
木论喀斯特常绿落叶阔叶林木本植物萌生特征
2021-09-01 10:03鲁梦珍宋同清彭晚霞刘坤平曾馥平
生态学报 2021年15期
鲁梦珍,杜 虎,宋同清,彭晚霞,刘坤平,苏 樑,曾馥平
1 中国科学院亚热带农业生态研究所亚热带农业生态过程重点实验室, 长沙 410125 2 中国科学院环江喀斯特生态系统观测研究站, 环江 547100 3 中国科学院大学, 北京 100049
森林更新是群落演替和植被恢复的重要过程,萌生更新与实生更新是森林更新过程中两种截然不同的更新方式[1-3]。实生更新是通过产生种子进行繁殖的有性生殖,萌生更新则主要指干基萌生和地下茎萌生,通过营养生长的方式形成多茎干的丛生植株[4]。萌生是植物抵抗干扰的一种重要方式,在遭受严重干扰后生物量大量损失的情况下,萌生植物能够利用地下茎和残留体(树桩)产生萌生茎干来实现自我更新,进而占领空间资源,坚守其固有的生态位,这就是“驻留生态位”[1,5-6]。干扰产生后,许多植物因为具有较强的萌生能力而保存下来,而一些萌生能力较弱的植物则有可能灭亡[5]。由此,萌生更新常常被认为是植物应对干扰和胁迫从而维持物种和种群稳定的一种特殊的生存策略。
我国西南喀斯特地区生态系统不稳定且抗干扰性差,导致生境高度异质、岩石出露率高、土壤瘠薄、土壤总量和容量低,形成水土流失严重等自然胁迫环境[7],其适生植物表现出一定的萌生特性[8],在喀斯特这种特殊的生境条件下,植物的实生更新受到限制,表现出种子数量少、传播和萌发困难,幼苗更新慢等特征[9],大多数植物具有较强的萌生能力[10]。目前,国内对植物产生萌生现象的机制以及环境因子对萌生能力的影响等相关方面的研究较少[11],仅有一些对少数样地萌生状态的调查[12]、营林树种萌蘖能力的研究[13]、以及对特殊干扰情况下个别树种萌生能力的单独研究[14]。对于喀斯特生境下植物的萌生特征、萌生能力与地形因子的适应关系及萌生对群落生物多样性的影响等方面有待进一步研究。本研究基于对木论国家级自然保护区大型森林动态监测样地的调查,以25 hm2大样地内萌生植物为研究对象,分析样地内萌生植物的萌生物种数量特征和植物的萌生能力与群落多样性的关系,揭示了萌生能力与地形因子的关联性和萌生更新在群落更新中的作用,以期为干扰后退化的喀斯特森林生态系统前期的植被恢复提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
木论国家级自然保护区位于广西环江毛南族自治县西北部(25°07′—25°12′ N, 107°54′—108°05′ E), 总面积10 800 hm2,海拔400—1000 m,为中亚热带石灰岩常绿落叶阔叶林(隐域性喀斯特森林植被顶极群落)分布区。该区域属亚热带季风气候区,年平均气温19.4℃,年降水量1500 mm,集中于4—9月,年均无霜期310 d,年均相对湿度为79%[15]。2014年在该保护区内,按照CTFS(the Centre for Tropical Forest Science)的标准建立25 hm2(500 m×500 m)固定永久监测样地(25°8′ N、108°0′ E),样地为正方形,东西、南北长均为500 m,样地海拔在442.6—651.4m之间。整个样地生境复杂,包括了从山顶、山坡到洼地等系列典型的“峰丛-洼地”生境类型。
1.2 植被调查
木论喀斯特常绿落叶阔叶混交林25 hm2样地于2014年完成首次植被调查,调查前运用全站仪和RTK将整个样地划分成625个20 m×20 m样方。本次研究以多茎干树种(multi-stemmed trees)的萌生现象来代表萌生更新,即新的茎干从主茎基部萌生出来,形成一丛多茎干的萌生植株。在样地调查时,记录萌生树种的种类、主茎胸径、新茎的数量与胸径等[16]。植被调查同时,调查各样方岩石出露率和土层厚度[15]。
1.3 数据处理
