
基于Cyt b基因的江苏省湖鲚种群遗传多样性和遗传结构分析
2021-09-08 07:45李大命唐晟凯刘燕山谷先坤殷稼雯蒋琦辰张彤晴潘建林
中国农学通报 2021年20期
李大命,唐晟凯,刘燕山,谷先坤,殷稼雯,蒋琦辰,张彤晴,潘建林
(江苏省内陆水域渔业资源重点实验室江苏省淡水水产研究所,南京 210017)
0 引言
湖鲚(Coilia nasustaihuensis)是一种常见的小型鱼类,一般认为其是刀鲚(Coilia nasus)的淡水定居生态类型,隶属于鲱形目、鳀亚科、鲚属,终生生活在江河和湖泊内,与洄游型刀鲚在生态习性和形态上有一定差别[1-2]。湖鲚喜集群,游泳迅速,拓殖能力强,且具有性成熟早、繁殖率高、食性广等特点[2-3]。近几十年来湖鲚资源量迅速增长,逐渐成为淡水湖泊鱼类群落中的绝对优势种群,同时也是湖泊渔业重要的捕捞对象[4-6]。目前湖鲚的研究主要集中在生长规律[7-8]、种群年龄结构[7-8]、食物组成[6,9]等基础生物学及种群数量变动等方面[3,10],而有关湖鲚的遗传学资料相对缺乏。掌握物种的遗传多样性是物种管理保护和开发利用需要了解的重要内容,也是评价生物遗传资源的重要依据。线粒体DNA(Mitochondrial DNA, mtDNA)具有相对较高的进化速率、严格的母性遗传以及几乎不存在重组等特点[11]。细胞色素b(Cytochrome b,Cyt b)基因是一段编码蛋白质的DNA 序列,其进化速度适中,被广泛运用于鱼类遗传多样性和遗传结构研究[12-14]。
迄今大部分针对鱼类的群体遗传学研究主要集中在经济鱼类或者濒危鱼类,而对经济相对偏低的小型鱼类的研究常被忽视[15]。一方面,湖鲚作为一种小型经济鱼类其天然资源面临过度开发利用、种群组成低龄化等问题[3,6];另一方面,湖鲚占据优势会导致鱼类群落结构向小型化发展,进而会导致渔产力下降及影响湖泊生态系统稳定[16-17]。因此,如何科学管理和合理利用湖鲚资源,对于湖泊渔业资源可持续发展及生态系统稳定具有重要意义。本研究采集江苏省长江水系的太湖、滆湖及淮河水系的高邮湖、洪泽湖、白马湖、骆马湖共6 个湖鲚群体,以Cyt b 基因为分子标记,研究江苏省湖鲚群体的遗传多样性、遗传结构及进化历史,以期了解湖鲚的遗传资源现状,进而为湖鲚种质资源的管理及利用提供重要参考。
1 材料和方法
1.1 样品采集
于2016—2018年,用刺网采集江苏省长江流域的太湖、滆湖及淮河流域的高邮湖、洪泽湖、白马湖、骆马湖6个湖泊的湖鲚群体共计224尾个体。采样时间、地点及样本量见表1。将样本用无水乙醇中固定,带回实验室置于-20℃低温冰箱保存。
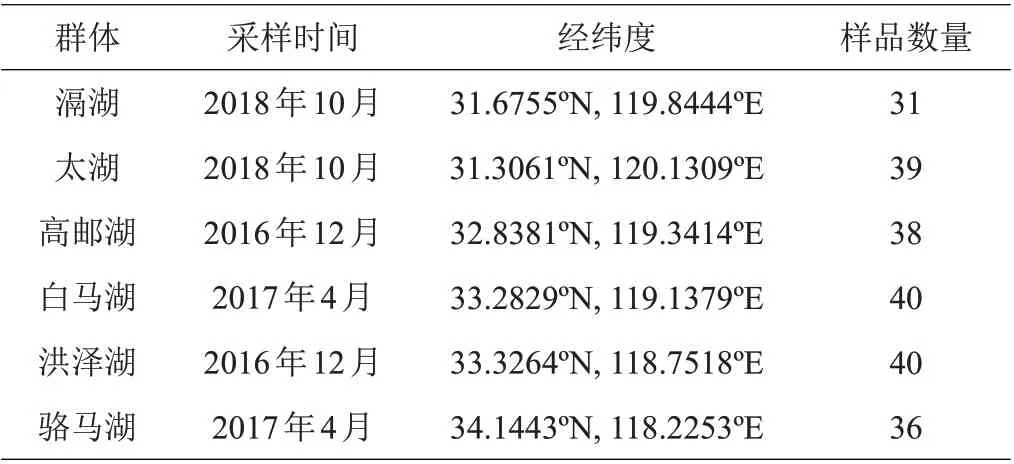
表1 湖鲚样品信息
1.2 基因组DNA提取、PCR扩增和序列测定
购置基因组DNA提取试剂盒,按照说明书步骤和要求提取样品的基因组DNA,所提取的总DNA 溶于TE 溶液,-20℃保存备用。使用分光光度计测定DNA浓度,采用1.0%琼脂糖凝胶电泳检验检测DNA 的完整性。
