
氨磺灵和秋水仙素诱导橡胶草四倍体效果的比较研究
2021-09-08 07:45杨玉双张继川张立群聂秋海刘实忠
中国农学通报 2021年20期
杨玉双,覃 碧,张继川,张立群,聂秋海,刘实忠
(1中国热带农业科学院橡胶研究所,海口 571101;2北京化工大学材料科学与工程学院,北京 100029;3北京玲珑蒲公英科技发展有限公司,北京 101102)
0 引言
橡胶草(Taraxacum kok-saghyzRodin,TKS)为菊科(Asteraceae)蒲公英属(Taraxacum)大角蒲公英组(Sect.Macrocornuta V.Soest)多年生草本植物(2n=2x=16),原产于哈萨克斯坦,欧洲及中国新疆等地[1]。橡胶草根部可合成天然橡胶,含胶率在2.89%~27.89%之间[2],其天然橡胶的分子结构与巴西三叶橡胶相同,均为顺式-1,4-聚异戊二烯,且物理机械性能与三叶橡胶非常相似,可用于工业化生产[3]。此外,橡胶草具有生长收获周期短(1年即可收获,而三叶橡胶树需要5~8年)[4];种植区域广(在亚热带和温带均可正常生长);适合机械化种植和采收,具有较强的生长繁殖能力等诸多优点,使其成为最具商业开发前景的产胶替代作物之一[5-6]。野生橡胶草的含胶量和生物量偏低,且变异大[7],不能满足橡胶草产业化发展的要求,为此,科学家们在橡胶草种质创新和改良方面做了大量工作,并取得了显著进步。其中,美国俄亥俄州立大学Cornish团队利用育种手段将橡胶草的平均含胶量提高到9%,小规模种植产胶量可达三叶橡胶树水平[8]。但由于橡胶草在大田中的生长活力和竞争力弱,自交不亲和的特性使群体的优良特性无法稳定遗传,使其实际的大田种植产量达不到预期效果,橡胶草的种质改良工作仍然任重道远。
多倍体育种作为植物育种的重要手段之一,因多倍体植株常具有花大、抗逆性强、生长健壮和生物量大等特点,被育种家们广泛应用于各种植物育种中[9-10]。20世纪40—50年代,欧美科学家利用秋水仙素诱导成功获得四倍体橡胶草,并培育出四倍体橡胶草Navashin等品种,含胶量和干根产量均明显提高,但由于年代久远,相关研究信息及材料已无法找到[11-12]。2018年Luo等[13]利用秋水仙素诱导橡胶草种子获得四倍体橡胶草植株,表现出一定的天然橡胶产量提高潜力。到目前为止,国内还没有关于橡胶草多倍体育种的报道。
秋水仙素作为一种常用的化学诱变剂,一直广泛地应用于植物多倍体诱导中[14-15],但其有效诱导率低,且因与植物蛋白的亲和性低,处理浓度大,对植物材料的毒害比较严重,容易造成诱导植株生长异常,甚至死亡[16-17]。Luo等[13]利用秋水仙素诱导橡胶草四倍体,诱变率最高只有4.92%,且秋水仙素的毒性导致橡胶草萌发率和成苗达率不到50%,许多诱导植株生长异常。可见,秋水仙素的毒性对橡胶草生长影响显著,其多倍体诱导效果很不理想。氨磺灵,作为一种除草剂类诱变剂,因其与植物微管蛋白的亲和性高,使用诱变浓度低,且对植物材料伤害较小,价格低廉等特点[18-19],在植物染色体加倍中的应用逐渐增多[20-21],且研究发现,氨磺灵在大蒜[22]、观赏姜花[23]、西瓜[24]等许多植物上的多倍体诱导效果要好于秋水仙素。