萌生能力指标的计算均以样方(20 m×20 m)为单位。本研究采用萌生率(Ratio of sprouting)、群落(样方)萌生物种丰富度比例(Richness proportion of sprouting species)、群落萌生物种个体比例(Abundance proportion of sprouting species)3个指标表征物种和群落水平的萌生能力[17]。其中萌茎数表示植物发生于其主干根部上的新茎数量,萌生物种丰富度指群落内已经发生萌生现象的物种种类,群落萌生个体数指群落内已经发生萌生现象的树种个体数量。具体公式如下:
萌生率=某一物种(或样方)的萌茎数/该物种(样方)所有个体数
群落萌生物种丰富度比例(RS)=萌生物种丰富度/群落总物种丰富度(表示群落萌生的物种数)
群落萌生物种个体比例(AS)=群落的萌生个体数/群落的所有个体数(表示萌生现象普遍性)
所有统计分析与作图均在R 3.3.6(R Development Core Team)中完成。采用回归分析方法分析萌生能力指标与群落多样性、环境因子间的关系。采用test检验常绿与落叶类群萌生能力之间的差异。使用vegan软件包计算多样性指数,基于建样地时获得的海拔数据计算出各样方的平均海拔、坡度、坡向和凹凸度地形因子,环境因子共包括土层厚度、岩石出露率、海拔、坡度、坡向、凹凸度。
2 结果与分析
2.1 样地内萌生物种的基本情况
本次调查共发现具有萌生现象的物种共197种,隶属59个科137个属,分别占样地植物科属种的86.8%、93.7%、91.3%,发生萌生的个体总数为20815株,占样地总个体数的20.0%。样地仅有30种物种不具有萌生现象。在木论大样地内仅有1个萌枝的个体占大多数,较少个体存在多个萌枝,单个植株萌生数7个以上的不足500株(图1)。样地每个样方的萌生个数在10—30个之间呈现单峰分布,每个20 m×20 m样方的萌生数会趋于一个稳定区间,此外,样方数和每个样方的萌生数在整体上呈现出反“J”型分布(图2)。

图1 木论大样地不同萌生数的植株数量统计

图2 木论大样地每个样方的萌生数
通过对样地内个体数量最高的前20个物种萌生率、萌生个体比例及萌生能力分析(表1),其萌生个体占整个样地萌生个体的64.23%。样地中萌生个体数较多的物种有小果厚壳桂(Cryptocaryamicrocarpa)、香叶树(Linderacommunis)、滇丁香(Luculiaintermedia)等。优势物种的萌生能力存在较大差异,萌生能力较强的物种是滇丁香、长管越南茜(Rubovietnamiaaristata)、火棘(Pyracanthafortuneana)、香叶树、米念芭(Tirpitziaovoidea)等。从谱系结构上看,萌生物种主要集中在茜草科滇丁香属和越南茜属、蔷薇科火棘属、亚麻科青篱柴属、樟科山胡椒属等类群。不同生活型植物的萌生能力存在显著差异(T=38.178,P<0.001),其中,常绿类群的萌生能力(0.75±0.01)远远高于落叶类群的萌生能力(0.25±0.01)(图3)。

图3 不同生活型的萌生能力

表1 样地内个体数排名前20的萌生物种萌生特征
2.2 萌生物种与群落多样性的关系
在调查的625个群落样方内,已经发生萌生现象的物种占群落物种丰富度比例的8.33%—88.89%,平均值为41.82%。萌生物种丰富度比例与物种丰富度(R2=0.01,P=0.011)、Shannon-wiener指数(R2=0.05,P<0.001)以及Pielou均匀度指数(R2=0.09,P<0.001)之间均表现出显著的正相关关系(图4)。群落的萌生能力在8.62%—56.47%之间,平均值为19.39%,群落萌生能力与物种丰富度(R2=0.08,P<0.001)、Shannon-Wiener指数(R2=0.17,P<0.001)、Pielou均匀度指数(R2=0.22,P<0.001)之间均表现出显著的正相关。群落萌生能力随着群落多样性指数的增大显著增强(图4)

