
火干扰恢复初期土壤呼吸及其组分与土壤温湿度关系1)
2021-09-07 11:45王梓璇
东北林业大学学报 2021年8期
王梓璇
(内蒙古农业大学,呼和浩特,010018)
周梅 赵鹏武 王鼎 石亮 赵威 刘喜才
(内蒙古赛罕乌拉森林生态系统国家定位研究站(内蒙古农业大学)) (内蒙古农业大学) (内蒙古毕拉河国家级自然保护区)
目前,越来越多研究证明火是影响生态系统碳通量变化的关键因素[1-2]。林火干扰后地表温度增加,加速北方森林冻土的融化,导致土壤碳损失,土壤呼吸增加[3]。吴畏[4]在火干扰对土壤呼吸及其组分的研究中发现,火干扰后短期内,兴安落叶松林土壤呼吸及其组分在不同程度有所增加。Köster et al.[5]研究发现,火烧后的土壤呼吸速率与未火烧样地无显著差异,原因是火干扰后根系呼吸减弱,土壤微生物呼吸增强。Amiro[6]认为森林野火减缓了凋落物分解,从而降低土壤异养呼吸。Hu et al.[7]研究发现,火灾后土壤呼吸下降,主要是土壤自养呼吸减少,而土壤异养呼吸没有明显变化。Taet al.[8]在研究火灾对阿拉斯加北部高地森林的影响时发现,火灾后土壤水分与土壤呼吸均减少。Muoz-Rojas et al.[9]研究表明,火干扰会增加土壤呼吸的温度敏感性,土壤温度、湿度可以很好的反映土壤呼吸变化。在特定立地条件时,利用土壤温度、湿度等因子,建立土壤温湿度与土壤呼吸的回归模型能更好地实现对土壤呼吸速率的动态预测。本研究以大兴安岭冻土区草类-兴安落叶松(Larixgmelinii)林火烧迹地为研究对象,分析火干扰初期重度火烧迹地土壤呼吸速率及其组分变化,探讨土壤呼吸速率与土壤温度、湿度的相关性,旨在为火后森林生态系统的恢复、重建提供参考。
1 研究区概况
依据中国内蒙古森林工业集团有限责任公司毕拉河林业局防火办扑火记录,选取2017年毕拉河“5.2”大火的火烧迹地为试验区。选定火烧前为草类-兴安落叶松林的重度火烧迹地为研究对象,同时选定距离火烧迹地4 km内立地因子相似的未火烧区域为对照区,2个区域分别布设3个30 m×30 m的固定观测样地。
2 研究方法
2.1 土壤呼吸及其组分观测
选择LGR公司便携式温室气体分析仪(UGGA)对2017年火烧迹地土壤呼吸及其组分进行测定,测定时间为2017—2019年(4—10月冻融期与生长季)。在每块样地内随机布设3组土壤呼吸采样环,共6个(3个土壤呼吸环、3个土壤异养呼吸环),组内2个土壤呼吸环间距不超过2 m,每组土壤呼吸环以等腰三角形的形式随机布设于样地内,且每组土壤呼吸环间距大于10 m。土壤呼吸环为直径20.0 cm、高13.5 cm的聚氯乙烯(PVC)环。将采样环纵向插入土中,露出地表的2 cm与气室连接,待PVC环与土壤接触紧密后进行土壤呼吸测定。
