
33份引进花生资源表型遗传多样性分析及在粤西地区的适应性初步评价
2021-08-26 21:36徐志军吴小丽胡小文刘洋
热带作物学报 2021年7期
徐志军 吴小丽 胡小文 刘洋
摘 要:為了解引进花生资源的遗传多样性和适应性,筛选可用于广东地区花生遗传改良的优异资源和高产品种,本研究对33份从国内收集的花生资源进行表型鉴定和在粤西地区的适应性初步评价。结果表明:13个农艺和产量性状的平均变异系数范围为4.80%~30.80%,13个性状的平均遗传多样性指数范围为2.52~2.90,遗传多样性丰富。相关分析表明,多数性状间的相关关系在2019年春、秋两个环境保持一致,部分性状间的相关性受环境因素影响。聚类分析可将33份资源划分为4个类群。主成分分析中,前3个因子累计贡献率为82.19%,可以反映表型性状的绝大部分信息。通过表型性状综合评价筛选出10份综合排名超过全部本地对照品种的资源。适应性评价表明引种后种子长、百果重、百果仁重、百仁重和出仁率5个性状出现极显著下降,其中‘花育23在2019年秋季的表现优于引种前,其余资源在这些性状上均有不同程度的下降;引进资源对早斑病和晚斑病抗性较弱,多数表现为感病;丛枝病发病鉴定表明共有19份资源感病,14份资源未见发病。研究结果可为引进资源在广东地区的利用和品种选育提供重要参考。
关键词:花生;引种;遗传多样性;综合评价;适应性
中图分类号:S565.2 文献标识码:A
Phenotypic Diversity of 33 Introduced Peanut Germplasm Accessions and Preliminary Adaptability Evaluation in West Guangdong
XU Zhijun1, WU Xiaoli2, HU Xiaowen1, LIU Yang1*
1. Zhanjiang Experiment Station, Chinese Academy of Tropical Agricultural Sciences / South Subtropical Crop Research Institute, Chinese Academy of Tropical Agricultural Sciences / Guangdong Engineering Technology Research Center for Dryland Water-saving Agriculture, Zhanjiang, Guangdong 524091, China; 2. Zhanjiang Academy of Agricultural Sciences, Zhanjiang, Guangdong 524094, China
Abstract: The phenotypic diversity and adaptability of 33 introduced peanut (Arachis hypogaea) germplasm accessions were evaluated in this study, aiming to understand and utilize the germplasm accessions in Guangdong. The result revealed that the mean coefficient of variation of 13 agronomic and yield related traits ranged from 4.80% to 30.80% and the mean diversity index ranged from 2.52 to 2.90, indicating the introduced germplasm maintained abundant genetic diversity. Correlation analysis showed that the correlationship between most traits kept stable in the spring and autumn of year 2019, while the correlationship between some traits was instability and affected by environmental factors. The 33 introduced peanut germplasm accessions were divided into four groups by cluster analysis. The first three principal components explained 82.19% of the phenotypic variation, reflecting most information of the 13 traits. Ten germplasm accessions were found to excel the 5 local control cultivars through comprehensive evaluation of phenotypic traits. Seed length, weight of 100 pods, weight of seeds in 100 pods, weight of 100 seeds and shell percentage were significantly decreased after introduced. And ‘Huayu 23 was better before introduced in autumn of year 2019, while the remain germplasm accessions showed different degree of decline in the five traits. The resistance of the introduced germplasms was poor to early and late leaf spot and most of them were susceptible. There were 19 germplasm infected peanut witches-Broom, while 14 germplasm were not infected through field investigation. These results would provide important reference for utilizing the introduced germplasm accessions to breed new peanut cutivars in Guangdong.
