
盛开在世界之巅的高寒植物
2021-09-06 18:37丁文娜星耀武
科学 2021年2期
关键词:生物多样性
丁文娜 星耀武
高寒生物区分布于亚高山森林边界(即高山树线过渡带)以上、永久雪线以下,是陆地生态系统中海拔最高的生物群落,也是唯一具全球分布性质的生物区,其在各大洲高山都有分布,面积约占全球陆地面积的3% [1]。高寒生物区有着严酷的自然环境和独特的动植物群落。虽然就植物多样性的数量而言,高寒生物区植物种类比热带、亚热带雨林低,但因其严酷的自然环境,高寒植物进化出了特殊的抗寒、抗旱、抗紫外线机制,它们是人类重要的基因宝库来源。近年来,全球环境变化的加剧,导致处于中低海拔地区的物种向高海拔地区渗透,压缩了高寒植物的生存范围,物种灭绝的风险增加,相关问题已成为长期以来植物学家和生态学家关注的焦点。
高寒生物区的全球分布
高寒生物区的海拔高度因所处纬度不同而不同,呈现随低纬度向高纬度变化而逐渐降低的规律,并与各地区树线高低甚至坡向密切相关。例如,欧洲阿尔卑斯山的树线平均在1800—2300米,其上的高寒生物区海拔在阿尔卑斯山西部是2200—2900米,在阿尔卑斯山南部则为2300—3000米。相较而言,纬度更低的青藏高原及周边地区的高寒生物区平均海拔在3800米左右,但是其树线(即森林上限)由于各种环境差异也有明显变化,湿润的东喜马拉雅山的树线在3500—3900米,西藏东部工布江达、洛隆的树线则为4400米,昆仑北坡的为3400—3600米,北部的祁连山则降至3200—3600米[2],东南部的横断山在3000—4000米,而位于树线以上的高寒生物区也随着树线的波动而产生变化[3]。不同地区的高寒生物区垂直分布的海拔范围,也因山地坡度的大小以及湿润度而异,通常在半湿润地区的海拔较高,在湿润地区的海拔偏低,在干旱低温的地区更低,在陡峭的山坡,海拔可下延至几百米。
高寒生物区在一众海拔数千米以上的高山上呈岛屿状分布,因此被称为“天岛”(sky island)[4],它们之间由不同海拔梯度的山地和低地所分隔,再加上各山地地区复杂的地形地貌和气候条件,构成了物种之间隔离分布和遗传分化的强烈驱动力;另一方面,已经预适应高寒环境的植物,也通过各种传播途径实现了在不同高山系统之间的迁移扩散和基因交流,形成了具有独特地理分布的高寒植物群。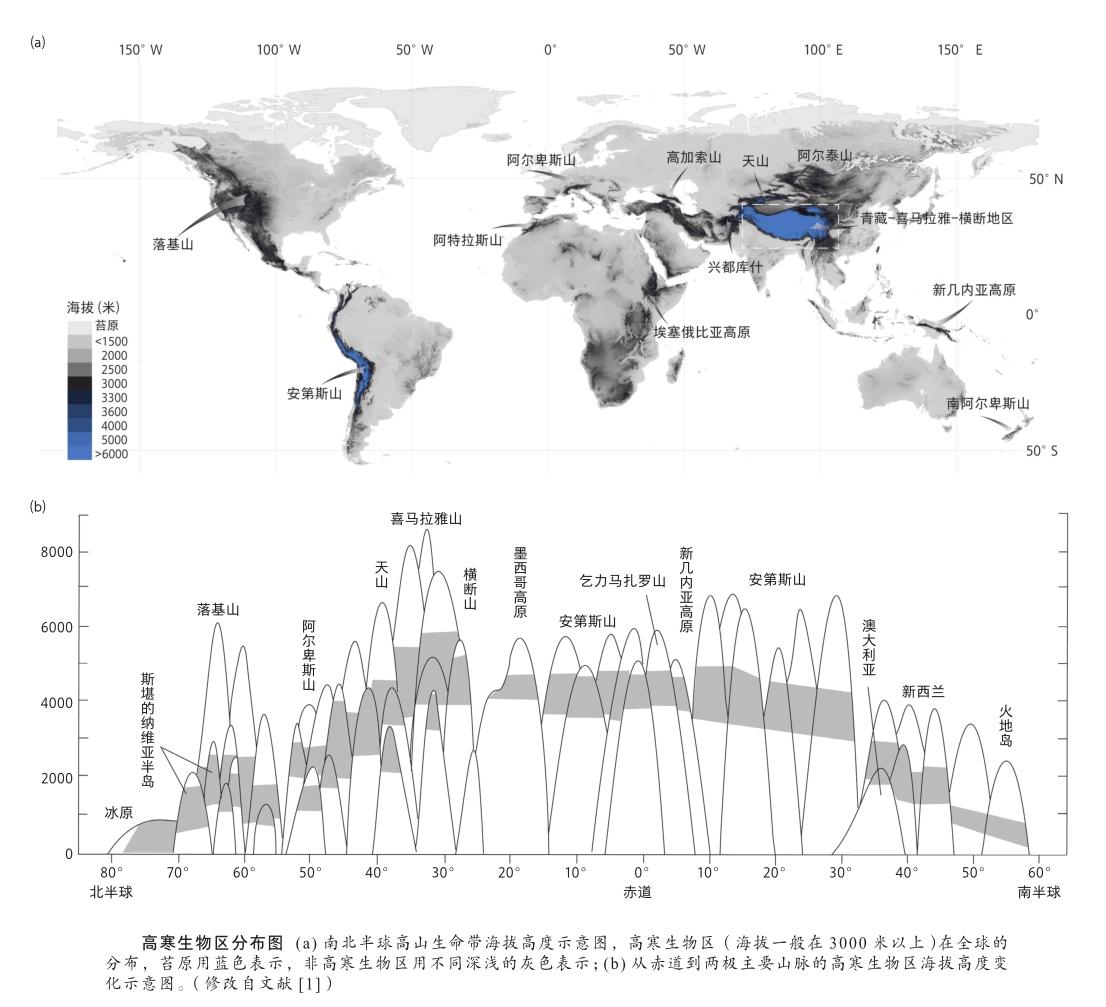
丰富多样的高寒植物