采用壕沟法测定土壤异养呼吸。在已布置好的土壤呼吸环附近2 m内选择1块直径为1 m的圆形区域,在其周围用铁锹挖30 cm宽的壕沟,垂直挖深到无根系生长深度(0.4~0.6 m),切断根后(不移走)铺入硬质塑料膜,以阻止根向圆形原状土内生长。拔除原状土区域地上植被并定期清理,以去除新生草本植被的自养呼吸对原状土区域内土壤异养呼吸测定的干扰。断根处理40 d后开始测定土壤呼吸。为确保断根区域根系死亡,在布设壕沟同时挖取3处圆形原状土区域,在开始观测土壤呼吸前将3处原状土区域内根系全部取出,经判别后发现根系死亡率达95%以上。有研究表明,因挖沟造成的死根及其分解对土壤累积呼吸的影响较小,约为3%~5%[10-11],故在布设壕沟40 d后开始测定。
RA=RS-RH;
(1)
CRA=[(RS-RH)/RS]×100%。
(2)
式中:RS为土壤呼吸速率;RH为土壤异养呼吸速率;RA为土壤自养呼吸速率;CRA为土壤自养呼吸贡献率。
2.2 土壤水热因子观测
在观测土壤呼吸的同时,使用手持长杆电子温度探针(JM-222)测定土壤呼吸环0~10 cm内土壤深度为10 cm处的温度(T10);使用时域反射仪(TDR300)测定每个土壤呼吸环附近0~10 cm内土壤深度为10 cm处的湿度(W10)。对土壤呼吸及其组分与土壤温度、湿度之间的关系进行回归模拟。
RS=αeβT。
(3)
式中:RS为土壤呼吸速率;T为土壤温度;α为0 ℃时的呼吸速率;β为温度反应系数。
Q10=e10β。
(4)
林火干扰不仅会增加土壤对大气的碳排放,还会影响森林土壤碳循环[13]。森林火灾影响季节性冻融层的排水,导致局部水、气的重新分布,迫使多年冻土中大量有机碳以CO2等气体形式释放[4,14]。大量研究表明,火烧迹地土壤CO2释放量高于未火烧区[15-17];也有研究表明,火灾后CO2的排放量较未火烧相比减少了40%~60%[18-19]。Grangedet et al.[20]、Plaza-lvarez et al.[21]的研究均表明,林火干扰恢复初期,土壤CO2释放量变化不明显。本研究发现,林火干扰后土壤呼吸速率先减少后增加。火烧当年的火烧样地土壤呼吸速率低于未火烧样地,但无明显变化。随着恢复时间的推移,火烧样地土壤呼吸速率逐渐高于未火烧样地,3 a的土壤呼吸速率增加总量为13.14%。虽然林火干扰恢复初期,由于根系死亡。土壤自养呼吸短时间内被抑制,但该效应会被林火干扰后土壤异养呼吸的增加所掩盖[22]。Hicke et al.[23]研究发现,由于较低的净初级生产力,土壤异养呼吸将会在林火干扰2 a后开始降低,林火干扰5 a后将低于未火烧前的水平。一般来说,火烧会降低土壤呼吸速率,降低程度受森林火灾的强度、火烧持续时间的影响[24],而本研究区域林火干扰后,土壤呼吸速率增大。为了进一步探究林火干扰对土壤呼吸的影响,对其组分进行分析,发现林火干扰会增加土壤异养呼吸速率,2017年增加了16.18%,2018年增加了54.71%,2019年增加了40.96%。原因是林火干扰后的恢复初期,大量灰分物质的分解为微生物活动提供了良好的养分基础,土壤微生物的数量、种类不同程度增加,增强了土壤微生物的呼吸作用[25-26]。
