花序内不同位置的种子生产模式
对花序内种子生产模式的研究表明(图4),虽然花序内不同位置间的单个种子重没有显著差异(P=0.530),但是每个果实的发育种子数(P<0.001)和发育种子重(P<0.001)随着花位置的升高而显著降低,而且上部花的结籽率显著低于基部花和中部花(P<0.001)。以上结果表明,毛翠雀花花序内的基部花具有更佳的雌性繁殖成效。
2.3 传粉者运动
根据观察,苏氏熊蜂(Bombus sushikini)是毛翠雀花在该地区唯一的有效传粉昆虫(图5),它将头部探入花内取食花蜜过程中,胸腹部能够充分接触到伸长的柱头和散粉的花药,从而完成花粉的输入和输出。累计记录329只苏氏熊蜂在花序上的移动方向,其中277只(84.19%)自下而上,11只(3.34%)自上而下,剩余41只(12.46%)访问1朵花后即离开花序。
2.4 去花对种子性状的影响
去花处理后,虽然剩余果实的发育种子重/果实显著增加,这主要是由于单个种子重显著增加,但是,发育种子数/果实和结籽率并没有显著变化(表1)。另外,去花处理对以上种子性状的影响与剩余果实着生的位置无关,因为处理和位置的交互作用不显著。
2.5 人工补授异花花粉
人工补授异花花粉后,花序基部、中部和上部的结籽率分别为0.797±0.003、0.793±0.003和0.789±0.004,花序内不同位置间结籽率无显著差异(P=0.306)。这表明毛翠雀花花序内上部花较低的结籽率是受到了传粉限制的影响。
3 讨论
多数研究结果表明,在植株花序水平上,基部的花(或早期开放的花)往往分配更多的資源到雌蕊或者胚珠的生产上,并且形成更多的果实和种子,而上部的花(或晚期开放的花)对雄蕊或者花粉的投入更多。例如,Ishii & Sakai(2002)对Narthecium asiaticum的研究表明,与基部花相比,顶部花的雄蕊重/(雄蕊重+雌蕊重)和花粉胚珠比更高;Buide(2004, 2008)的研究发现,Silene acutifolia的胚珠数、坐果率、种子数/果实、结籽率和种子重从花序基部到远端显著减少;徐旭剑等(2017) 对云南草蔻 (Alpinia blepharocalyx)的研究也发现,每花胚珠数、结实率和结籽率由花序基部到顶部依次降低,每花花粉胚珠比由基部到顶部依次增加。与以上结果相同,毛翠雀花花序内的花部特征(花粉数、胚珠数、雄蕊数、雄蕊鲜重、雌蕊鲜重)和雌性繁殖成功(发育种子数/果实、发育种子重/果实、结籽率)在不同位置间存在显著差异。上部花具有更多的雄性繁殖投入,而基部花表现出更高的雌性繁殖成效。
3.1 资源竞争
资源竞争假说认为,花序内早期形成的花对母体资源具有优先占领的优势,因此,如果去除早期的花,资源有可能重新分配给后期发育的花和果实。例如,在雄全同株植物喜树(Camptotheca acuminata)的聚伞花序中,去除部分头状花序后,剩余头状花序的花序直径、小花直径明显增加,位置间差异消失(徐申林和刘文哲,2011)。去除自交植物喉毛花(Comastoma pulmonarium)的顶花后,其他位置果实的种子数与对照植株相同位置的果实相比,均呈现增加趋势(张婵等,2011)。然而,对毛翠雀花进行去花处理后,只是增加了剩余果实的单个种子重,而发育种子数和结籽率并没有提高,这说明花序内从基部到上部种子数的减少和结籽率的降低并不是位置间的资源竞争造成的。
3.2 雌雄异熟导致的交配环境变化
在许多高山植物中,交配环境已经被证实是果实生产的关键限制因素之一(Lee, 1988;Krner, 2003)。毛翠雀花的上部花在人工补授异花花粉后,结籽率显著提高,说明上部花受到了传粉限制的影响。根据交配环境假说,在雌雄异熟植物中,雌性阶段花和雄性阶段花在种群中的相对丰富度随着开花季节发生变化,从而使不同时期开放的花在花粉接受和花粉扩散机会上存在差异,进而引起雌性和雄性相对适合度的改变,最终影响花序内的最佳性分配模式(Brunet & Charlesworth, 1995)。