Keywords: peanut; variety introduced; genetic diversity; comprehensive evaluation; adaptability
DOI: 10.3969/j.issn.1000-2561.2021.07.010
花生(Arachis hypogaea)是我国重要的经济作物、油料作物和食用作物,在国民经济发展和对外贸易中具有重要地位,在保障我国食用油安全和供给方面具有重要意义。花生在我国广泛种植,种植面积在国内大宗作物中位居第七,种植业产值仅次于水稻、小麦和玉米,种植效益优势明显[1]。花生在广东地区的种植面积超过3.32× 105 hm2,是广东地区最主要的油料作物。但是,与国内其他省份相比,广东地区的花生单产仍处于较低水平。2018年,广东地区的花生平均单产仅为3135 kg/hm2,显著低于同期全国平均水平(3750 kg/hm2),与新疆(4755 kg/hm2)、河南(4740 kg/hm2)、山东(4410 kg/hm2)等地区花生平均单产差异还较大[2]。品种引进是丰富种质资源遗传多样性的有效手段,也是提高作物产量的重要方式之一[3-4]。因此,对外地引进的花生品种进行表型鉴定和适应性评价,开展遗传多样性研究,筛选优异育种资源和适宜本地种植的高产品种,对于广东地区花生产业发展具有重要的意义。
作物的表型是基因表达和环境因子共同作用的结果[5],是作物遗传结构的直观反映,也是研究作物遗传多样性的传统方法[6],因其直观、便利、成本低的特点,而被广泛地用于作物种质资源评价中。适应性评价是引进资源利用的前提,通过对初引进资源进行评价,能够及早发现引进资源可能出现的退化、品质下降、病虫害等问题,为引进品种的栽培、管理、繁殖和利用提供依据[2]。在花生上,李付振等[7]、姜瑶等[3]先后对外地引进的花生品种进行了表型鉴定和适应评价,鉴定出适宜浙江和河北黑龙港低平原地区种植的花生品种;陈崇果等[8]通过表型鉴定,分析了12个北方花生品种在黔中地区的适应性,筛选出适宜黔中地区2个高产花生品种。本课题组2018年从其他省份收集了33份花生资源,这些资源以育成品种为主,包含少量的农家种和品种衍生品系。本研究以这些资源为实验材料,对其13个表型性状进行鉴定,通过相关分析、聚类分析、主成分分析、引种前后差异显著性分析和抗性鉴定,了解引进花生资源的遗传多样性和在广东粤西地区的适应性,从而为引进资源在广东地区的利用和品种选育等提供参考。
1 材料与方法
1.1 材料
供试材料为33份花生资源,于2018年从河南、山东、河北、四川、吉林、黑龙江和云南地区收集,以广东省粤西地区5个主要的花生栽培品种作为对照(表1)。
1.2 方法
1.2.1 表型鉴定 2019年春播前,调查引进花生资源的荚果长、荚果宽、荚果长宽比、种子长、种子宽、种子长宽比、百果重、百果仁重、百仁重、出仁率10个性状。2019年春、秋两季连续在中国热带农业科学院湛江实验站基地,对引进的资源进行种植,每份材料种植1个小区,每小区3行,每行单粒种15粒,3次重复,行距30 cm,株距15 cm,常规田间管理。收获时调查主茎高、第一侧枝长和总分枝数,收获后调查荚果长、荚果宽、荚果长宽比、种子长、种子宽、种子长宽比、百果重、百果仁重、百仁重、出仁率。叶斑病和丛枝病抗性鉴定,按照前述方式进行种植,整个生育期不使用杀菌剂和杀虫剂。所有性状调查按照《花生种质资源描述规范和数据标准》[9]执行。