如表3所示,2017—2019年观测期内火烧样地T10年均值较对照样地相比分别增加13.91%(2017年)、13.13%(2018年)、11.36%(2019年),T10年均值升高比例由高到低依次为:2017年、2018年、2019年。综合3 a观测数据显示,火烧样地T10年均值较对照样地相比显著增加了12.12%(P<0.05)。火烧样地T10升高比例随着恢复时间的增加而降低,原因是火干扰后,随着地上植被的恢复,植被覆盖度略有上升,起到了良好的遮阴效果,从而降低了土壤温度。
RS=aW2+bW+c。
(5)
式中:a、b、c为拟合常数;W为土壤湿度。
ln(RS)=f+h×T+i×W+g×T×W。
(6)
式中:ln(RS)为土壤呼吸速率自然对数转换;f、h、i、g为拟合常数[12]。
综合3 a观测数据可知,火干扰会导致RS发生变化。火干扰当年RS降低,随着恢复时间的推移,火烧样地RS逐渐高于对照样地。火干扰后恢复期的前3年,火烧样地较对照样地相比,RS增加了13.14%;RH增加了38.38%;RA降低了34.09%,火烧样地与对照样地的RH、RA均差异显著(P<0.05)。
2.3 数据处理
采用Excel 2016对土壤呼吸及其组分、土壤温湿度进行初步计算。采用SPSS 20.0软件对土壤呼吸及其组分、土壤温湿度(T10、W10)进行单因素方差分析,并对其进行回归分析(式3,式5)。将土壤呼吸及其组分进行自然对数转换,对T10、W10及其交互作用进行多元逐步回归分析(式6)[12]。
对土壤呼吸及其组分与T10、W10进行回归分析(表4)。结果显示,对照样地、火烧样地的土壤呼吸及其组分均随T10的升高而增强,T10与土壤呼吸及其组份呈极显著正相关(R2>0.28,P<0.01)。对照样地与火烧样地RS、RH与T10拟合度大于RA与T10拟合度;火烧样地RS、RH与T10的拟合度大于对照样地,RA与T10的拟合度小于对照样地。对照样地与火烧样地土壤呼吸及其组分与W10呈显著正相关关系,但判定系数较低(R2<0.25,P<0.05)。火烧样地RS、RH与W10的拟合度小于对照样地,RA与W10的拟合度大于对照样地。对照样地与火烧样地土壤温度与土壤湿度的交互作用(W10×T10)与土壤呼吸及其组分均呈极显著正相关(R2<0.48,P<0.01)。火烧样地RS、RA与W10×T10的拟合度低于对照样地,可见,林火干扰降低了RS、RH与W10;RA与T10;RS、RA与W10×T10的拟合度,增加了RS、RH与T10;RA与W10;RH与W10×T10的拟合度。
3 结果与分析
3.1 火干扰恢复初期土壤呼吸速率及其组分差异