毛翠雀花属于雄性先熟,一朵花中全部花药依次散完粉后,柱头才成熟。对于雄性先熟、顺次开花的植物,当植株上早开的花处于雄性阶段时,种群中交配机会(有效的柱头)缺乏,而当其处于雌性阶段时,种群内有丰富的雄性阶段花,因此,早期花偏雌的性分配具有适应意义,有助于减少花粉损失并提高胚珠异花授粉机会;而晚开的花在雌性阶段缺少异花花粉的几率较大(这可能是造成毛翠雀花花序内上部花结籽率较低的原因之一),从而增加了雄性繁殖投入,有利于花粉的对外扩散(Brunet & Charlesworth, 1995;Brunet, 1996;Buide, 2008)。在毛翠雀花种群中,随着开花时间从早到晚的变化,种群中早期以雄花为主,而晚期以雌花为主,种群交配环境由偏雄向偏雌变化,因此,植株个体水平的性分配模式则相应的由偏雌转向偏雄。花序内早期花偏雌、晚期花偏雄的性分配已经在许多雄性先熟的植物中得到证实(Brunet, 1996;Kudo et al., 2001;Hiraga & Sakai, 2007)。例如,Ishii & Harder(2012)对雄性先熟植物Delphinium glaucum的研究发现,花序内随着开花时间的推移,平均每朵花的雄蕊数增加而胚珠数减少,同时雄性期延长而雌性期缩短。对雄性先熟植物露蕊乌头(Aconitum gymnandrum)的研究发现,花序内较晚开放的花有更长的雄性期和更短的雌性期,性分配在时间上偏雄(李琳等,2016)。Huang et al.(2004)比较了雌性先熟植物华北耧斗菜(Aquilegia yabeana)和雄性先熟植物A. caerulea,结果发现,前者早开的花具有偏雄的性分配而后者相反,支持了交配环境假说。
3.3 传粉者定向运动导致的交配环境变化
除了雌雄异熟,传粉者在花序内的定向访花也会造成花间交配环境的差异,从而影响性分配模式(Brunet & Charlesworth, 1995;Harder et al., 2000)。对于顶向式开花的毛翠雀花,其有效传粉者苏氏熊蜂一般是按照从花序基部到上部的顺序访花。传粉者来访时自下而上的定向运动,使得基部花有更多的机会接受外源花粉,有利于实现雌性功能;而上部花在传粉者离开时能够带走其大量花粉,更有利于花粉扩散、实现雄性功能。因此,为了获得最佳的雌雄适合度,毛翠雀花选择了早期花偏雌而晚期花偏雄的性分配。另外,这种花序内传粉者的定向运动难以避免地会造成同株异花授粉,使得上部花缺少异花花粉,这可能是毛翠雀花上部花结籽率较低的主要原因。在其他一些具有传粉者定向访花特点的植物中,也发现了相似的结果,例如,Kudo et al.(2001)对Corydalis ambigua的研究表明,花序内顶部花种子数的减少是缺少异花花粉的结果,因为主要的访花昆虫(一种熊蜂蜂王)是从花序底部到顶部的顺序访花的。马晓丽等(2011)对蒙古沙冬青(Ammopiptanthus mongolicus)的研究发现,花序内上部花坐果率、结籽率和种子质量较低的主要原因是花序内传粉者定向运动而非资源限制。值得注意的是,毛翠雀花花序内早期花偏雌的性分配能够在一定程度上减少由传粉者定向运动造成的同株异花授粉,降低花粉折损(pollen discounting)和胚珠折损(ovule discounting),从而减少雌雄适合度代价。
4 结论
综上所述,由于雄性先熟的开花特征和传粉者自下而上的访问顺序,毛翠雀花花序内较早受访的花具有较高的异花授粉几率,因此,雌性繁殖投入较高;较晚受访的花其花粉输出幾率较高,而胚珠异花授粉几率较低,因此,雄性繁殖投入较高。这种资源分配模式使毛翠雀花能够在严酷的高山环境中最大限度地实现个体适合度,是长期进化过程中对其交配环境的一种适应机制。
参考文献:
AINSWORTH C, 2000. Boys and girls come out to play: The molecular biology of dioecious plants [J]. Ann Bot, 86(2): 211-221.