1.2.2 适应性评价 通过对种植前后的荚果和种子10个性状的差异显著性分析,并对引进资源在湛江地区叶斑病抗性和丛枝病发病情况进行鉴定,对资源的适应性进行初步评价。
1.3 数据分析
1.3.1 多样性分析 参照孙珍珠等[10]的方法对性状划分等级,即±kσ(其中k=0、0.5、1.0、1.5、2.0,σ为标准差),将这些性状表型值划分10个等级,每1级中材料所占的比例(Pi)用于计算多样性指数,遗传多样性指数即Shannon-Wiener index(H′)信息指数,其计算公式为:H′=-ΣPi×lnPi,式中Pi为某性状第i级中包含的材料份数所占材料总份数的百分比。
1.3.2 数据标准化 通过模糊隶属函数法将33份花生种质的13个表型性状函数值进行标准化,计算公为:X(ij)=(Xij-Xjmin)/(Xjmax-Xjmin)。X(ij)表示隶属函数值,Xij为品种i在性状j的测定值,Xjmin与Xjmax为供试品种表型j的最小值和最大值。
1.3.3 聚类分析 参照董胜君等[11]的方法对表型标准化后的数据采用Ward法以平方欧氏距离为遗传距离进行聚类分析。
1.3.4 综合评价 参照综合评价方法[12],使用R软件包对33份花生资源的13个表型性状进行主成分分析。参照1.3.2中的方法使用隶属函数对主成分分析因子得分系数矩阵进行标准化,结合权重系数来计算综合评价F值。
1.3.5 适应性评价 使用Origin 8.0软件对33份花生资源引种前后荚果和种子10个性状进行差异显著性检验。
使用Excel、R和Origin 8.0软件进行数据统计分析和绘图。
2 结果与分析
2.1 花生资源的变异及多样性
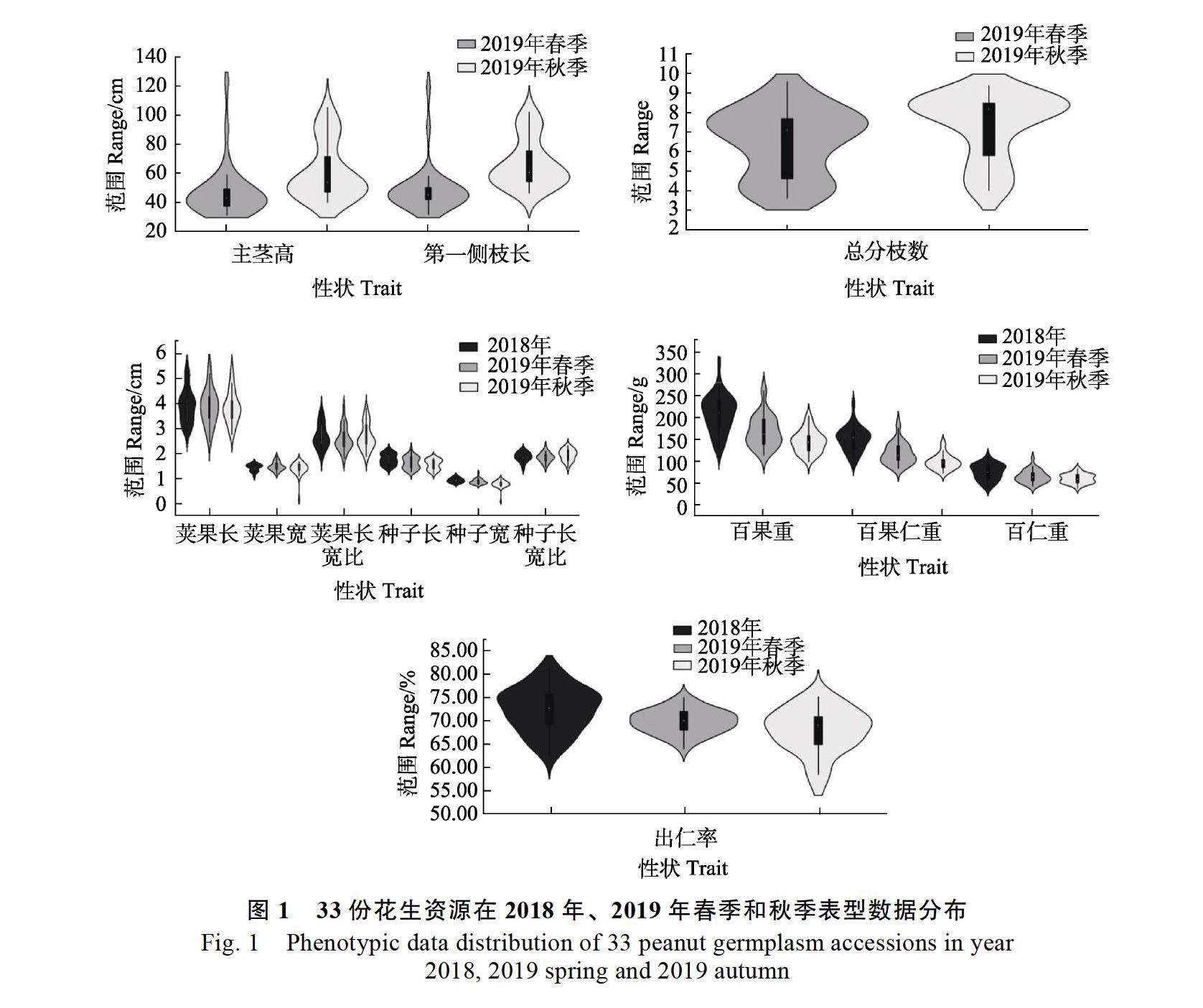
33份花生材料2019年春、秋两季表型鉴定表明,13个农艺和产量性状的表型分布和变异差异明显(图1,表2)。13个性状在2019年春季的变异系数范围为3.66%~37.33%,2019年秋季的变异系数范围为7.05%~31.35%,两季平均为4.80%~30.80%,其中主茎高、第一侧枝长、总分枝数变异系数较大,均在20%以上,出仁率变异系数最小,平均为4.80%,表明该性状与其他性状相比受环境影响较小。13个性状的平均遗传多样性指数范围为2.52~2.90,遗传多样性丰富,多样性指数在2019年春季和秋季存在一定的波动,其中主茎高变化最大,荚果长宽比变化最小,表明环境对不同性状的影响不同。对33份材料2019年春季和秋季两个环境的数据进行单因素方差分析表明,主茎高、第一侧枝长、种子宽、百果重、百果仁重春秋两季表现出极显著差异(P<0.01),总分枝数、出仁率在春秋两季表现出显著差异(P<0.05),其余性状在春秋兩季受环境影响较小(表2)。
2.2 花生资源表型性状间的相关性分析