使用通用引物L14724和H15915扩增Cyt b序列,引 物 序 列 为 L14724:5’- GACTTGAAAAACCACCGTTG- 3’,H15915:5’- CTCCGATCTCCGG ATTACAAGAC-3’[18]。PCR反应总体系为50µL,包括2×TaqMaster Mix(TaKaRa)25µL、上下游引物各1.0 µL(0.01 mmol/L)、DNA模板1.0µL,超纯水补足体积。将反应液充分混匀,10000 r/min冷冻离心30 s,将反应管放入PCR 仪。PCR 反应程序为:94℃变性4 min;94℃变性40 s,53℃退火50 s,72℃延伸60 s,共35 个循环;最后72℃延伸10 min。
PCR产物用琼脂糖凝胶电泳检测,凝胶成像系统拍照记录。用胶回收纯化试剂盒将目标扩增回收纯化,送至上海生工生物工程有限公司进行双向测序。
1.3 数据分析
使用DNAstar 软件和Clustal X 2.0 软件对测序结果进行序列编辑、校对和排序。使用Mega7.0 软件统计碱基组成,计算群体间的遗传距离。以大银鱼(Protosalanx hyalocranius,GenBank 登 录 号:NC_024109.1)和凤鲚(Coilia mystus,GenBank 登录号:MH465026.1)为外类群,采用邻接法(Neighborjoining,NJ)构建单倍型分子系统进化树,系统树中节点的自举置信水平应用自引导估计,重复次数为1000次。使用DNA SP 5.0 软件计算群体的单倍型多样性(h)和核苷酸多样性(π)。利用Arlequin 3.5软件进行分子方差分析,估算遗传变异在种群间和种群内的分布,计算种群间的遗传分化指数及其P值,并采用中性检验以及核苷酸不配对分布来检测种群历史动态。使用Network 软件构建单倍型网络关系图,检测各个单倍型间的进化关系。
2 结果
2.1 Cyt b序列变异及种群遗传多样性
本研究获得224尾样本的Cyt b基因全序列,长度为1141 bp。224条序列共检出62个变异位点,其中单一位点35个,简约位点27个,没有插入/缺失位点。碱基组成分析显示,碱基A、T、G和C的平均含量分别为27.8%、30.6%、14.5%和27.1%,A+T 的含量(58.4%)明显高于G+C的含量(41.6%),表现出明显的A T偏好和反G偏倚特点。
224 尾个体共定义57 种单倍型(H1~H57),其中H49 是6 个种群的共享单倍型,也是占比最高的单倍型(表2)。滆湖、太湖、高邮湖、白马湖、洪泽湖及骆马湖群体分别拥有16种、17种、13种、13种、13种和8种单倍型,且每个种群都拥有数量不等的独享单倍型。6个湖鲚种群的平均单倍型和核苷酸多样性分别为0.783±0.028和0.00260±0.00019,其中滆湖种群的遗传多样性最高,骆马湖种群的遗传多样性最低(表2)。

表2 基于Cyt b基因的湖鲚群体遗传多样性参数
2.2 种群遗传结构分析
6个群体间的遗传距离显示(表3),滆湖与高邮湖群体间的遗传距离最大,为0.00419,洪泽湖和骆马湖群体间的遗传距离最小,为0.00178。长江水系太湖和滆湖群体间的遗传距离为0.00418,淮河水系4个湖鲚群体间的遗传距离为0.00178~0.00240。

表3 湖鲚群体间的遗传距离(对角线下方)和分化系数(对角线上方)
