
雷公藤甲素对RA 患者T-bet/GATA3及CXCL10/CXCR3 表达的影响*
2021-09-08 02:29王立新张彤刘秀婵王毅
天津中医药 2021年8期
王立新,张彤,刘秀婵,王毅
(天津市天津医院,天津 300211)
类风湿关节炎(RA)是一种以关节炎、滑膜增生和骨质破坏为特征的慢性自身免疫性疾病,受累滑膜组织中常有大量淋巴细胞和巨噬细胞浸润,其中T 细胞在RA 的发病中处于中心地位[1]。雷公藤是治疗关节肿痛的传统中药,是治疗RA 药物中疗效得到肯定的单味中药。雷公藤及其提取物对RA 的调节机制主要涉及对T 淋巴细胞、B 淋巴细胞、巨噬细胞、树突状细胞以及多种细胞因子表达的影响[2]。雷公藤甲素(TP)是从雷公藤中分离获得的最主要的抗炎、免疫抑制活性成分[3]。研究显示TP 可以降低T 细胞对多种细胞因子如白细胞介素(IL)-2、干扰素-γ(IFN-γ)、IL-4、肿瘤坏死因子-α(TNF-α)、IL-17 等的分泌,以及抑制T 细胞增殖分化从而缓解RA 炎症反应[4]。从Th1 相关的基因调控和趋化因子表达等方面考察TP 的作用机制还值得深入探索和研究。
T-bet 和GATA3 分别是Th1/Th2 细胞的转录因子[5],Th1 细胞分泌的IFN-γ 通过JAK1/JAK2 信号途径激活、诱导T-bet 基因表达,而T-bet 又可以诱导Th1 分化继续产生IFN-γ,形成正反馈环路。与T-bet 作用相反,GATA3 促进Th0 向Th2 细胞分化,这两个基因参与Th1/Th2 细胞分化平衡[6]。此外,IFN-γ 诱导的趋化因子(CXCL)10 通过CXCL 受体3(CXCR3)介导活化T 细胞,尤其是Th1 细胞迁移[7-8]。在RA 患者的血清、滑液(SF)和滑膜组织(ST)中,CXCL10 表达水平升高,因此CXCL10 升高也成为宿主发生免疫反应,尤其是Th1 细胞主导的免疫反应发生的标志[9]。与CXCL9 和CXCL11 相比,CXCL10与CXCR3 亲和力更高,CXCL10 不仅对T 细胞的发育、迁移、黏附起着重要作用,还能活化单核细胞及自然杀伤细胞参与自身免疫疾病的发生过程[10]。以CXCL10/CXCR3 为靶标调节免疫细胞的活化和定位用于RA 治疗的研究也取得一定进展[11]。
TP 对IFN-γ 表达具有明确的抑制作用,但TP 对RA 患者的免疫调节是否也会影响T-bet/GATA3 和CXCL10/CXCR3 的表达值得探索。本研究欲通过CD3 抗体诱导活化RA 患者T 细胞,在分析TP 对Th1、Th2 细胞分化及相关细胞因子分泌影响基础上,进一步考察TP 对T-bet/GATA3 以及CXCL10/CXCR3表达的影响,旨在为TP 治疗RA 的作用机制进行更深入的补充。
1 材料与方法
1.1 材料、主要试剂及仪器
1.1.1 临床资料 本研究经天津市天津医院伦理委员会审核通过(2019 医伦审103 号),所有研究对象均签署知情同意书。选取2019 年11 月—2020 年1 月在天津市天津医院感染风湿免疫科就诊的21 例RA 患者作为研究对象,其中女性16 例,男性5 例。患者年龄为17~89 岁,平均年龄(56.15±17.66)岁,诊断参照美国风湿病学会(ACR)/欧洲抗风湿病联盟(EULAR)2009 年RA 诊断标准。