图4 木论大样地中萌生物种丰富度比例和个体比例与群落物种多样性指数之间关系
2.3 植物萌生现象与环境因子的关系
从图5可以看出,土层厚度、海拔和凹凸度对群落RS均有显著影响(P<0.05)。群落(样方)水平的RS随土层厚度的增大呈显著下降的趋势(P=0.041),随海拔的升高和凹凸度的增加呈显著上升的趋势(P<0.001),土层厚度、海拔和凹凸度对群落RS的解释量分别为13.3%、9.5%、12.7%。由图6可知,土层厚度、岩石出露率、坡度和凹凸度对群落(样方)AS均有显著影响(P<0.05),除土层厚度与群落AS呈负相关外(P=0.002),群落AS随岩石出露率、坡度和凹凸度的增加均呈显著上升的趋势(P<0.001),四个环境因子对群落RS的解释量分别为24.5%、20.4%、16.3%、23.3%。

图5 木论大样地中萌生物种丰富度比例与环境因子之间关系
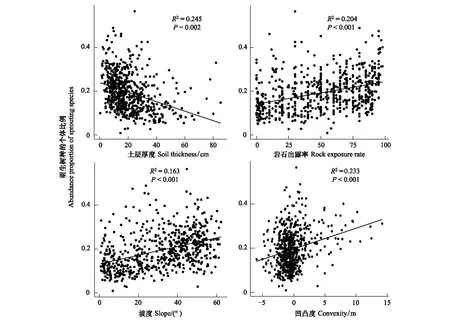
图6 木论大样地中萌生物种多度比例与环境因子之间关系
3 结论与讨论
木论25 hm2样地中具有萌生更新现象的物种丰富,样地内86.8%的物种都存在萌生更新现象,不具萌生更新现象的物种仅占13.2%,由此可见,萌生现象在木论大样地木本植物中非常普遍。植物的萌生能力受生境、干扰、资源等因素的影响[3],相比于热带喀斯特森林生态系统[10]和亚热带常绿森林生态系统[16],独特的地形和气候条件共同促进了木论样地特殊生境的形成,这可能是木论喀斯特森林生态系统萌生更新现象较高的潜在原因。木论25 hm2样地中部分生境土壤贫瘠, 岩石出露率较高, 坡度较大,不利于种子萌发,由于萌生植株强大的根系,能够有效地从土壤中吸收养分[18],使得萌生更新能够为幼苗提供更多的物质基础,从而提高幼苗的存活率和生长速度[19]。同时,萌生植物树皮的厚度与胸径的比值显著高于实生种子更新的植物,有助于植物抵抗外界干扰[20]。另外,在受到干扰后,由于萌生植株的“持续生态位”(persistence niche)效应,帮助植物降低生物量损失,加快群落演替的过程[8]。这些独特的优势增强了萌生植株对环境的适应力和对资源的利用能力,从而导致萌生更新在喀斯特常绿落叶阔叶林内地位突出,萌生更新作为一种重要的森林更新方式,对于生态系统的维持和稳定有着重要作用[21]。样地内优势物种的萌生能力存在较大差异,说明不同物种有不同的更新策略。本研究中,一些物种如茜草科的滇丁香和长管越南茜、蔷薇科的火棘、樟科的香叶树等,较其他物种更倾向于采取萌生更新策略,一方面原因可能是由于这些树种自身竞争力强。另外,这些树种多生活在土壤贫瘠,岩石出露率高,地形坡度大生存条件恶劣的位置,进一步说明萌生是物种在遭遇干扰后为维持种群更新所作出的适应,反映出萌生能力与地形因子存在一定的关联性。
样地内常绿类群的萌生能力显著高于落叶类群。样地调查的227种木本植物中,有142种为常绿树种,85种为落叶树种。研究区内高温多雨且分布不均匀、碳酸盐岩溶蚀性强[22],上坡位光照强烈、土壤贫瘠,为适应干旱等恶劣条件,植物通过落叶来减少叶片水分及养分的消耗[23],而常绿树种多生活在水分充足的洼地,受雨水冲击大,幼树容易遭受水淹、落石等毁灭性干扰,使得植物多采取萌生更新策略以迅速恢复生态位,这可能是样地内常绿树种较落叶树种萌生能力强的原因之一。