高寒生物区虽然自然环境极端恶劣,却也存在着一个色彩缤纷、生机勃勃的植物世界。就植被类型而言,高寒生物区主要包括高山灌丛、草甸和冰缘带植被等[1],其独特的生物群落特征是长期适应严苛自然环境的结果。由于低温、昼夜温差大、风大、空气稀薄、紫外辐射强烈等环境因素影响,高山植物普遍表现出与高山树线以下非乔木生活型植物显著不同的部分形态和生理属性,其植物区系组成主要是适应冰雪或严寒生境的低矮或匍匐状的灌木、禾草型植物、多年生草本、莲座状植物和各类型垫状植物[1],许多高寒植物的花朵大而艳丽,花部器官复杂多样。据粗略估计,全球高寒生物区有8000—10000种高等植物[1],分属于100科2000余属,占已知高等植物总数的4%左右。其中菊科(Asteraceae)、虎耳草科(Saxifragaceae)、十字花科(Brassicaceae)、毛茛科(Ranunculaceae)、石竹科(Caryophyllaceae)、蔷薇科(Rosaceae)、莎草科(Cyperaceae)、景天科(Crassulaceae)、报春花科(Primulaceae)、龙胆科(Gentianaceae)等科的植物组成了高山主要的植物种类[1]。
高寒生物区以青藏高原及周边山系(包括喜马拉雅山、横断山、天山等)、高加索山、落基山、安第斯山、阿尔卑斯山等地区分布最为集中。其中,青藏高原及周边的横断山和喜马拉雅山,拥有全球海拔最高的高寒生态系统和丰富的高寒植物多样性。这当中以横断山高寒生物多样性尤其丰富,据最近统计,除亚种和变种外,横断山高寒地区的种子植物可达3030种,其中包括12种裸子植物[5]。按照2003年统计的全球高寒地区植物种类计算,仅仅横断山高寒植物就占全球高山植物种类的30% [5],其物种丰富度可与热带安第斯山的帕拉莫草原(有3431种维管植物)相媲美,更是高加索地区(1000种)的3倍、阿尔卑斯 山(650种)的5倍[1],同时也远高于面积更广阔的青藏高原腹地。横断山高寒生物区也是全球生物多样性热点地区中最脆弱的地区之一,可谓是生物多样性热点地区中的热点。
基于生物地理学方法研究特定生物区的起源与演化
目前研究长时间尺度生物群的演化历史,主要基于历史生物地理学方法。它是一门研究生物分布格局及其形成历史的学科,尤其致力于探索环境变化与生物分布间的联系,因此也是一个与其他学科交叉性较强的学科。物种的地理分布同时受到现在和历史因素的影响,其分布格局会由于无法跨越的地理屏障或者不能适应的历史或当今生态条件所限。因此,在区域生态系统中,各生物类群所构成的生物多样性的形成与演变,受到不同尺度地质运动、气候变迁、地貌变化和生态过程的共同影响,并最终落脚于物种形成、适应性进化、拓殖、物种存续和灭绝事件等综合过程[6]。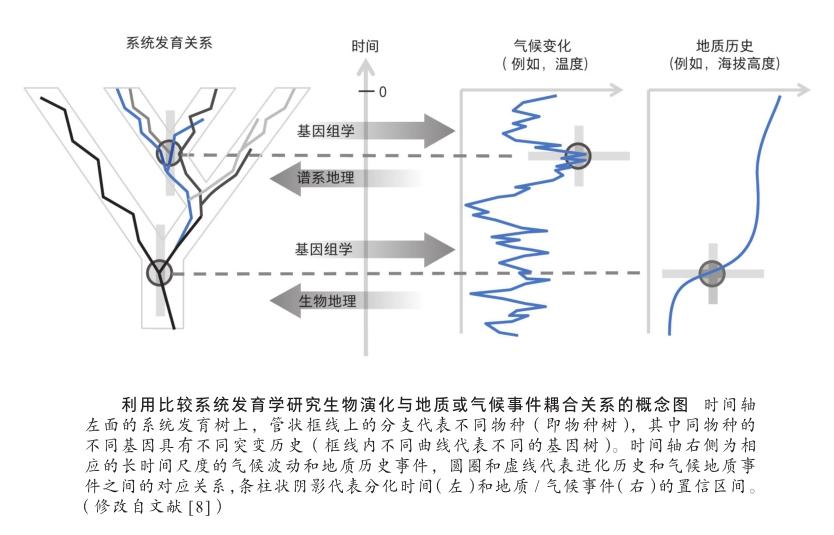