1.1.2 试剂和仪器 GT-T551 无血清培养基购自Takara 公司,重组人IL-2(rhIL-2)购自北京双鹭药业股份有限公司,TP 购自成都曼思特生物科技有限公司,人外周血淋巴细胞分离液购自天津市灏洋生物制品科技有限公司,CCK-8 试剂盒购自东仁化学科技(上海)有限公司,cDNA 合成试剂盒、微量样本总RNA 提取试剂盒、荧光定量检测试剂盒均购自天根生化科技有限公司。anti-human CD3 抗体、异硫氰酸荧光素(FITC)anti-human CD4 抗体、PE antihuman IL-4、APC anti-human IFN-γ、PerCP-Cy5.5 anti-human CXCR3、细胞固定剂、破膜剂、1 000×布雷菲德菌素A(BFA)均购自Biolegend 公司。L929成纤维细胞由中国医学科学院生物医学工程研究所提供,Luminex 细胞因子检测试剂由国典医药提供,引物合成由上海生工提供。倒置显微镜型号为Olympus CKX41SF,流式细胞仪型号为美国BD Calibur,聚合酶链反应(PCR)扩增仪型号为美国伯乐CFX-96,Luminex 检测仪器型号为Luminex 200,酶标仪型号为奥地利Tecan Sunrise。
1.2 实验方法
1.2.1 考察TP 细胞毒性 本研究选取体外细胞毒性分析模型细胞L929 评价TP 的细胞毒性。取对数生长期的L929 细胞,以1×104个/mL 密度接种于96 孔板,实验分为空白组(单纯培养基)、对照组(TP浓度为0 nmol/L)、实验组(TP 浓度分别为2.5、5、10、25 和50 nmol/L),每组5 个复孔,分别培养48、72 h。去掉培养上清,用磷酸缓冲盐溶液(PBS)洗涤细胞两遍,每孔加入含有10%的CCK-8 新鲜培养基100 μL,37 ℃、5%二氧化碳(CO2)孵育2 h,采用酶标仪检测450 nm 的吸光度(A)值。细胞存活率=[(As-Ab)/(Ac-Ab)]×100%,其中As 为实验孔(细胞+TP),Ac 为对照孔(细胞),Ab 为空白孔(单纯培养基)。
1.2.2 TP 对RA 患者单个核细胞(PBMC)增殖活性的影响 收集10 mL 肝素抗凝全血,无菌条件下与PBS 缓冲液1∶1 混合,将细胞悬液缓慢转移至等体积的淋巴细胞分离液上层,室温条件下490×g 离心30 min。吸取中间白膜层单个核细胞,用10 倍体积PBS 洗涤(490×g,离心10 min)两遍。用含1%双抗、1 000 U rhIL-2 的GT-T551 培养基调整细胞浓度为1×106个/mL,每孔200μL 接种于CD3 单抗(0.5μg/mL)包被的96 孔板中,分为对照组(TP 浓度0 nmol/L)、实验组(TP 浓度分别为2.5、5nmol/L),每组4 个复孔。在37 ℃、5%CO2条件下培养48 h 后,用CCK-8检测细胞增殖情况。
1.2.3 流式细胞仪检测Th、Th1、Th2 细胞亚群变化和CXCR3 表达情况 取培养48 h 的PBMC 细胞,向培养孔内加入终浓度为1×BFA,在37 ℃条件下孵育4 h,用于阻止细胞内因子的释放。离心收集细胞并用PBS 洗涤(350×g 离心5 min)两遍。每组分别加入FITC anti-human CD4 和PerCP-Cy5.5 antihuman CXCR3 抗体各5 μL,室温避光孵育20 min;1 mL PBS 洗涤两遍后,加入0.5 mL 固定液室温避光固定20 min;PBS 洗涤两遍后加入1×破膜剂1 mL重悬细胞,350×g 离心5 min,重复2 次。最后分别加入5 μL 的PE anti-human IL-4 和APC anti-huma IFN-γ 抗体,室温避光孵育20 min,PBS 洗涤后置于流式细胞仪检测。