在广西木论喀斯特常绿落叶阔叶林中,随着萌生物种种类的增多和萌生物种多度的增加,群落物种多样性指数逐渐上升。从萌生物种分布特征来看,萌生树种具有高度聚集分布的特点,通常会因茎干之间在资源(光照、营养、空间等)获取上存在激烈的竞争,导致植株的死亡率较高,不容易形成长期固定的空间格局,从而为其它个体和物种的补充提供了机会,这在一定程度上增加了群落生物多样性。研究区喀斯特常绿落叶阔叶林面积大,地形复杂多样,植物在容易发生周期性水淹的地区受干扰大,萌生力强的物种通过萌枝迅速进行自我更新维持种群的繁衍,从而能够维持原有物种不灭亡[1, 5-6],这样,该种群能够在此区域保持竞争优势,长期存在,而木论样地中干扰强度低、土壤生产力相对较高,种子更新占优势的地区,木本植物的幼苗存活率高,当有轻微干扰时,既保留了原有物种的生存,同时也给群落外其它物种提供迁入的机会,在一定程度上提高了群落生物多样性。对西双版纳刀耕火种弃耕地的群落演替研究也表明,萌生会导致群落演替前期生物多样性较高[24],其主要原因可能是由于在群落演替早期, 萌生植物通过自我更新改善群落生境,为其他物种的定居提供条件,以此来增加群落生物多样性[25-26]。而叶铎等[17]对浙江古田山亚热带常绿阔叶林中的研究发现,萌生会降低群落生物多样性,主要是由于萌生植株在资源利用、种间竞争上比其他物种占优势[4, 27],萌生植株占领生态位后使其他物种难以定居,从而降低群落内物种多样性。另外,萌生与实生更新不同,萌生是通过残留器官来产生萌枝[2],因此缺少代际间遗传信息的交流,所以在一定程度上降低了群落内的物种周转率[28-29],最终导致群落生物多样性下降。
萌生更新和种子更新是植物两种截然不同的更新策略,它们之间的协调主要体现在物质分配和竞争上。一般来说,干扰发生的频率和干扰强度影响植物繁殖对策的转换。在没有干扰的情况下,由种子更新形成的单干型树木具有较强的资源竞争优势[30],但是由于缺乏“持续生态位”效应,留给群落外其他物种迁入的机会较多,因此干扰后植物群落结构和种类组成将发生根本性改变[31]。相反,在高频率干扰下,萌生种类具有资源竞争优势,使群落外其他种类难以有入侵定居的机会,群落内因保留了较多原有萌生能力强的物种而降低了物种多样性[31]。木论样地内洼地受雨水冲击大且人为干扰严重,群落内树种往往选择萌生更新的方式繁殖后代,使得其他物种难以入侵,导致群落内生物多样性较干扰频率低的地区低。
生境假说认为,萌生更新能力随树木的生长环境不同而不同[3]。本研究发现,土层厚度对木论样地木本植物群落的萌生能力影响较大,土层厚度增加会降低物种的萌生能力,这可能是由于在土层较薄的地区,土壤生产力低,实生更新的种子生长受到限制,因此通过增强萌生能力来维持种群的稳定[32]。另一个可能的原因是在土壤贫瘠的地区,贮水能力不高,容易受雨水冲刷的干扰,一方面限制了生态系统的有性更新,另一方面由于干扰强度高,植物必须通过萌生来实现快速更新,恢复原有生态位。本研究结果显示,凹凸度和坡度的增加会提高植物的萌生能力。这一结论与郭屹立等[10]结论相似,萌生能力与凹凸度呈正相关,凹凸度越小生境越潮湿。这可能是由于凹凸小的生境中,水分条件充足,土壤生产力高,但是光照条件受限制,植物更倾向于向径直生长,以获取充足的光照维持自身的需要,而在凹凸度大的生境中,光照强烈,水分土壤条件差,植物个体只有通过横向生长占据横向的生态位空间。坡度通过影响植被的受干扰情况来影响植物的萌生能力[33],在喀斯特特殊地形的生境中,通常坡度大的生境受水分条件影响较大,更容易遭受雨水冲刷,使得土层不稳定,植物在这些易受干扰的地段上萌生能力通常更强,这一现象进一步支持了Belingham[33]等和Ye等[34]对萌生植物的研究。
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
落叶果树(2021年6期)2021-02-12
乡村地理(2019年2期)2019-11-16
————水溶蚀岩石的奇观
家教世界(2019年4期)2019-02-26
现代园艺(2018年1期)2018-03-15
体育科技(2016年2期)2016-02-28
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
文化月刊·下旬刊(2014年6期)2014-08-28