系统发育学的方法是现代历史生物地理学的根基,它由系统发生来推测祖先的分布。最早基于简约性原则,从现存分类群的分布重建其祖先类群的地理分布,其中扩散—隔离分化分析强调了扩散的重要性,它用扩散和灭绝来解释生物地理分布所需的最小代价,这被认为是最简约假说。目前,最流行的还是扩散—灭绝—分支进化模型(dispersal-extinctioncladogenesis)[7],该模型采用最大似然法推测物种扩散至新地区的历史、原分布区的灭绝,以及用隔离分化或者支序发生,解释分类群在不同区域间的分布式样,该理论框架也可用贝叶斯方法来实现。随着近年来分子生物学的发展,不仅能根据物种遗传距离构建物种的系统发育关系、重建生物类群的祖先分布区,还可以依此为基础结合化石校正分子钟,追溯物种的分化时间和演化历史。此外,还可利用比较系统发育学方法(comparative phylogenetic methods)估算物种的多样化速率,分析植物区系的成分来源是就地演化還是物种迁移,比较不同区系或类群的进化速率,推断其地理分布格局的形成历史、物种多样性随时间的变化等进化过程,并可判定历史事件(如地质历史和气候变化等)对现代生物多样性分布格局的影响,这些对于深入探讨多样性的起源有重要的意义。
高寒生物区的起源与演化
国际学术界针对高寒植物多样性形成的研究,主要集中在几大山地生物多样性热点地区。热带安第斯帕拉莫高山草原被认为是全球高寒植物多样性分化最快的高寒生物区,其起源时间大概在350万年前,与安第斯山北部地区隆升至现在高度的时间一致,研究发现其高寒类群主要来自低海拔的安第斯草原和临近的亚马孙地区。欧洲地中海高山地区高寒生物区的形成主要在更新世,而阿尔卑斯山等地高寒生物区可能形成于距今800—400万年, 但是该生物区大部分类群的多样化是在第四纪。新西兰的南阿尔卑斯山地区的高寒不早于500万年。东非高寒生物区的形成时间在距今800—700万年。极地苔原也是直到上新世北极永久冰盖形成之后才形成的[9]。因此,从目前全球高寒植物多样性的形成历史来看,高寒生物区是一个比较年轻的生物区,主要受到晚中新世以来各大洲山体最后阶段的抬升和第四纪冰期的驱动。另外,许多现在高山的年龄也都晚于渐新世,比如北安第斯山和东非高山;欧洲高山的隆升历史虽然可以追溯到中生代,但主要的隆升发生在距今1000—200万年间。当然也有例外,北美的落基山于始新世就已达到一定高度,很可能在晚始新世在针叶林以上存在过高寒生物区,但是该地区新近纪以来的强烈的气候变化(全球变冷和地中海气候的形成等)导致该地区古老的生物区发生了剧烈转变,且没有一直延续至今。其现今的高寒生物区是距今1000万年以来在经历气候转折期和新近构造运动的基础上才形成的。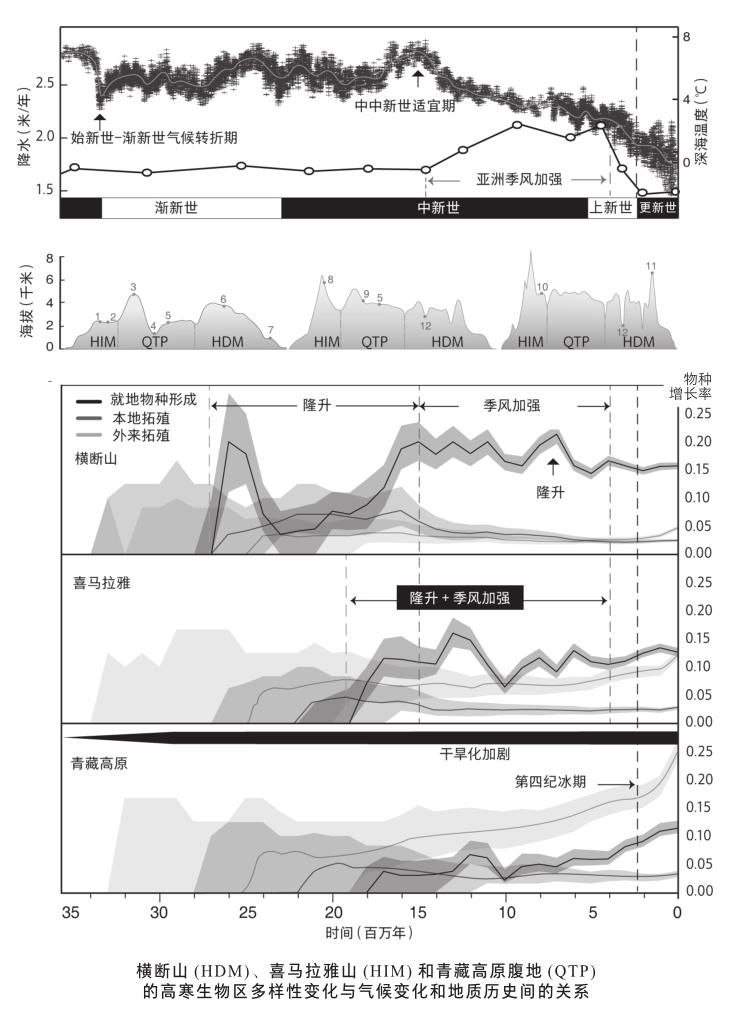
在笔者最新一项研究中,发现高寒植物在横断山可能自渐新世就已经出现(距今约3000万年前),并自晚渐新世到中中新世以来,物种演化速率快速增加[9]。这一重建结果,与独立根据化石证据和地质学证据得出的当时可能已存在海拔4000 米以上高山的情况吻合,也与横断山悠久的隆升历史,以及在区域构造和气候变化共同影响下形成复杂的地形地貌密不可分,这导致横断山高寒生物区可以在此基础上发展并存续下来。