对13个表型性状进行相关性分析表明,多数性状的相关关系在2019春、秋两个环境保持稳定,部分性状间的相关关系受环境因素影响较大(表3)。在2019年春、秋两季,主茎高与第一侧枝长、荚果长、荚果长宽比呈极显著正相关,与总分枝数呈极显著负相关;第一侧枝长与总分枝数呈极显著负相关,与荚果长、荚果长宽比呈极显著正相关;荚果长与荚果长宽比、种子长宽比、百果仁重呈极显著正相关;荚果宽与种子宽呈极显著正相关;荚果长宽比与种子长宽比呈极显著正相关;种子长与种子长宽比、百果重、百仁重呈极显著正相关,与百果仁重呈显著正相关;百果重与百果仁重和百仁重呈极显著正相关;百果仁重与百仁重呈极显著正相关。其中主茎高和第一侧枝长、百果重和百果仁重相关系性最高,在2019年两季相关系数分别为0.9729、0.9551和0.9855、0.9172。部分性状间的相关,如百果重与主茎高、第一侧枝长、总分枝数间在2019春季表现出相关关系,但在2019秋季相关关系不显著,可能是受环境影响表现出不稳定的相关关系。
2.3 基于13个表型性状的聚类分析
对33份花生资源和5个对照品种使用2019年表型性状的平均值进行聚类分析,在欧式距离为1.6时,可以将38份材料分为4个类群(图2)。类群Ⅰ包括13份材料,其中以‘四粒红为代表的多粒型材料聚成一个均为红花生亚群a,该类群株高、第一侧枝长普遍较高,且总分枝数较少;另一个亚群b主要为普通型和中间型。类群Ⅱ包括22个品种,其中亚群c主要为珍珠豆型,为小粒或中粒型花生;亚群d主要为普通型,为大粒型花生。类群Ⅲ包括1个品种‘鲁花11;类群Ⅳ包括2个多粒型花生品种。38份花生材料表型聚类结果能够反映品种的部分遗传信息,性状间的聚类与性状间相关关系一致。
2.4 花生资源表型性状的主成分分析
主成分分析可在不损失或很少损失原有信息的前提下,将原来个数较多且彼此相关的指标转换为新的个数较少且彼此独立的综合指标[13]。对13个表型性状进行主成分分析,前3个主成分特征值均大于1,累计贡献率82.19%,可解释表型性状的绝大部分信息(表4)。其中第1主成分特征值最大,为5.41,贡献率为41.59%,其中主茎高、第一侧枝长、总分枝数、荚果长和荚果长宽比的特征向量绝对值较大,说明该主成分主要反映株型和产量有关的信息;第2主成分,贡献率为28.00%,其中百仁重的特征向量绝对值最大,其次是种子长、百果重、荚果宽、百果仁重、种子宽和出仁率,表明该成份主要反映产量有关的信息;第3主成分贡献率为12.60%,荚果宽和种子长宽比特征向量绝对值最大,也主要反映产量有关信息。
2.5 花生资源表型性状的综合评价
根据主成分分析获得3个因子得分公式:
F1=0.3926X1+0.3923X2–0.3537X3+0.3903X4–
0.1242X5+0.3938X6+0.0292X7–0.2324X8+0.1938X9+ 0.2548X10+0.2805X11–0.0874X12+0.0197X13
F2=0.0737X1+0.0264X2–0.1310X3–0.1398X4– 0.3198X5+0.1004X6–0.4111X7–0.3133X8–0.1581X9–0.3834X10–0.3180X11–0.4649X12+0.2974X13
F3=0.1698X1+0.1506X2–0.2314X3–0.0268X4+ 0.4297X5–0.1450X6–0.4535X7–0.2903X8–0.5691X9+0.1712X10+0.1863X11–0.1014X12–0.0211X13