1.2.4 Luminex 检测细胞因子表达水平 收集PBMC细胞培养上清,分别取50 μL 标准品和待测样本,各加入50 μL 抗体标记微球,铝箔纸封板,(800±50)r/min震荡条件下室温孵育2 h;洗板两次,在所有孔中加入50 μL 的Biotin-检测抗体,室温条件下震荡避光孵育1 h;洗板两次,在所有孔中加PE-链霉亲和素,50 μL/孔,室温条件下震荡避光孵育0.5 h。在所有孔中加100 μL 洗液,室温震荡2 min 使微球悬浮,在Luminex 200 仪器上检测。
1.2.5 实时荧光定量PCR(RT-qPCR)检测基因表达变化 离心收集培养的PBMC 细胞沉淀,PBS 洗涤后按照试剂盒说明书提取样本RNA;用反转录试剂盒37 ℃水浴60 min,将RNA 反转录为cDNA 模板。引物序列检索自Primerbank,使用Pubmed blast分析引物的特异性。具体序列见表1。检测T-bet、GATA3、CXCL10 和CXCR3 基因的表达情况,内参基因为GAPDH。每个样本重复检测3 次。

表1 T-bet、GATA3、CXCL10、CXCR3、GAPDH 引物序列Tab.1 Primer sequences of T-bet,GATA3,CXCL10,CXCR3,GAPDH
1.3 统计学方法 数据采用Graphpad Prism 8.0 软件进行处理,实验结果以均数±标准差(±s)表示,多组间比较采用单因素方差分析,多个实验组与对照组之间的两两比较采用Dunnett 法,P<0.05 为差异有统计学意义。
2 结果
2.1 TP 细胞毒性考察 在TP 浓度<2.5nmol/L,培养48 h 条件下对L929 细胞存活率没有影响(P>0.05),当浓度升高到50 nmol/L 时,细胞存活率为(90.93±2.56)%。随着作用时间延长至72 h,TP 浓度在10 nmol/L 以上时,细胞存活率降低至90%以下,见图1。以上结果表明浓度在25 nmol/L 以内,药物作用时间为48 h 时TP 无显著细胞毒性(P>0.05)。后续实验在此培养时间及安全剂量范围内进行。

图1 TP 对L929 细胞存活率的影响(±s,n=5)Fig.1 Effect of TP on L929 cell survival rate(±s,n=5)
2.2 TP 对RA 患者PBMC 增殖活性的影响 经CD3抗体刺激培养48 h 后,细胞形成显著增殖团,显示出CD3 抗体可以有效刺激PBMC 细胞增殖,见图2。经CCK-8 检测,TP 浓度为2.5 nmol/L 和5 nmol/L时,PBMC 细胞存活率受到显著抑制(P<0.05),两组细胞存活率分别为(82.7±27.57)%、(55.76±21.99)%,并且TP 对PBMC 细胞的增殖抑制作用具有显著的剂量依赖性。见图3。
图2 倒置显微镜观察PBMC 细胞增殖情况(×50)Fig.2 Inverted microscope observation of the cell proliferation of PBMC(×50)

图3 不同实验条件下PBMC 细胞存活率比较(±s,n=4)Fig.3 Comparison of survival rate of PBMC in different experimental condition(±s,n=4)
2.3 PBMC 中Th 细胞、Th1、Th2 细胞亚群比例变化 实验组在TP 浓度为2.5 和5 nmol/L 时细胞亚群比例分别为(47.17±10.34)%和(44.91±11.89)%,均低于对照组Th 细胞亚群比例[(51.34±8.75)%],两组与对照组比较差异具有统计学意义(P<0.01 或P<0.001),且Th1 细胞亚群受抑制程度高于Th 细胞亚群,相反Th2 细胞亚群有升高趋势,但与对照组比较差异无统计学意义(P>0.05)。而Th1 与Th2比值在TP 调节下显著降低(P<0.05 或P<0.001),见图4。以上结果提示,TP 可以抑制Th、Th1 细胞增殖,降低Th1 与Th2 比值来调节Th1 和Th2 亚群比例,减少促炎细胞。本研究结果提示TP 可能会提高Th2 细胞亚群比例,进而抑制Th1 细胞的炎症反应发挥对RA 的免疫调节作用。