本研究以橡胶草幼芽为试验材料,采用浸芽法比较氨磺灵和秋水仙素2 种诱变剂的诱变效果,分析四倍体橡胶草的形态变化,旨在探寻获得适用于橡胶草的四倍体诱变剂和诱变方案,为橡胶草的种质创新和品种选育提供理论依据和技术支持。
1 材料与方法
1.1 材料与试剂
试验材料为橡胶草品系CXCH,2017 年种植于中国热带农业科学院橡胶研究所橡胶草种质资源圃,2018年初收获种子。
2种诱变剂分别选择上海阿拉丁生化科技股份有限公司的氨磺灵(分析标准品)和上海麦克林生化科技有限公司的秋水仙素粉末(含量≥99%)。
试验在中国热带农业科学院橡胶研究所橡胶草种质资源圃,于2018年9月—2020年5月份进行。
1.2 方法
1.2.1 多倍体诱导 采用浸芽法进行诱变处理。设置秋水仙素浓度为0.05%、0.1%、0.2%;氨磺灵浓度为0.01 mmol/L(约0.0003%),0.02 mmol/L(约0.0007%),0.04 mmol/L( 约0.0014% )和0.08 mmol/L( 约0.0028%),对照采用无菌水浸泡30 min。橡胶草CXCH种子用1%双氧水浸泡、杀菌1 h,之后用水冲洗干净,放入带滤纸的培养皿中萌发,当种子萌发,胚根长至5~10 mm左右时,进行诱变处理。处理时间分别为30 min,1 h,2 h,4 h 和8 h。每组处理100 个幼芽,3个重复。处理结束后用无菌水冲洗3 遍,然后移栽至培养土中正常培育,待植株生长至5~7片叶时,进行倍性筛选。
1.2.2 多倍体鉴定 流式细胞仪倍性分析:流式细胞倍性分析参考康向阳等[25]的方法,取待测植株新鲜叶片0.1~0.15 g 放入直径6 cm 的玻璃培养皿中,滴加500 μL 细胞核提取液(CyStain UV Precise P05-5002,Partec 德国),用锋利刀片一次性快速切碎叶片至糊状,用尼龙网滤膜(30 μm)过滤到5.0 ml 离心管中,加入1.6 mL 的DAPI 染色液(Solarbio),混匀染色30 s,将样品移至上样管,用流式细胞仪(Sysmex CyFlow Cube8,德国)检测。
细胞学倍性鉴定:采用橡胶草根尖压片法,参考杨玉双等[26]的方法,切取待测植株约2 mm 长根尖,洗净后用2 mmol/L 8-羟基喹啉溶液室温预处理4 h,卡诺氏固定液(无水乙醇:冰醋酸=3:1)在4℃冰箱中固定24 h,无菌水漂洗5次,60℃条件下1.0 mol/L盐酸解离10 min,再用无菌水漂洗5次,然后用改良卡宝品红染色5 min 后压片。在莱卡DM68 全自动生物光学显微镜(德国)下观察染色体变化情况,拍照记录。
1.2.3 诱变相关参数测定 发芽率和成苗率测定:诱变处理后,将各试验组幼芽分别移栽至相应花盆中正常培育,30天以后计算成苗率。利用流式细胞仪和根尖染色体压片技术筛选、鉴定四倍体植株,统计、计算诱导率和嵌合体率。