通过与其临近的喜马拉雅山和青藏高原高寒地区植物多样性的演化过程对比发现, 横断山高寒生物区不仅具有较高的就地演化速率,也是其他高寒地区植物多样性的主要来源,仅横断山就分别贡献了迁入喜马拉雅山和青藏高原的高寒类群总量的42%和44%,尤其自距今250万年以来,从横断山高寒地区扩散到喜马拉雅山和青藏高原高寒地區的速率明显加快,这表明在气候明显波动期,横断山地区是高寒植物的重要避难所。其南北走向的高山纵谷可为遭受极端气候的北方类群和高寒类群提供迁徙的廊道,而且山地动植物的垂直迁移也可在较短时间、较短距离内完成;与此同时,横断山一系列的高山和峡谷导致的地理和生境的巨大差异,也为物种的隔离分化创造了条件,成为许多动植物种类的新种分化地。更新世冰期时,由于全球变冷,山岳冰川发育,树线很可能比现在低,高寒生物区的面积比现在大;因此,邻近山地的高寒生物区由于树线的降低,彼此之间的交流也会增加。
在以往的谱系地理研究中也发现:不同物种在冰期、间冰期的扩张方式不同,但都不同程度地从横断山或其周边其他地区回迁到青藏高原腹地。即使是位于赤道附近的埃塞俄比亚高地和乞力马扎罗山,在冰期时其高寒生物区的海拔也比现在道低1000—1500米[10],使得冰期时高寒类群之间的联系更加密切。值得注意的是,在青藏高原内部大部分地区,即使是在末次盛冰期时,高原上面还有许多地区没有被冰川覆盖,所以冰期时高原面上仍然分布有大量植物的微型避难所。因此,现代高寒生物区是在漫长而复杂的造山运动和气候变化驱动下形成的。不同地区的高寒物植物多样性形成过程,由于历史和现代环境差异,呈现出明显的时空上的不同演化特征。而古老的横断山高寒植物区系,在全球高寒植物的演化过程中又在其中扮演着什么样的角色,仍值得进一步探索。
[1]K?rner C. The alpine life zone//Alpine plant life: functional plant ecology of high mountain ecosystems. K?rner C, 2 ed. Berlin: Springer, 2003: 9-20.
[2]武素功, 杨永平, 费勇. 青藏高原高寒地区种子植物区系的研究.云南植物研究 1995, 17(3): 233-250.
[3]Bouff ord D E. Biodiversity hotspot: Chinas Hengduan Mountains. Arnoldia, 2014, 72(1): 24-35.
[4]Hughes C E, Atchison G W. The ubiquity of alpine plant radiations: from the Andes to the Hengduan Mountains. New Phytol, 2015, 207(2): 275-282.