对湖鲚群体间的遗传变异进行分子方差分析(AMOVA)(表4),结果表明,8.55%的变异发生在群体间,91.45%的变异发生群体内,说明分子变异主要来源于群体内个体间,群体间的Fst值统计检验具有极显著性差异(表4)。将6 个种群分为长流水系组群(滆湖、太湖种群)和淮河水系组群(高邮湖、白马湖、洪泽湖和骆马湖种群),进行分子方差分析,结果表明,长江水系组群和淮河水系组群的组间分子变异占总变异的百分比为1.08%,组内群体间分子变异占7.95%,群体内分子变异占90.97%,两组群的分子变异主要来自群体内,两组间的Fct值统计检验无显著性差异,组内群体间的Fsc值和群体内的Fst值统计检验有极显著性差异(表4)。

表4 湖鲚种群间遗传差异的分子方差分析
进一步分析两两种群间的遗传分化(表3),结果显示,滆湖群体与其他5个群体间的Fst均大于0.15,且差异极显著(P<0.01),表明滆湖群体与其他群体间存在极显著遗传差异。除滆湖群体外,其他5 个群体间的Fst均小于0.05,且差异不显著(P>0.05),表明5个群体间没有显著的遗传分化。
2.3 分子系统发育分析
以凤鲚和大银鱼为外类群,基于邻接法构建湖鲚Cyt b基因单倍型之间的系统发育树(图1),结果显示,所有的湖鲚单倍型聚为一支,各种群的单倍型混杂分布,没有形成与地理分布相对应的遗传结构,只有滆湖种群的部分单倍型(H19~H25)形成了置信度较高的进化分支。利用Network软件构建了湖鲚单倍型的进化网络关系(图2),结果显示,单倍型H49 是6 个群体的共享单倍型,位于网络进化图中心,数量也最多,推测可能为湖鲚的祖先单倍型。各种群的单倍型并没有按地理位置形成独立的分支,而是以主单倍型为中心的形成星形放射状结构。滆湖群体的部分特有单倍型(H19~H25)与祖先单倍型相距较远,形成了相对独立的进化分支。单倍型网络结构图进一步支持了分子系统发生树。

图1 基于Cyt b序列构建的湖鲚单倍型系统发育树

图2 湖鲚的Cyt b基因单倍型进化网络图
2.4 种群历史动态分析
对湖鲚群体进行Tajima’sD和Fu’sFs中性检验分析,结果显示,滆湖群体的Tajima’sD和Fu’sFs中性检测值为负值,但统计检验均不显著;其他5个群体的中性检测值为负值,且统计检验有显著或极显差异(表5)。歧点分布图分析显示,滆湖群体的歧点分布图呈多峰型,其他5个群体的歧点分布图呈单峰型(图3)。由此可以推测,除滆湖群体外,其他5个群体进化过程中经历过种群扩张。将6个群体作为一个整体进行分析,结果显示,歧点分布图呈单峰型(图3),中性检测值为负值且有极显著性差异(表5),表明湖鲚群体在历史上经历了显著的种群扩张。

图3 湖鲚种群Cyt b基因序列的歧点分布图

表5 基于Cyt b基因的湖鲚群体中性检验及种群扩张时间估算
根据种群的扩张参数τ来估算群体的扩张时间,线粒体DNA的蛋白质编码基因变异速率为2%每百万年[19]。经计算,太湖、白马湖和骆马湖种群的扩张发生在约0.11 百万年前,高邮湖种群的扩张发生在约0.12百万年前,洪泽湖种群的扩张发生在约0.13 百万年前。整个湖鲚种群的扩张发生在约0.23百万年前(表4)。
3 讨论
3.1 湖鲚种群遗传多样性分析
在线粒体的众多基因中,Cyt b 基因进化速度适中,转换或颠换比例较大,被广泛应用于鱼类系统发育及遗传多样性研究。本研究对6 个湖鲚群体的Cyt b基因进行PCR扩增和测序,分析群体间序列组成及变异。结果显示,所有序列共发现62 个变异位点,变异率为5.43%。湖鲚群体的Cyt b 序列碱基组成基本一致,其中AT的含量(58.4%)明显高于GC含量(41.6%),而碱基G的含量最低,表现出明显的AT偏好性。