将F1、F2、F3得分使用隶属函数进行归一化后,根据各主成分的贡献率权重(0.5060、0.3406、0.1533)得出每个品种的综合得分,对33份花生材料表型性状进行综合评价(表5)。38份材料的综合得分范围为0.1938(‘海花1号)~0.7440(‘LXSL),平均值为0.4084。F值与13个表型性状进行相关分析表明,F值与主茎高、第一侧枝长、
荚果长、荚果长宽比呈极显著正相关,与总分枝数、荚果宽、种子长、种子宽、百仁重呈极显著负相关(表6),因此在对花生资源进行综合评价时F值越大,株高越高,子粒越小,增产潜力越小,排名越靠后(表5)。与5份对照品种进行比较发现,‘海花1号‘黑花生‘吉花22‘大白沙‘冀花2号‘鲁花1号‘吉花19‘冀花5号‘鲁花11‘拔二罐10份材料综合排名高于5份对照材料,这些材料在总分枝数、荚果和种子大小要优于本地品种;‘丰花1号‘豫花22‘花育51‘花育19号‘白沙1016‘冀花4号‘天府3号7份材料排名高于2份对照材料;‘花育910‘远杂9102‘豫花40‘花育234份材料排名高于1份对照材料。
2.6 花生资源的适应性评价
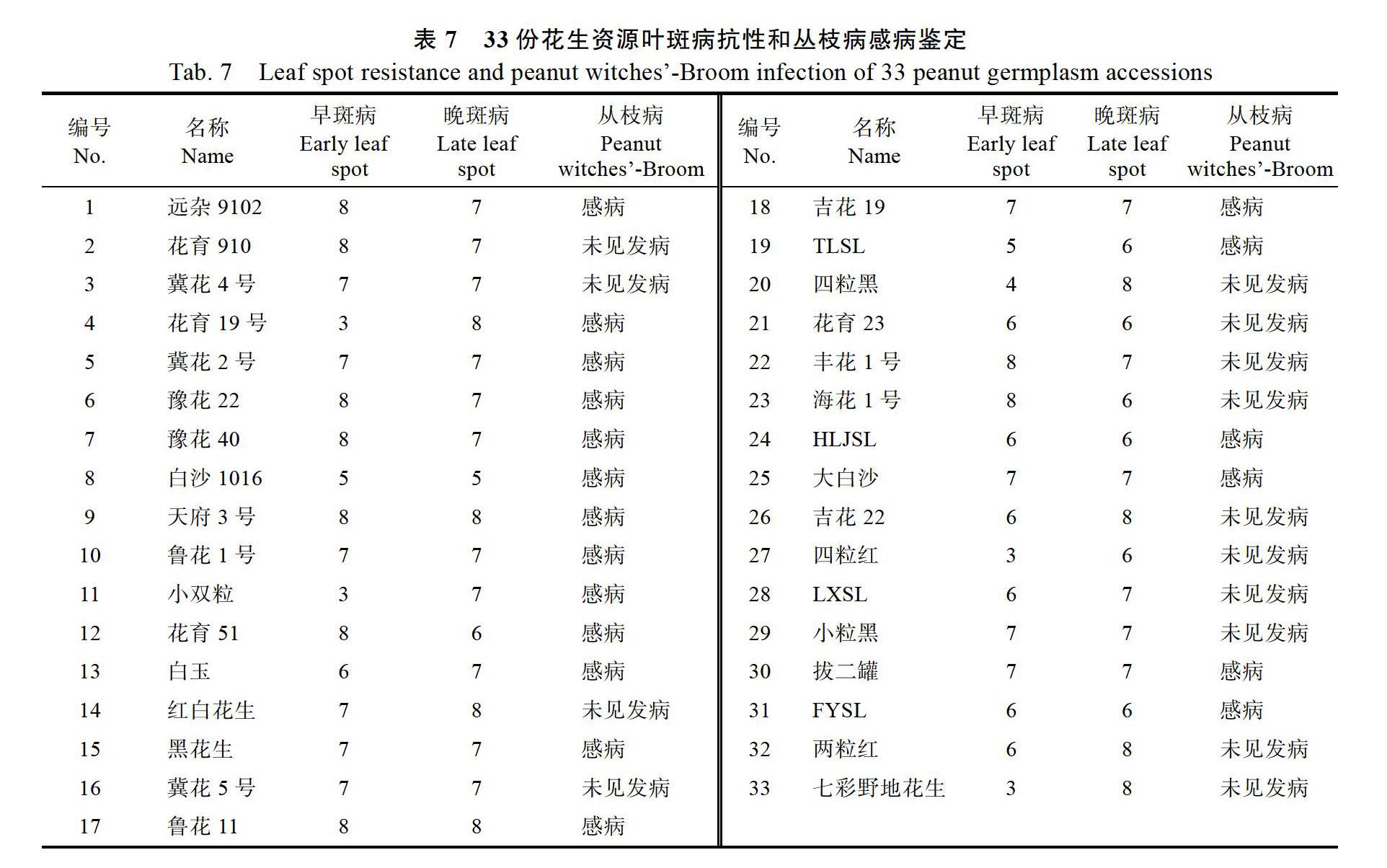
从图1可知,10个产量性状2018年(引种前)和2019年春、秋两季数据分布差异明显。如图3所示,从原产地引种到粤西地区(湛江)后,种子长、百果重、百果仁重、百仁重和出仁率5个性状在2019年春季、秋季和2019年平均值均极显著降低,种子宽在2019年春季显著降低,在2019年秋季和2019年平均值均極显著降低;荚果宽仅在2019年春季显著上升,荚果长宽比仅在2019年春季显著降低,荚果宽和荚果长宽比在2019年秋季和2019年平均值与引种前差异不显著;种子长宽比仅在2019年秋季显著上升;荚果长在2019年春季、秋季和2019年平均值均与引种前差异不显著。以上结果表明,引种后环境主要影响种子长、种子宽、百果重、百果仁重、百仁重和出仁率等性状。对单个品种在2019年春秋 两季的表型数据进行比较发现,2019年秋季,‘花育23在种子长、种子宽、百果重、百果仁重和百仁重5个性状上的表现均优于引种前,其余品种在种子长宽、果重和仁重上均有不同程度的下降,初步推测‘花育23对粤西地区秋季具有相对较好的适应性。对引进材料进行花生早斑病和晚斑病抗性鉴定表明,部分品种表现出早斑病或晚斑病抗性,如‘花育19‘小双粒‘四粒红中抗早斑病,‘白沙1016表现出晚斑病抗性,但抗性水平不高;引进资源多数对早斑病和晚斑病的表现为感病(表7)。花生丛枝病发病情况调查发现,有14份材料在春秋两季未见发病,19份材料感病(表7)。
3 讨论
表型鉴定是作物遗传研究的重要基础,是种质资源遗传多样性研究中最简单、最直观的方法。开展表型遗传多样性研究[13-14],有助于从整体上认识作物性状的多样性程度,为优异资源的挖掘和利用提供理论和实践依据[10,15-16]。