1.2.4 形态特征比较 气孔特征比较:参照刘欢等[16]的方法,选取对照和多倍体植株各6株,撕取其植株叶片的下表皮,利用莱卡DM68全自动生物光学显微镜(德国)对气孔的大小、密度等性状进行观察测定,并计算气孔的长度、宽度,统计气孔密度(每mm2中的气孔数目),并拍照记录。
叶形态比较:待处理后植株长出12 片新叶后,选取对照和多倍体植株各6 株,测定其叶长、叶宽(叶片中间最宽部位)、叶厚(叶片中间最厚部位),叶形指数(叶长/叶宽),每指标数据连续测3次。
花和种子形态测量:待处理后植株开花结果后,选取对照和多倍体植株各6 株,测定其花的长度、直径(花基部最宽部位)和花杆的直径(与花的连接处)、种子粒长和粒宽(种子最宽处),每指标数据连续测3次。
1.3 数据分析
本试验数据用使用IBM SPSS Statistics 19 (IBM,New York,USA)软件进行数据处理、方差分析及LSD多重比较等,所有数据用平均值表示。
2 结果与分析
2.1 诱变植株倍性鉴定
利用流式细胞仪对氨磺灵和秋水仙素处理的待测植株进行倍性检测。结果显示,氨磺灵和秋水仙素处理后都能诱导染色体加倍。流式细胞倍性鉴定结果显示(图1),对照橡胶草二倍体(2n=2x=16)的荧光通道值为200(图1A);变异植株,有的在荧光通道值400处出现一个单峰,DNA 含量较对照增加了一倍,为四倍体(图1B);有的在荧光通道值200和400处同时出现峰,为二倍体和四倍体的混倍体,即嵌合体(图1C)。根尖染色体压片观察结果显示(图2),二倍体植株的根尖细胞内染色体数目为2n=2x=16(图2A);诱导后的四倍体植株染色体数目为2n=4x=32(图2B),与流式细胞仪的倍性鉴定结果一致。具体倍性鉴定统计结果见表1。
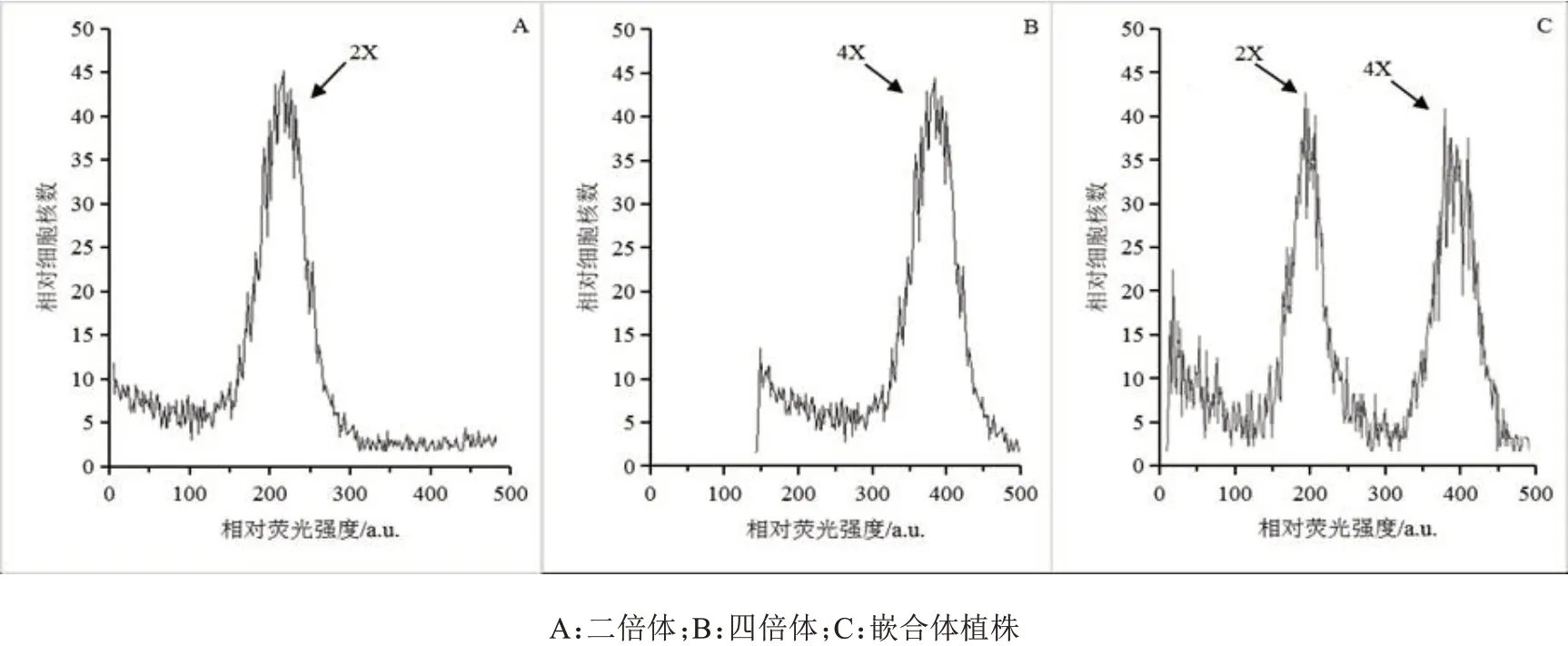
图1 橡胶草流式细胞倍性分析

图2 橡胶草二倍体与四倍体的根尖染色体数比较
2.2 2种诱变剂对橡胶草多倍体诱变效果比较
氨磺灵不同浓度及时间诱变处理均能导致一定程度的诱变效果(表1)。随着处理浓度和处理时间的增加,氨磺灵处理的成苗率呈下降趋势。同一处理浓度下,随着处理时间的增加,变异率逐渐减少,处理浓度为0.04 mmol/L时,不同处理时间的变异率均高于相应其它处理浓度的变异率,其中,0.04 mmol/L 处理30 min的诱变率最高,为14.08%。
秋水仙素的各处理也能产生一定程度的变异。随着秋水仙素处理浓度和处理时间的增加,其成苗率呈下降趋势,其中,0.2%处理浓度对橡胶草的成苗率影响最为明显,当处理时间为30 min 时,成苗率仅为37%,当处理时间≥1 h 时,成苗率为0.00%。在0.05%和0.1%处理浓度下,随着处理时间的增加,其诱变率逐渐降低,其中,0.1%秋水仙素处理30 min 的诱变率最高,为5.56%。0.2%处理浓度下的不同处理时间的变异率均为0.00%。