单倍型多样度(h)和核苷酸多样度(π)是衡量物种或种群线粒体DNA变异程度的2个重要指标,其值越大,表明群体的遗传多样性就越高[20]。本研究通过Cyt b 基因对江苏省6 个湖泊湖鲚群体进行遗传多样性分析,结果显示,6个群体的单倍型多样性为0.717±0.060~0.875 ± 0.044,核 苷 酸 多 样 性 为0.00180 ±0.00034~0.00425±0.00033,6个群体总体单倍型和核苷酸多样性分别为0.783±0.028 和0.00260±0.00019。Grant 和Bowen 以单倍型多样性0.5 为临界值、核苷酸多样性0.005 为临界值将单倍型多样性和核苷酸多样性之间的关系分为4种类型:低h(<0.5)和低π(<0.005);高h和低π;低h和高π;高h和高π。可以看出,湖鲚种群的遗传多样性属于第二种类型,具有较高的单倍型多样性,表明湖鲚种群具有较丰富的遗传多样性,这可能与湖鲚的栖息生境及种群大小有密切关系。湖鲚性成熟早、繁殖率高、食性广、游泳迅速、适应能力强,具有广阔的生存空间,形成了种群数量庞大的群体,奠定了湖鲚具有丰富遗传多样性的基础。Ma 等[21]报了国内其他湖泊湖鲚群体的遗传多样性,如巢湖、洞庭湖、鄱阳湖等群体也都显示了较丰富的遗传多样性。种群遗传多样性形成与进化历史密切相关,种群具有高h和低π可能受到瓶颈效应的影响[20]。一般来说,种群受遭遇瓶颈效应后数量迅速增加,导致种群单倍型多样性迅速升高,由于核苷酸多样性的积累时间远大于单倍型多样性,因此核苷酸多样性仍处于较低水平。一般用Tajima’sD和Fu’sFs中性检验推测种群的历史动态,当中性检验结果为负值,并且在统计学上有显著性差异,可能预示着种群经历过一个扩张的历史[22-23]。本研究结果显示,湖鲚群体的Tajima’sD和Fu’sFs中性检测结果均为负值,统计检验均有显著或极显著差异,这说明湖鲚种群进化过程中偏离了中性选择,经历了种群扩张,进而形成了高h和低π的遗传多样性模式。
3.2 湖鲚种群遗传结构分析
遗传分化指数Fst是反映群体遗传组成的重要参数,Fst值越大则表示遗传分化程度越大。Freeland等[24]提出,以Fst值0.05、0.15和0.25为临界值指示遗传分化程度。本研究中,未分组的AMOVA结果显示,湖鲚总的Fst值为0.08553,且具有极显著差异,表明种群间有显著遗传分化。而分组的AMOVA 结果显示,组间的Fct为0.01084(P>0.05),表明组间无显著遗传差异,组内种群间的Fsc值为0.08035,种群内的Fst值为0.09032,且都具有极显著差异,表明湖鲚组内种群间及种群内均有显著遗传分化。两两群体间的Fst统计结果表明,滆湖群体与其他群体间的Fst值均大于0.15,且统计检验具有极显著性差异。除滆湖群体外,其他5 个群体间Fst值为-0.01588~0.01533,且统计检验没有显著性差异。可以得出,滆湖群体与其他群体间具有显著遗传分化,而其他群体间无遗传分化。同时,Cyt b 基因单倍型的分子系统进化树显示,6 个群体的单倍型没有聚成各自的分支,单倍型间的聚类关系没有与群体地理位置形成对应关系,仅有滆湖的部分单倍型形成了独立的进化分支,系统进化树支持群体间的遗传关系。