图4 TP 对Th 细胞及Th1 和Th2 细胞亚群的影响(±s)Fig.4 Effect of TP on Th,Th1 and Th2 cell subpopulation(±s)
2.4 TP 对CXCR3 受体表达的影响 表达CXCR3受体的细胞亚群在TP 作用下比例显著降低(P<0.05),且TP 对CXCR3+细胞亚群的抑制同样具有剂量依赖性,见图5。CXCR3通过与CXCL9、CXCL10、CXCL11等因子相互作用引导炎症细胞向炎症部位趋化,参与RA 疾病的发生、发展,因此抑制CXCR3 受体表达同样可以减少免疫细胞向炎症部位聚集。
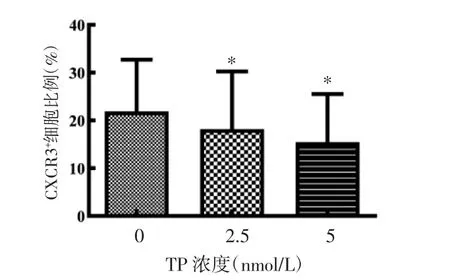
图5 TP 对CXCR3 受体表达的影响(±s)Fig.5 Effect of TP on the expression of CXCR3(±s)
2.5 TP 对PBMC 分泌因子表达的影响 本研究中除TNF-α 外,TP 对其余细胞因子的表达抑制作用均具有剂量依赖性,见图6。在考察的细胞因子中不仅包括促炎因子,也包括抗炎因子。由此可见TP 对细胞因子表达的抑制作用是非特异性的,显示出TP 具有很强的抑制细胞因子表达的作用。对于TNF-α,当TP 浓度为2.5 nmol/L 时可以显著降低TNF-α 表达水平,随着浓度升高为5 nmol/L 时,TNF-α 浓度与对照组比较无统计学差异(P>0.05)。在此浓度下,PBMC 细胞存活率显著降低,因此TNF-α 表达没有变化可能是TP 对免疫细胞活性抑制所致。随着TP浓度升高,在表现出抑制作用的同时,也会对免疫细胞产生一定毒性。

图6 TP 对细胞因子表达的影响(±s)Fig.6 Effect of TP on the expression of cytokines(±s)
2.6 TP 对T-bet/GATA3 及CXCL10/CXCR3 基因表达的影响 与对照组比较,两个浓度的TP 对Tbet 和GATA3 基因表达及T-bet/GATA3 比值均无影响(P>0.05),所以在本研究中并未观察到TP 对这两种基因表达的调节作用。
CXCL10 的基因相对表达量在TP 浓度为5nmol/L时受到了抑制(P<0.05),见图7,这与其蛋白表达水平变化趋势一致,并且CXCL10 蛋白表达受IFN-γ诱导,在TP 作用下IFN-γ 表达显著下降,也可能是诱导CXCL10 表达降低的原因之一。
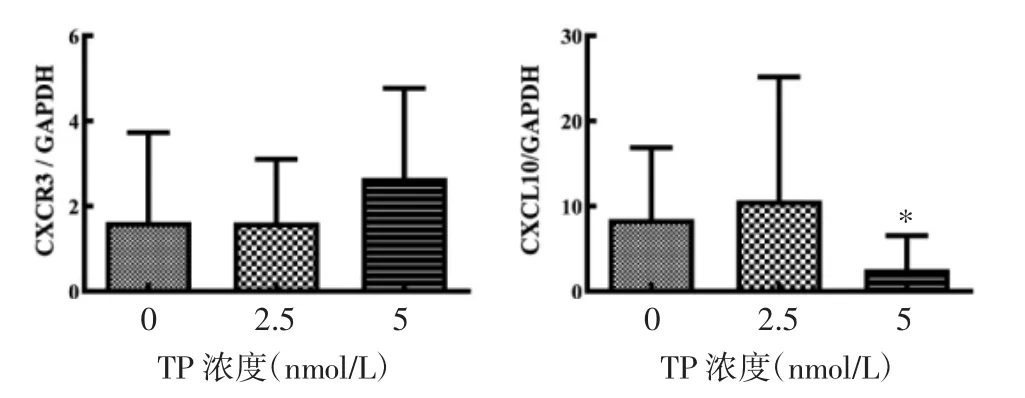
图7 TP 对CXCR3 和CXCL10 基因表达的影响(±s)Fig.7 Effect of TP on the expression of CXCR3,CXCL10(±s)
3 讨论
RA 是一种以滑膜炎症为主的慢性自身免疫性疾病。中药雷公藤具有活血化瘀、去湿止痛的作用,是治疗关节肿痛的传统药物,TP 是其最主要的抗炎与免疫抑制活性成分[12]。本研究通过体外实验,从细胞、细胞因子和基因水平分别考察了TP 对RA 患者PBMC 的调节作用。低浓度TP 对L929 细胞没有毒性,但对活化的PBMC 细胞增殖具有剂量依赖的抑制作用,这与之前报道TP 治疗窗口窄、毒性大相一致[13]。