ASHMAN TL, HITCHENS MS, 2000. Dissecting the causes of variation in intra-inflorescence allocation in a sexually polymorphic species,Fragaria virginiana (Rosaceae) [J]. Amer J Bot, 87(2): 197-204.
BRUNET J, CHARLESWORTH D, 1995. Floral sex allocation in sequentially blooming plants [J]. Evolution, 49(1): 70-79.
BRUNET J, 1996. Male reproductive success and variation in fruit and seed set in Aquilegia caerulea (Ranunculaceae) [J]. Ecology, 77(8): 2458-2471.
BUIDE ML, 2004. Intra-inflorescence variation in floral traits and reproductive success of the hermaphrodite Silene acutifolia [J]. Ann Bot, 94(3): 441-448.
BUIDE ML, 2008.Disentangling the causes of intra-inflorescence variation in floral traits and fecundity in the hermaphrodite Silene acutifolia [J]. Amer J Bot, 95(4): 490-497.
CHARNOV EL, 1982. The theory of sex allocation [M]. Princeton: Princeton University Press: 63-80.
CRUDEN RW, 1977. Pollen-ovule ratios:A conservative indicator of breeding systems in flowering plants [J]. Evolution, 31(1): 32-46.
DAFNI A, 1992. Pollination ecology: A practical approach [M]. Oxford: Oxford University Press.
DIGGLE PK, 1995. Architectural effects and the interpretation of patterns of fruit and seed development [J]. Ann Rev Ecol Syst, 26(1): 531-552.
DIGGLE PK, 2003. Architectural effects on floral form and function: A review [M]//STUESSY T, HSRANDL E, MAYER V. Deep morphology: Toward a renaissance of morphology in plant systematics. Liechtenstein: Ganter Verlag: 63-80.
FAN BL, ZHAO ZG, MENG JL, et al.,2008. Position-dependent sexual distribution of Aconitum gymnandrum Maxim [J]. Acta Ecol Sin, 28(6): 2909-2915. [樊寶丽, 赵志刚, 孟金柳, 等, 2008. 露蕊乌头(Aconitum gymnandrum Maxim)花序内位置依赖的性分配 [J]. 生态学报, 28(6): 2909-2915.]
GUITIN J, GUITIN P, MEDRANO M, 2001. Causes of fruit set variation in Polygonatum odoratum (Liliaceae) [J]. Plant Biol, 3(6): 637-641.
HARDER LD, BARRETT SCH, COLE WW, 2000. The mating consequences of sexual segregation within inflorescences of flowering plants [J]. Proc Roy Soc London Ser B, 267(1441): 315-320.
HIRAGA T, SAKAI S, 2007. The effects of inflorescence size and flower position on biomass and temporal sex allocation
in Lobelia sessiliflora [J]. Plant Ecol, 188(2): 205-214.
HUANG SQ, TANG LL, YU Q, et al., 2004. Temporal floral sex allocation in protogynous Aquilegia yabeana contrasts with protandrous species: Support for the mating environment hypothesis [J]. Evolution, 58(5): 1131-1134.
ISHII HS, HARDER LD, 2012. Phenological associations of within- and among- plant variation in gender with floral morphology and integration in protandrous Delphinium glaucum [J]. Afr J Ecol, 100(4): 1029-1038.
ISHII HS, SAKAI S, 2002. Temporal variation in floral display size and individual floral sex allocation in racemes of Narthecium asiaticum (Liliaceae) [J]. Amer J bot, 89(3): 441-446.
KRNER C, 2003. Alpine plant life: Functional plant ecology of high mountain ecosystems [M]. Berlin: Springer.
KUDO G, MAEDA T, NARITA K, 2001. Variation in floral sex allocation and reproductive success within inflorescences of Corydalis ambigua (Fumariaceae): Pollination efficiency or resource limitation? [J]. J Ecol, 89(1): 48-56.
LEE TD, 1988. Patterns of fruit and seed production [M]//DOUST JL, DOUST LL. Plant reproductive ecology pattern and strategies. Oxford: Oxford University Press: 179-202.