比较2种诱变剂的诱变效果(表1):氨磺灵的处理浓度范围为0.01~0.08 mmol/L,即0.0003%~0.0028%,明显低于秋水仙素0.05%~0.2%的诱变处理浓度;氨磺灵处理的成苗率(60%~75%)要明显高于秋水仙素处理的成苗率(0~59%),而且其最高变异率14.08%也高于秋水仙素处理的最高变异率5.56%;而氨磺灵处理的嵌合体率为27.27%,明显低于秋水仙素处理的75.00%。综上可知,相对于秋水仙素处理,氨磺灵处理橡胶草的成苗率和变异率高,嵌合体率低,整体诱变效果更好,其中,0.04 mmol/L 氨磺灵处理30 min 的成苗率为71%,变异率14.08%,嵌合体率25%,为最佳诱变条件。

表1 氨磺灵和秋水仙素对橡胶草多倍体的诱导效果

续表1
2.3 四倍体植株与野生型二倍体植株形态特征比较
与野生型二倍体相比,四倍体植株叶片肥厚,叶色浓绿,生长较为缓慢,约60 天后,生长速度恢复正常。氨磺灵和秋水仙素处理后的四倍体植株形态特征变化基本一致。
气孔特征:与野生型二倍体植株相比,四倍体植株的气孔密度显著减少(P<0.01),气孔显著增大。二倍体植株气孔大小为16.87 μm×6.77 μm(长×宽),四倍体植株气孔为20.67 μm×9.07 μm;气孔长和宽分别比二倍体增加22.53%和33.97%(表2)。
叶片形态:四倍体植株叶片与二倍体相比,叶长、叶宽和叶型指数无显著变化,叶片厚度显著增加(表2,图3A)(P<0.01),比二倍体叶片增厚85.71%。
花和种子特征:与二倍体植株相比,四倍体植株的花苞长度无显著变化,而花苞直径和花杆直径显著增大(P<0.01),分别增大了23.51%和36.77%;四倍体植株的种子大小和千粒重均显著大于二倍体植株(P<0.01),其中,四倍体的种子千粒重达到了0.95 g,是二倍体种子千粒重的2.11倍(表2,图3B和3C)。

图3 二倍体与四倍体橡胶草形态特征比较

表2 二倍体与四倍体橡胶草植株的形态特征参数比较
3 讨论
3.1 诱变剂和诱变条件的选择
氨磺灵是二硝基苯胺类除草剂,也是一种有丝分裂抑制剂,与秋水仙素的诱导作用类似,通过抑制纺锤丝的形成,使染色体加倍[27]。本研究中,氨磺灵处理的成苗率在60%~75%之间,而秋水仙素的成苗率仅在0%~59%之间,当处理浓度为0.2%,处理时间≥1 h时,成苗率为0.00%,可见秋水仙素对橡胶草的毒害作用要大于氨磺灵,这与孔素萍等[22]对大蒜多倍体诱导的研究结果大致相似,其原因可能因为秋水仙素与植物微管蛋白的亲和性要比氨磺灵低,秋水仙素的诱变浓度一般要高于氨磺灵诱变浓度,且秋水仙素毒性要高于氨磺灵。诱变率和嵌合体率是判断诱变剂诱变效果的重要指标。Sakhanokho等[23]诱导观赏姜花多倍体结果显示,氨磺乐灵诱变的效果(最高诱变率15%)优于秋水仙素(最高诱变率13%)。
阎志红等[24]在西瓜四倍体诱导时,氨磺灵的四倍体诱导率高于秋水仙素,分别达33.3%和13.3%;孔素萍等[22]对大蒜多倍体诱导的研究结果显示,氨磺灵的诱变率(20.25%)要高与秋水仙素(12.25%),其嵌合体率(2.45%)要低于秋水仙素(3.10%);而储丽红等[17]在安祖花多倍体诱导研究结果显示,秋水仙素的四倍体诱导率高于氨磺灵,分别为45.10%和42.22%。可见,2 种诱变剂对不同植物的诱变效果存在差异。本研究中,氨磺灵的最高诱变率在14.08%,嵌合体率为27.27%;而秋水仙素的最高诱变率为5.56%,嵌合体率为75.00%,这与Luo 等[13]获得的秋水仙素诱导四倍体橡胶草的最高诱变率(4.92%)一致。综上所述,相对于秋水仙素,氨磺灵因具有高成苗率、高变异率和低嵌合体,而更适用于橡胶草多倍体诱变。其中,氨磺灵处理0.04 mmol/L处理30 min的诱变率最高,为14.08%,是橡胶草四倍体诱变的最佳处理条件。