太湖和滆湖位于长江以南同属长江流域;高邮湖、白马湖、洪泽湖和骆马湖位于长江以北均属淮河流域。滆湖与太湖群体间有极显著的遗传分化,而太湖群体与淮河流域的4 个群体间没有显著遗传差异,表明湖鲚群体的遗传结构与群体所处水系及地理位置的远近没有相关性。而Ma 等[21]对中国4 个水系的10 个湖鲚种群的遗传多样性和遗传结果进行分析,发现地理距离是导致的种群遗传分化的主要因素。通常情况下,同一水系内群体间有广泛的基因交流,群体间不会出现遗传分化或遗传分化较小;地理距离越大,基因交流的概率会随之减小,更容易出现遗传分化[25-27]。但地理距离与遗传关系不相关的现象在鱼类中也有较多报道[28-29]。湖鲚种群的遗传结构可能与种群的栖息环境、水利工程及种群历史动态等因素有关。首先,6 个湖泊的生态环境及水文有较大差异,不利于鱼类之间的基因交流。其次,由于兴建了大量的水利工程,直接阻碍了湖泊及水系间鱼类的基因交流。滆湖与其他湖泊没有直接的水道连接或连接水道受阻,而其他湖泊通过京杭大运河连通,促进了湖泊间鱼类种群的扩散和基因交流[30]。最后,在种群进化过程中,滆湖种群较为稳定,没有发生显著的种群扩张,而其他湖泊种群均经历了显著的种群扩张过程,种群扩张过程促进了种群间的基因扩散,有利于消除种群间的遗传差异。
3.3 湖鲚种质资源管理和利用建议
目前,湖鲚已成为江苏省淡水湖泊鱼类群落中的优势种群,也是渔业生产中的重要捕捞对象。同时也面临资源过度开发利用、种群组成趋于低龄化等问题。本研究结果显示,江苏省6 个湖鲚种群遗传多样性较为丰富,均具有高单倍型多态性和低核苷酸多态性的特征。滆湖种群的遗传多样性最丰富,拥有的私有单倍型最多,且与其他种群之间具有显著的遗传分化,应将滆湖群体单独作为一个进化显著单元进行管理和利用,其他5 个群体作为一个整体进行管理和利用。一方面,需要严格控制湖泊渔业资源的捕捞强度,禁止对湖鲚资源过度开发利用,可通过增大渔具网目尺寸和提高开捕年龄来改善湖鲚种群年龄组成。另一方面,可通过适量增殖放流肉食性的翘嘴鲌、鳜等鱼类,控制湖鲚种群数量,优化湖泊鱼类群落结构,促进湖泊渔业资源可持续健康发展。
4 结论
总之,江苏省6 个湖泊湖鲚群体的遗传多样性比较丰富,且呈现出高单倍型多样性和低和低核苷酸多样性的特点,单倍型多样性和核苷酸多样性分别为0.717 ± 0.060~0.875 ± 0.044 和0.00180 ± 0.00034~0.00425±0.00033,在进化过程中,滆湖群体较为稳定,没有发生显著的种群扩张,而其他群体则经历了明显的种群扩张过程,这也是导致滆湖群体与其他群体产生遗传分化的因素之一。滆湖群体与其他群体间的Fst值均大于0.15,且统计检验具有极显著性差异(P<0.01),滆湖群体与其他群体间具有极显著的遗传分化,因此应将滆湖群体与其他群体作为不同的进化显著单元进行管理和利用。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
海洋信息技术与应用(2022年1期)2022-06-05
儿童时代·幸福宝宝(2020年9期)2020-09-08
少儿美术(快乐历史地理)(2019年4期)2019-08-27
小学阅读指南·低年级版(2017年2期)2017-03-23
金色少年(奇趣科普)(2016年8期)2016-09-21
癌变·畸变·突变(2014年2期)2014-03-01