在花生上,通过表型鉴定,姜慧芳等[17-18]以6390份栽培花生资源为材料,通过植物学类型分层和15个表型性状的排序和聚类分析,构建了花生核心种质和小核心种质;肖宇等[19]对福建省审(认)定的23个花生品种11个表型性状进行了评价,发现开花习性、种皮颜色和种仁大小3个性状表现一致;江建华等[20]对来源于5个国家的126份花生材料进行了遗传多样性研究,22个分级性状和17个农艺性状遗传多样性指数的均值分别为0.813和1.947,遗传变异丰富,用于遗传改良的潜力巨大;孙东雷等[13]对40份花生资源的17个表型性状进行了综合评价,筛选了8个可以作为花生资源进行评价的指标的性状,这些研究都促进了对花生资源的认识、研究和利用。本研究对从我国其他省份引进的33份花生材料的株型和产量相关性状进行了連续表型鉴定,3个株型和10个产量相关性状的平均变异系数范围为4.80%~30.80%,Shannon-Wiener多样性指数平均范围为2.52~ 2.90,变异和遗传多样性丰富,具备用于广东地区高产花生遗传改良的潜力。
聚类分析可以反映种质资源间的遗传差异和各类群的特点[21]。前人研究表明,基于表型与基于分子标记的聚类分析的结果基本一致,都能够有效地评价资源的遗传多样性[22-23]。本研究通过表型聚类将33份资源划分为4个类群,处于同一类群或亚群的资源之间具有较近的亲缘关系,如
类群Ⅰ亚群a中,‘TLSL‘HLJSL‘LXSL‘FYSL均是从‘四粒红中衍生出来的资源;类群Ⅰ亚群b中‘丰花1号‘花育23‘花育51都有‘伏花生的血缘且同属于山东地区培育的品种;类群Ⅱ亚群c中,‘白沙1016是‘远杂9102的母本,利用‘远杂9102为亲本又培育了‘豫花22和‘豫花40,这些品种聚到了同一个分枝;此外来源于同一种植区花生品种聚到同一分枝或者亚群,也反映了同一种植区花生品种遗传背景较为狭窄。这些结果表明本研究结果与前人一致[13, 19, 24],表型聚类结果能够直观地反映收集花生资源的遗传背景,可以有效用于对资源遗传背景的初步评估。
作物资源表型评价是育种研究的重要环节,基于隶属函数和主成分分析的综合评价,已在水稻[12]、陆地棉[25]、谷子[26] 和许多园艺作物[3, 11, 27]的表型评价中广泛应用。在花生研究上,孙东雷等[13]通过主成分分析,将17个性状划分成子粒含油量、含糖量和丰产三类因子,对40份花生资源的优劣进行了综合考察。本研究利用相同方法对33份花生材料的13个性状进行了综合评价,主成分分析将表型性状划分为株型和产量二类因子,通过计算引进材料和本地主推品种的综合得分,筛选出10份综合性状优于本地品种的花生资源,这些资源与本地品种相比产量潜力更高,可用于本地品种产量相关性状的遗传改良。
适应性评价是作物品种和资源引进利用中的关键环节[3, 7-8]。本研究发现从其他省份引进的花生资源在粤西地区种植后,除荚果长和宽、荚果长宽比、种子长宽比变化较小外,其余6个产量性状都存在着显著的下降,这6个性状是花生仁产量的重要组成因素。种子长和宽极显著下降是造成百果重、百果仁重、百仁重和出仁率下降的重要因素。种子变小或发育早衰是引种中常见现象,前人研究表明,日照长度、土壤积温、昼夜温差等因素都会影响花生仁的生长和发育[28-29],与原产地相比,广东地区土壤积温更高、昼夜温差更小,不利于花生生殖生长期的物质积累,可能会造成花生仁发育的早衰。对单个品种在2018和2019年比较发现,2019年秋季,‘花育23种子长、种子宽、百果重、百果仁重和百仁重5个性状上的表现均优于引种前,是潜在的秋花生利用资源,需要对该资源秋植进行进一步的评价。