火干扰恢复初期不同观测年份土壤呼吸及其组分均值与变化范围(见表1)。火灾发生当年(2017年)土壤呼吸及其组分最大值均出现在对照样地;火灾发生第2年、第3年土壤呼吸速率(RS)与土壤异养呼吸速率(RH)最大值均出现在火烧样地,土壤自养呼吸速率(RA)最大值均出现在对照样地。
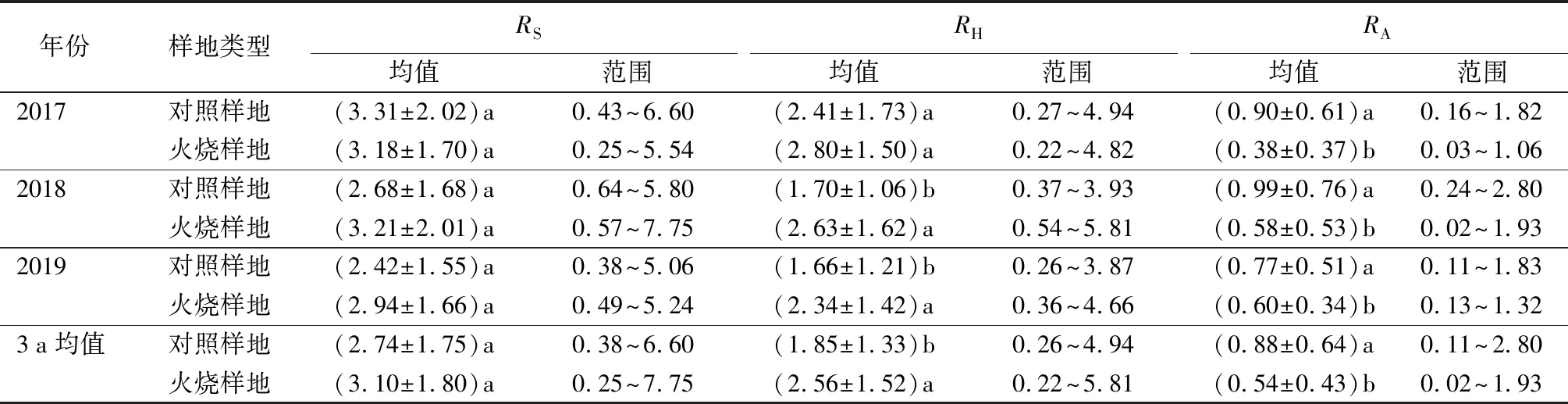
表1 火干扰恢复初期不同观测年份土壤呼吸及其组分均值与变化范围 μmol·m-2·s-1
火灾发生当年,火烧样地RS年均值小于对照样地,比对照下降3.93%,无显著性差异(P>0.05)。火灾发生后第2年、第3年,RS年均值均表现为火烧样地大于对照样地,RS分别增加了19.78%、21.49%。3 a观测期内RH年均值均表现为火烧样地大于对照样地,其中2018年、2019年火烧样地RH年均值显著大于对照样地(P<0.05)。3 a观测期内火烧样地较对照样地相比,RH分别增加了16.18%(2017年)、54.71%(2018年)、40.96%(2019年)。3 a观测期内RA年均值均表现为火烧样地显著小于对照样地(P<0.05)。火烧样地较对照样地相比,3 a观测期内RA分别降低了57.78%(2017年)、41.41%(2018年)、22.08%(2019年),随着恢复时间的推移,火烧样地RA逐渐升高。
土壤呼吸速率对温度敏感度为温度敏感系数(Q10)。对照样地RS、RH、RA的Q10值分别为2.61、2.87、2.22;火烧样地RS、RH、RA的Q10值分别为2.38、2.47、2.03。火烧样地与对照样地土壤呼吸及其组分的Q10值由大到小排列顺序一致,依次为:RH、RS、RA,火烧样地Q10值均小于对照样地。可见,火干扰降低了土壤呼吸及其组分对T10的敏感性。
对比以作平行线求交点法所制梯形槽参数与实际梯形槽参数(如表3),可知以作平行线求交点法所制梯形槽参数与理想梯形槽参数完全相符,故为保证梯形槽电机槽满率、绕线工艺性及电机性能,建议采用以Ansoft计算所得理想梯形槽参数为基础,利用作平行线求交点法制梯形槽电枢冲片。
观测期内土壤自养呼吸贡献率(CRA)月均值3 a观测数据显示(表2),火烧样地CRA均小于对照样地。对照样地CRA月均值最大值均出现在9月。火灾发生当年,火烧样地CRA月均值最大值出现在8月;火灾发生第2年、第3年,CRA月均值最大值均出现在9月。火烧样地较对照样地相比,3 a观测期CRA分别降低了62.24%(2017年)、51.84%(2018年)、33.49%(2019年)。2017—2019年该区域健康林分(对照样地)CRA为33.27%,火烧样地CRA为17.68%,火烧样地CRA较对照样地相比显著下降了46.86%(P<0.05)。可见,火干扰降低了CRA,随着恢复时间的推移,CRA呈上升趋势。土壤异养呼吸贡献率(CRH)与CRA规律呈相反趋势。