3.2 倍性的鉴定
倍性鉴定是倍性育种的重要环节,简单、快速、有效的鉴定倍性能大大减少工作量,实现早期筛选与应用,加快育种进程。染色体计数法是鉴定倍性最直观、最可靠的方法之一,但制片难度较大,工作量大,而且嵌合体植株很难检出,故不宜用来进行大批量的植株倍性快速鉴定[28],由于橡胶草幼苗根尖细小,取材困难,用染色体计数法确定植株倍性就更难实施。流式细胞仪分析法根据测定细胞的DNA 含量比较染色体倍性,是近几年发展起来的较为快捷、准确鉴定多倍体植株的方法,已在许多植物倍性鉴定中成功应用[29],但由于不同植物的细胞结构不同及次级代谢产物的特殊性和复杂性,流式细胞仪鉴定植株倍性的适用性也不同[30]。本研究显示,利用流式细胞仪可在植株幼苗阶段可快速鉴定出多倍体、嵌合体,且流式细胞仪鉴定结果与传统的染色体数观察鉴定结果一致,证明该方法用于橡胶草植株倍性分析是有效的。此外,学者们在许多植物多倍体诱导研究中表明,四倍体植株常表现为根、茎、叶、花、果实等器官明显增大,叶下表皮气孔增大、密度减少等特点[29-30]。本研究显示,橡胶草四倍体植株,较野生型二倍体植株,叶片显著增厚(P<0.01),花苞和种子显著增大(P<0.01),叶下表皮气孔显著增大、密度显著减少(P<0.01)。这与Luo等[13]获得的四倍体橡胶草形态变化一致,进一步说明这些形态特征可作为橡胶草多倍体初步快速鉴定和育种的有效方法。
4 结论
氨磺灵和秋水仙素2种诱变剂均能诱导出四倍体橡胶草。氨磺灵诱变效果和成苗率均高于秋水仙素,更适合用于橡胶草多倍体诱变,氨磺灵0.04 mmol/L处理30 min 的诱变率最高,为14.08%,是橡胶草四倍体诱变的最佳处理条件。利用流式细胞仪可在植株幼苗阶段快速鉴定出多倍体、嵌合体,说明该方法可用于橡胶草快速倍性鉴定。对比形态特征显示,橡胶草四倍体植株叶片增厚,花苞和种子增大,叶下表皮气孔增大、密度显著减少,与二倍体植株差异显著,可作为多倍体初步筛选标准。本研究初步探明了2种诱变剂对橡胶草染色体加倍的诱导效果,为今后橡胶草多倍体育种提供理论依据和技术支持。
猜你喜欢
园艺与种苗(2023年1期)2023-03-17
园艺与种苗(2023年1期)2023-03-17
科学与社会(2021年3期)2021-12-02
今日农业(2020年14期)2020-12-14
诊断学(理论与实践)(2020年6期)2020-12-09
生殖医学杂志(2019年11期)2019-02-15
广东农业科学(2018年5期)2018-08-08
发明与创新(2017年22期)2017-06-10
临床与实验病理学杂志(2017年3期)2017-03-07
方圆(2016年15期)2016-09-14