广东粤西地区属于热带季风气候,温度和湿度都适宜花生叶斑病的发生和传播,本研究对引进资源的叶斑病抗性鉴定发现,除极少数资源表现出叶斑病抗性外,大多数资源都表现出感病,部分在原种植区抗性较好的品种,也表现出感病,这可能是由于叶斑病小种分化和2019年粤西地区温湿度条件更适宜病原菌的繁殖和扩散造成的,另外叶斑病田间鉴定受环境影响较大,也是影响鉴定结果的重要因素。因此需要对这些资源,特别是在原种植区叶斑病抗性较好的资源进行进一步的抗性鉴定,对感病资源进行利用时,需要选择本地叶斑病抗性较好的品种和资源配置组合。花生丛枝病是广东等热带地区常见的花生病害,在秋花生上的发生尤为严重,一般由小绿叶蝉取食花生韧皮部将花生植原体传播到花生植株,导致植株严重矮化,节间缩短,腋芽大量萌发出丛生枝条,叶片细小黄化[30],感病花生荚果很少或者不结实,通常会造成感病花生荚果减少或不结实,严重影响花生的产量和品质[31]。前人研究发现植原体进入花生韧皮部后均会导致花生丛枝病[31],花生品种对小绿叶蝉的抗性是花生丛枝病发生存在差异的原因[32]。本研究通过连续两季田间鉴定初步获得14份未见发病的花生资源,这些资源通过进一步鉴定排除小绿叶蝉的取食偏好性,可以作为抗小绿叶蝉的花生材料用于花生丛枝病的遗传改良。
参考文献
[1] 廖伯寿. 我国花生生产发展现状与潜力分析[J]. 中国油料作物学报, 2020, 42(2): 161-166.
[2] 中华人民共和国国家统计局2018年数据[DB/OL] (2019-09-28) [2021-06-20]https://data.stats.gov.cn/easyquery. htm?cn=E0103/.
[3] 都真真, 李锡香, 宋江萍, 等. 228份引进大蒜资源的表型多样性分析及适应性初步评价[J]. 植物遗传资源学报, 2019, 20(5): 1186-1196.
[4] 姜 瑶, 王希全, 赵 杰, 等. 黑龙港低平原地区直播春花生高产品种筛选[J]. 河北农业大学学报, 2017, 40(3): 26-31.
[5] 周 波, 江海东, 张秀新, 等. 部分引进牡丹品种的形态多样性[J]. 生物多样性, 2011, 19(5): 543-550.
[6] 万映伶, 刘爱青, 张孔英, 等. 菏泽和洛阳芍药品种资源表型多样性研究[J]. 北京林业大学学报, 2018, 40(3): 110-121.
[7] 李付振, 宋度林, 吴学龙, 等.12个花生新品种在浙江的适应性试验[J].浙江农业科学, 2014 (10): 1523-1524, 1529.
[8] 陈崇果, 任明刚, 冯明友, 等. 12个北方花生新品种在黔中地区的适应性和丰产性差异分析[J]. 中国种业, 2019 (11): 52-56.
[9] 姜慧芳, 段乃雄, 任小平. 花生种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2006: 128-l33.
[10] 孙珍珠, 李秋月, 王小柯, 等. 宽皮柑橘种质资源表型多样性分析及综合评价[J]. 中国农业科学, 2017, 50(22): 4362-4372.