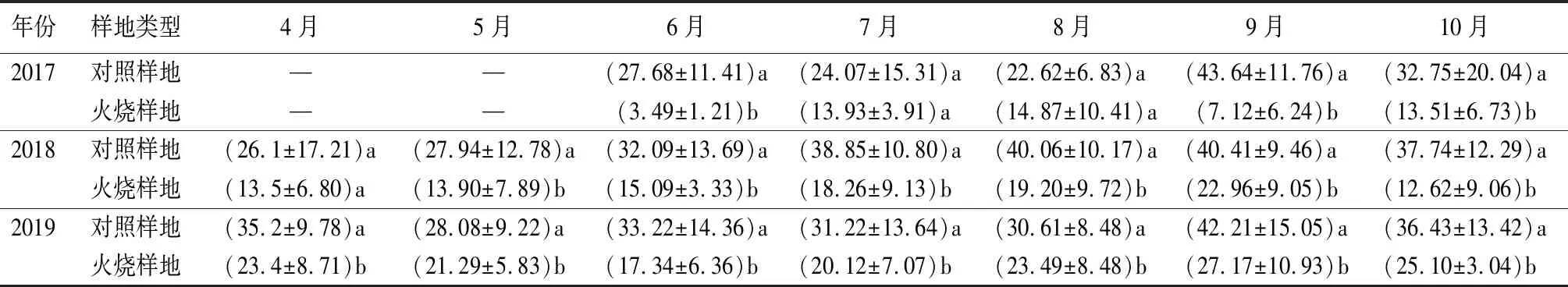
表2 火干扰恢复初期土壤自养呼吸贡献率的月均值
3.2 火干扰恢复初期土壤温湿度变化
低速冲击过程中的能量转化主要是冲头的动能(KE)转化为波纹夹芯结构的内能(IE),而波纹夹芯结构的内能主要由塑性应变能(PE)、弹性应变能(EE)和其他损耗(DE)组成,将其他损耗的能量统称为伪应变能(DE)。在反弹的过程中,波纹夹芯结构的弹性应变能又转化为冲头的动能,因此最终的能量转化主要是冲头的部分动能转化为波纹夹芯结构的内能(主要是塑性应变能)。

表3 火干扰恢复初期不同观测年份土壤温湿度
3 a观测期内,火烧样地W10较对照样地相比分别增加13.97%(2017年)、9.94%(2018年)、10.92%(2019年),且2019年火烧样地W10年均值显著高于对照样地(P<0.05)。W10年均值升高比例由高到低依次为:2018年、2017年、2019年。综合3 a观测数据显示,火烧样地W10较对照样地相比显著升高11.12%(P<0.05)。原因是火干扰后高层植被郁闭度下降,植被截流降水减少,导致雨水下渗。
3.3 火干扰恢复初期土壤呼吸及其组分与土壤温湿度关系
2.3.2 病死率 3项研究[3,6,16]报道了病死率,各研究间无统计学异质性(P=0.12,I2=48.1%),采用固定效应模型进行分析,详见图3。Meta分析结果显示,两组患者病死率比较差异无统计学意义[RR=1.08,95%CI(0.83,1.40),P=0.56]。
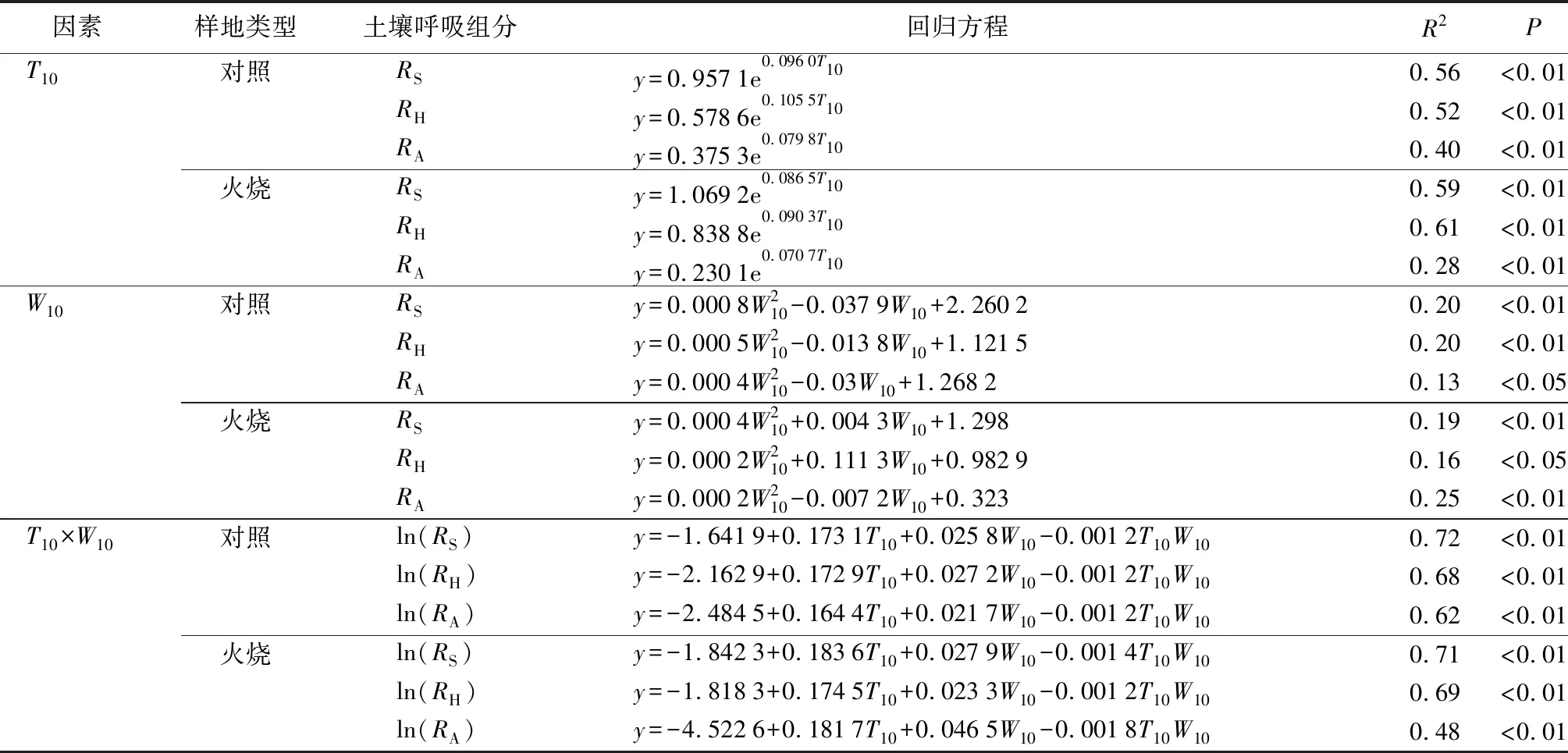
表4 火干扰恢复初期土壤呼吸及其组分与土壤温度和土壤湿度回归关系
邓小平社会管理改革思想是邓小平理论的重要组成部分。邓小平在吸收马克思主义社会管理思想素养的基础上,依据当代国际环境,结合中国改革开放的具体实际,认为社会管理改革迫在眉睫。分析和总结邓小平重视社会管理改革的原因,对当今坚持和推进社会管理改革具有十分重要的意义。
4 讨论
4.1 火干扰对土壤呼吸及其组分的影响
式中:Q10为土壤温度敏感系数,代表温度变化10 ℃时土壤呼吸的相对变化倍数;β为温度反应系数。
本小节将采用仿真实验对本文所提算法的杂波抑制性能进行验证,并与原有SAR-STAP算法进行比较,实验参数如表2所示。图6给出了两种算法的自适应天线方向图,图中所有曲线均为500次Monte-Carlo实验的平均结果。从图6(a)中可以看出,当α=2时(即高斯杂波背景条件下)两种算法性能基本相同,两条曲线几乎重合,都能够在目标多普勒位置形成高增益,同时在杂波多普勒中心位置形成凹口。而从图6(b)、图6(c)、图6(d)可以看出,随着α的减小,原有算法自适应方向图在杂波中心位置凹口深度明显变浅,杂波抑制性能显著下降,与之相比,无论是在杂波还是信号方向新算法都能获得较为理想的性能。