LI L, LU NN, FAN BL, et al., 2016. Effect of flowering time on floral sexual durations and phenotypic gender in dichogamous Aconitum gymnandrum [J]. Biodivers Sci, 24(6): 665-700. [李琳, 路宁娜, 樊宝丽, 等, 2016. 雌雄异熟植物露蕊乌头开花时间对雌雄功能期及表型性别的影响 [J]. 生物多样性, 24(6): 665-700.]
LI YN, ZHAO XQ, CAO GM, et al., 2004. Analyses on climates and vegetation productivity background at Haibei Alpine Meadow Ecosystem Research Station [J]. Plateau Meteorol, 23(4): 558-567. [李英年, 赵新全, 曹广民, 等, 2004. 海北高寒草甸生态系统定位站气候、植被生产力背景的分析 [J]. 高原气象, 23(4): 558-567.]
MA XL, TAN DY, LI XR, 2011. Variation in floral sex allocation, pollinator movement and reproductive success in Ammopiptanthus mongolicus inflorescences [J]. Biodivers Sci, 19(4): 432-440. [马晓丽, 谭敦炎, 李新蓉, 2011. 蒙古沙冬青花序内性分配的变化、传粉者运动与繁殖成功 [J]. 生物多样性, 19(4): 432-440.]
MIDGLEY J, BOND W, 1989. Leaf size and inflorescence size may be allometrically related traits [J]. Oecologia, 78(3): 427-429.
THOMSON JD, 1989. Deployment of ovules and pollen among flowers within inflorescences [J]. Evol Trend Plant, 3(1): 65-68.
TORICES R, AFONSO A, ANDERBERG AA, et al., 2019. Architectural traits constrain the evolution of unisexual flowers and sexual segregation within inflorescences: An interspecific approach [J]. BioRxiv: 356147.
WIGGAM S, FERGUSON CJ, 2005. Pollinator importance and temporal variation in a population of Phlox divaricata L. (Polemoniaceae) [J]. Amer Midland Nat, 154(1): 42-55.
WOLFE LM, DENTON W, 2001. Morphological constraints on fruit size in Linaria canadensis [J]. Int J Plant Sci, 162(6): 1313-1316.
XU SL, LIU WZ, 2011. Gender expression and gender allocation in Camptotheca acuminata [J]. Chin J Plant Ecol, 35 (12): 1290-1299. [徐申林, 刘文哲, 2011. 喜树花序内性别表达及性别分配 [J]. 植物生态学报, 35(12): 1290-1299.]
XU XJ, SUN S, CAO GX, 2017. Sexual allocation of different parts in the inflorescence of Caulis in Yunnan [J]. Guihaia, 37(3): 335-341. [徐旭剑, 孙杉, 操国兴, 2017. 云南草寇花序内不同部位的性分配 [J]. 广西植物, 37(3): 335-341.]
ZHANG C, HU LJ, WANG Y, et al., 2011. Effects of the position on floral traits and reproductive success of Comastoma pulmonarium (Gentianaceae) [J]. Plant Divers Resour, 33(5): 495-502. [张婵, 胡莉娟, 王赟, 等, 2011. 位置效应对喉毛花(龙胆科)的花特征和繁殖成功的影响 [J]. 植物分类与资源学报, 33(5): 495-502.]
ZHANG C, ZHA SQ, YANG YP, et al., 2012. Effects of the yellow barbs of the staminodes on reproductive success of Delphinium caeruleum (Ranunculaceae) [J]. Biodivers Sci, 20(3): 348-353. [张婵, 查绍琴, 楊永平, 等, 2012. 蓝翠雀花退化雄蕊上的黄色髯毛对其繁殖成功的影响 [J]. 生物多样性, 20(3): 348-353.]
ZHAO ZG, DU GZ, LIU ZJ, 2005. Sex allocation in hermaphroditic plants [J]. Acta Ecol Sin, 25(10): 2725-2733. [赵志刚, 杜国祯, 刘左军, 2005. 雌雄同花植物的性分配 [J]. 生态学报, 25(10): 2725-2733.]
(责任编辑 周翠鸣)