Th1 和Th2 免疫反应失衡是RA 发病的重要原因[14]。分泌IL-2、TNF-α、IFN-γ 等促炎细胞因子的效应细胞Th1 在TP 作用下百分比降低,然而Th2细胞没有受到显著调节,因此并未观察到TP 抑制Th1 同时会促进Th2 细胞的特异性调节作用。这种现象可能与药物剂量、作用时间以及免疫微环境中细胞因子、趋化因子等的影响相关,有待进一步研究。Th1/Th2 比值显著降低,表明TP 可以减少PBMC中效应细胞比例从而缓解RA 中的炎症反应。
TNF-α 和IL-6 被认为是RA 滑膜细胞因子网络的中心枢纽[15],TNF-α 不仅会诱导Th1 型细胞因子的大量产生,使Th1/Th2 细胞的比例进一步失调,也能诱发关节结构和软骨的破坏,参与滑膜炎症反应[16]。现有研究报道靶向TNF-α 和IL-6 的中和抗体可以有效抑制滑膜炎症反应[17],因此对TNF-α和IL-6 表达的抑制也是TP 参与RA 免疫调节的重要机制。具有免疫调节作用的IL-4 可以诱导巨噬细胞向M2 型转化,促进IL-10 分泌,并可以将细胞毒T 细胞(CTL)转化为分泌IL-4 的无细胞毒性细胞,抑制IFN-γ 的表达[18]。尽管IL-4 和IL-10 作为抗炎因子可以协同抑制Th1 细胞分化,减少相关炎症因子的分泌及炎性细胞浸润[15],但本研究中这两种细胞因子在TP 作用下表达水平同时降低,可能是因为TP 使具有调节作用的如Th2、Treg 等免疫细胞活性受到抑制所致。
CXCR3 与CXCL10 主要参与调节T 细胞、NK 细胞和巨噬细胞的迁移、分化与激活。CXCR3 是体内分泌IFN-γ 的Th1 细胞所必需的[19]。抗CXCR3 抗体AMG487 可以降低RA CIA 小鼠体内Th1、Th17以及Th22 细胞百分比并提高Treg 细胞比例[20]。CXCL10 可与表达在T 细胞表面的CXCR3 受体结合促进其活化并向Th1 细胞方向分化,在炎症细胞向滑膜组织迁移及活化过程中发挥了关键作用[21]。Lee 等[7]研究强调了CXCL10 在RA 发病机制中的重要性,并提供了先前尚未确定的CXCL10 促进关节炎发展机制的细节。因此TP 对CXCL10 和CXCR3 表达的抑制,可以进一步缓解Th1 介导的炎症反应,也丰富了其对RA 免疫调节的作用机制。
与Th1 和Th2 细胞分化相关的T-bet 和GATA3基因[22]表达没有显著变化,一方面可能是经TP 抑制后表达该基因的细胞活性降低,或者由于这两种基因具有复杂多样的生物学功能,如T-bet 不仅是Th1 型免疫的关键调节因子,在T 淋巴细胞、B 淋巴细胞、树突状细胞、NK 细胞效应细胞的建立和维持中也发挥着极其重要的作用[23]。而GATA3 基因也会调控Treg 细胞的分化[24],因此在TP 作用下并未体现出明显变化。
本研究显示TP 对RA 具有显著且广泛的非特异性免疫抑制作用。与体外培养细胞系不同,实验中各组样本间统计数据相对离散,正如李彦等[25]对雷公藤肝毒性的评价所述,造成这种现象的原因与每个个体免疫状态、对药物的敏感性以及基因表达差异有关。因此TP 作为临床用药还需进一步提高其靶向性,降低毒副作用,如何用于RA 临床治疗还有很大研究空间。
猜你喜欢
中国医药科学(2022年5期)2022-05-05
昆明医科大学学报(2021年12期)2021-12-30
现代临床医学(2021年4期)2021-07-31
祝您健康(2020年5期)2020-05-14
科学导报(2020年25期)2020-04-28
植物保护(2019年2期)2019-07-23
包头医学院学报(2019年12期)2019-06-08
河北渔业(2015年1期)2015-01-19
河北渔业(2015年1期)2015-01-19
湖北农业科学(2014年15期)2014-10-28