[5]Li X H, Zhu X X, Niu Y, et al. Phylogenetic clustering and overdispersion for alpine plants along elevational gradient in the Hengduan Mountains Region, southwest China. J Syst Evol, 2014, 52(3): 280-288.
[6]Rahbek C, Borregaard M K, Antonelli A,et al. Building mountain biodiversity: geological and evolutionary processes. Science, 2019, 365(6458): 1114-1119.
[7]Ree R H, Moore B R, Webb C O, et al. A likelihood framework for inferring the evolution of geographic range on phylogenetic trees. Evolution, 2005, 59(11): 2299-2311.
[8]Baker P A, Fritz S C, Dick C W, et al. The emerging field of geogenomics: constraining geological problems with genetic data. Earth Sci Rev, 2014, 135: 38-47.
[9]Ding W N, Ree R H, Spicer R A , et al. Ancient orogenic and monsoon-driven assembly of the worlds richest temperate alpine fl ora. Science, 2020, 369(6503): 578-581.
[10]Assefa A, Ehrich D, Taberlet P, et al. Pleistocene colonization of afro-alpine “sky islands” by the arctic-alpine Arabis alpina. Heredity, 2007, 99(2): 133-142.
关键词:高寒生物区 生物多样性 横断山 生物地理 ■
猜你喜欢
环球人文地理·评论版(2016年5期)2017-01-03
农业与技术(2016年15期)2016-11-09
河北渔业(2016年9期)2016-11-05
现代交际(2016年14期)2016-08-15
科技视界(2016年4期)2016-02-22