胡同欣等[27]的研究发现,林火干扰后土壤自养呼吸速率显著低于未火烧样地,本研究结果与其相同。林火干扰会对植物根系造成破坏,进而影响植物根系对土壤有机碳的吸收。在短期内,林火干扰对植物活细根生物量的降低是不可逆的,且林火干扰对植物根系的影响持续时间较长[19,28]。孙龙等[29]在中度林火干扰对土壤呼吸组分影响的研究中发现,未火烧样地的土壤自养呼吸贡献率变化范围为17.50%~45.30%,火烧样地土壤自养呼吸贡献率为7.60%~37.80%,火烧样地土壤自养呼吸贡献率显著低于未火烧样地(P<0.05)。本研究结果与其相似,3 a观测期内未火烧样地土壤自养呼吸贡献率为33.27%,火烧样地为17.68%,火烧样地土壤自养呼吸贡献率显著小于未火烧样地(P<0.05)。火烧迹地土壤自养呼吸的显著降低是造成土壤自养呼吸贡献率降低的主要原因。
晏阳初是平民教育者,他曾在香港、美国等地学习政治经济,通过中西文化对比,他找到了中国“事事不如人”的根本原因——中国的人,具体的说是中国的平民,其中农民占绝大多数。由此,他提出了 “民族再造”的思想,主张从中国农民的“愚、贫、弱、私”四大弊病入手,进行农村的政治、教育、经济、自卫、卫生和礼俗六个方面的建设。1926年,晏阳初在河北定县开始乡村平民教育实验,他为农村建设开出的药方是“除文盲,做新民”,他主张“博士下乡”,用文艺教育来治疗愚昧,用生计教育来治疗贫穷,用卫生教育来治疗体弱,用公民教育来治疗自私。
4.2 火干扰对土壤温湿度的影响
植被状况是决定土壤温度的重要因素。植被可以调节区域气候,从而影响林下生物与非生物[30-31]。火灾破坏了森林生态系统原有的植被结构,使土壤地表层接受更多的太阳辐射,冻土区域土壤温度升高[32-33]。大量研究表明,林火干扰后冻土区土壤温度上升,而土壤湿度变化规律尚不确定;多数研究表明,林火干扰后冻土区土壤湿度增加[34-36];也有研究发现,火灾后土壤含水量降低[37-38]。林火干扰后冻土区土壤湿度的差异是由于火烧程度、观测区域不同,若观测区域排水状况良好,林火干扰则会降低土壤湿度;若观测区域排水较差,林火干扰则会增加土壤湿度,甚至使林地变为湿地、池塘[39-42]。
分别称取5.00 g样品于烧杯中,加入70%乙醇溶液25 mL,摇匀,超声波水浴浸提30 min,6000 r/min离心15 min,取上清液,滤渣再加70%乙醇溶液同样条件水浴、离心。合并两次上清液,用70%乙醇定容至50 mL。
4.3 土壤呼吸及其组分与土壤温湿度关系