[11] 董勝君, 孙永强, 陈建华, 等.野杏无性系表型性状多样性分析及综合评价[J]. 植物遗传资源学报, 2020, 21(5): 1156-1166.
[12] 胡标林, 万 勇, 李 霞, 等. 水稻核心种质表型性状遗传多样性分析及综合评价[J]. 作物学报, 2012, 38(5): 829-839.
[13] 孙东雷, 卞能飞, 陈志德, 等. 花生种质资源表型性状的综合评价及指标筛选[J]. 植物遗传资源学报, 2018, 19(5): 865-874.
[14] 颉刚刚, 欧阳丽婷, 谢 军, 等. 新疆地区欧洲李叶片表型性状多样性及亲缘关系分析[J]. 植物资源与环境学报, 2018, 27(3): 72-78.
[15] 陈 巍, 王力荣, 朱更瑞, 等. 基于SSR标记和生物学性状进行桃遗传多样性的比较分析[J]. 植物遗传资源学报, 2009, 10(1): 86-90, 96.
[16] Campbell D R. Using phenotypic manipulations to study multivariate selection of floral trait associations[J]. Annals of Botany, 2009, 103(9): 1557-1566.
[17] 姜慧芳, 任小平, 张晓杰, 等. 中国花生小核心种质与ICRISAT微核心种质的SSR遗传多样性比较[J]. 作物学报, 2010, 36 (7): 1084-1091.
[18] 姜慧芳, 任小平, 廖伯寿, 等. 中国花生核心种质的建立及与ICRISAT花生微核心种质的比较[J]. 作物学报, 2008, 34(01): 25-30.
[19] 肖 宇, 陈剑洪, 郭陞垚, 等. 福建省近年来审(认)定花生品种的遗传多样性分析[J]. 植物遗传资源学报, 2015, 16(4): 728-733, 742.
[20] 江建华, 倪皖莉, 管叔琪, 等. 126份花生品种主要农艺性状遗传多样性分析[J]. 中国农学通报, 2014, 30(3): 117-123.
[21] 杨学乐, 张 璐, 李志清, 等.苦荞种质资源表型性状的遗传多样性分析[J]. 作物杂志, 2020(5): 53-58.
[22] 洪彦彬, 梁炫强, 陈小平, 等. 花生栽培种(Arachis hypogaea)类型间遗传差异的SSR分析[J]. 分子植物育种, 2008, 6(1): 71-78.
[23] 康红梅, 李保云, 孙 毅. 花生表型及SSR遗传多样性的研究[J]. 植物遗传资源学报, 2012, 13(1): 66-71, 76.
[24] 禹山林. 中国花生品种及其系谱[M]. 上海: 上海科学技术出版社, 2008.
[25] 代攀虹, 孙君灵, 何守朴, 等. 陆地棉核心种质表型性状遗传多样性分析及综合评价[J]. 中国农业科学, 2016, 49(19): 3694-3708.
[26] 王海岗, 贾冠清, 智 慧, 等. 谷子核心种质表型遗传多样性分析及综合评价[J]. 作物学报, 2016, 42(1): 19-30.
[27] 万述伟, 宋凤景, 郝俊杰, 等. 271份豌豆种质资源农艺性状遗传多样性分析[J]. 植物遗传资源学报, 2017, 18(1): 10-18.
[28] 甄志高, 段 莹, 王晓林, 等. 花生籽粒比重与生态特性的研究[J]. 花生学报, 2004, 33(1): 32-34.
[29] 赵 瑞, 许瀚卿, 樊冬丽, 等. 气候变化对中国花生生产的影响研究进展[J]. 中国农学通报, 2017, 33(21): 114-117.
[30] 万琼莲, 蔡 红, 杨子祥, 等. 云南花生丛枝植原体secY基因序列分析及结构预测[J]. 云南农业大学学报(自然科学版), 2013, 28(2): 163-168.
[31] 陈慕容, 张曙光, 郑冠标, 等. 华南花生丛枝病发病规律及防治研究[J]. 广东农业科学, 1994, 21(2): 33-35.
[32] 耿显胜, 舒金平, 王浩杰, 等. 植原体病害的传播、流行和防治研究进展[J]. 中国农学通报, 2015, 31(25): 164-170.
责任编辑:沈德发
猜你喜欢
长江蔬菜·学术版(2016年12期)2017-01-12
科学与财富(2016年26期)2016-12-01
医学信息(2016年29期)2016-11-28
大学教育(2016年11期)2016-11-16
企业导报(2016年9期)2016-05-26