土壤温度被认为是影响土壤各种气体排放的重要因素[43-44]。冻土区土壤呼吸与土壤温度呈正相关。冻土带生态系统呼吸作用对当前气候变暖有一定正反馈效应[25,45]。Song et al.[16-17]研究发现,中国东北冻土区土壤CO2通量与土壤温度呈正相关关系;孙龙等[29]的研究发现,10 cm土壤层的土壤温度与火烧迹地土壤呼吸、土壤异养呼吸均呈极显著正相关关系(P<0.01),与土壤自养呼吸呈显著正相关(P<0.05),火烧样地的土壤呼吸、土壤异养呼吸与不同土壤层温度的正相关性大于未火烧样地,土壤自养呼吸与不同土壤层温度的正相关性小于未火烧样地。本研究中,林火干扰后的恢复初期,土壤呼吸及其组分与10 cm土壤层的温度均呈极显著正相关关系(P<0.01)。林火干扰初期,土壤呼吸速率、土壤异养呼吸速率与土壤温度的相关性一定程度增加,土壤自养呼吸速率与土壤温度的相关性一定程度降低。
温度敏感系数是描述土壤呼吸速率对土壤温度的敏感程度。Q10受植物种类、温度、土壤含水量、营养状况、呼吸基质有效性等的影响[46]。林火干扰后,随着植被、土壤性质、土壤温湿度的变化,Q10也会发生变化。任军等[47]研究中指出,生长季Q10值为1.1~10.0。本试验中,林火干扰恢复初期Q10值也在此范围(1.92~2.87)。林火干扰会降低土壤呼吸Q10值,这与大多数研究相同[17,48]。林火干扰恢复初期,火烧样地土壤呼吸及其组分Q10值均小于未火烧样地。其原因是Q10值随土壤温度升高而减小[49]。
有研究表明,土壤湿度与CO2通量相关性较强[43,50];还有研究表明,极端的条件时,土壤含水率才会成为土壤呼吸的主要调控因素[51-52]。本研究发现,火烧样地与对照样地土壤呼吸及各组分与土壤湿度呈正相关关系,但拟合系数相对较低(R2>0.13;P<0.05)。原因是土壤水分对有机质分解有较强的控制作用(好氧和厌氧)[53]。土壤湿度是影响土壤微生物活性的主要因素[54],林火干扰改变了冻土区土壤环境,进而对土壤呼吸及其组分与土壤湿度的相关性产生影响。
5 结论
通过毕拉河兴安落叶松林重度火烧迹地土壤呼吸定点观测,研究林火干扰恢复初期土壤呼吸及其组分与影响因子关系,得出以下结论:林火发生当年,林火干扰对土壤呼吸与土壤异氧呼吸的影响较小,而随恢复时间的推移,林火干扰对土壤呼吸与土壤异养呼吸有增强作用,对土壤自养呼吸有抑制作用。林火干扰后土壤异养呼吸的增加会抵消土壤自养呼吸的减少。林火干扰后土壤层10 cm处土壤温度、湿度均升高,土壤呼吸速率、土壤异养呼吸速率与T10的拟合度增强,土壤自养呼吸与T10的拟合度降低。林火干扰降低了土壤呼吸及其组分对T10的温度敏感系数。火烧样地与未火烧样地土壤呼吸及其组分与土壤湿度呈显著正相关关系,但拟合系数较低。
猜你喜欢
绿色天府(2022年9期)2022-11-19
中国农业气象(2022年8期)2022-08-24
世界热带农业信息(2022年8期)2022-07-19
贵州畜牧兽医(2022年3期)2022-06-28
林业调查规划(2022年2期)2022-05-07
土壤(2022年1期)2022-03-16
现代计算机(2021年7期)2021-05-12
中国新闻周刊(2020年6期)2020-03-08
江苏农业科学(2017年21期)2017-12-13
