
剑麻EST-SSR在丝兰麻和中美麻中的通用性分析
2021-08-26 08:58张燕梅李俊峰鹿志伟杨子平周文钊
热带作物学报 2021年7期
张燕梅 李俊峰 鹿志伟 杨子平 周文钊

摘 要:采用聚丙烯酰胺凝胶电泳技术,对剑麻‘H.11648中的100对SSR引物在丝兰麻和万年兰中的通用性进行分析,同时构建DNA指纹图谱。结果表明100对SSR引物,在龙舌兰属、丝兰麻属、中美麻属中分别有68对、52对和52对SSR引物扩增出目标产物条带,扩增产物所占比例分别为68%、52%和52%,多态性引物分别为18对、12对和9對,多态性引物所占比例分别为29.51%、23.08%和17.31%。检测位点数分别为76、44和40个,平均每对引物多态位点数分别为2.67、2.42和2.22个。引物C67656和C61418可将龙舌兰属的6份种质区分开来,引物C65059可将丝兰麻属的5份种质区分开,引物C24110可将中美麻属的4份种质区分开。以上研究结果表明,从‘H.11648开发的SSR引物在龙舌兰及其近缘属中是可以通用的,筛选出的SSR引物可用于龙舌兰及其近缘属种质资源的鉴定和遗传多样性分析等研究。
关键词:剑麻;EST-SSR;丝兰麻;中美麻;通用性
中图分类号:S563.8 文献标识码:A
Transferability Analysis of Sisal EST-SSR Markers in Yucca and Furcraea vent
ZHANG Yanmei, LI Junfeng, LU Zhiwei, YANG Ziping, ZHOU Wenzhao*
South Subtropical Crops Research Institute, Chinese Academy of Tropical Agricultural Sciences / Zhanjiang City Key Laboratory for Tropical Crops Genetic Improvement, Zhanjiang, Guangdong 524091, China
Abstract: In this study, polyacrylamide gel electrophoresis was used to analyze the generality of 100 pairs of SSR primers from ‘H.11648 in Agave and its related genus, and DNA fingerprinting was constructed. The results showed that 68, 52 and 52 pairs of SSR primers could amplify the expected products in Agave, Yucca and Furcraea vent, respectively. The proportion of amplified products was 68%, 52% and 52%. 18, 12 and 9 pairs of SSR primers showed polymorphic and the polymorphic percentage was 29.51%, 23.08% and 17.31%. The total number of polymorphic loci was 76, 44 and 40 with average number of polymorphic loci amplified by each primer was 2.67, 2.42 and 2.22, respectively. Primers C67656 and C61418 could discriminate six germplasms in Agave, Primer C65059 could discriminate five germplasms in Yucca, Primer C24110 could discriminate four germplasms in Furcraea vent. The above results indicated that the SSR primer in ‘H.11648 was transferable in Agave and its related genera, which laid a foundation for germplasm identification and genetic diversity analysis of Agave and its related genera.
Keywords: sisal; EST-SSR; Yucca; Furcraea vent; transferability
DOI: 10.3969/j.issn.1000-2561.2021.07.003
剑麻属龙舌兰科(Agavaceae)多年生单子叶植物,全世界有20个属约670种,主要分布在墨西哥等热带、亚热带地区[1],我国现保存有7个属约100余份种质资源,其中龙舌兰属植物较多,作为一种重要的热带经济作物,除生产纤维外,剑麻茎心可用来酿酒[2],麻渣可做饲料、制药以及肥料等[3],剑麻亦可作为园林植物,有重要的观赏价值。
微卫星DNA(microsatellite DNA),又称简单重复序列(simple sequence repeat, SSR),是由1~6个核苷酸为重复单位串联而成的长达几十个核苷酸的重复序列,广泛分布于整个基因组中,按其来源可分为基因组SSR(gSSR)和表达序列标签SSR(EST-SSR)。与其他分子标记相比,SSR标记具有多态性高、重复性好、共显性遗传等优点[4],可以区分纯合子和杂合子,广泛应用于遗传分析[5]、种质资源鉴定[6]以及遗传作图[7]和QTL分析[8]等领域。关于SSR引物在不同植物中的通用性研究报道较多,王瑞晶等[9]利用小麦EST- SSR对47份偃麦草进行遗传多样性分析,通用性达到87.91%。文明富等[10]研究表明,木薯EST- SSR在麻风树和橡胶树中的通用性分别为37.36%和26.37%。随着测序技术和生物信息技术的迅猛发展,全球公共数据平台进一步开放共享,尤其是对于非模式植物和基因组信息未知的物种提供了良好的机遇,既可以缩短研发时间,又能节约大量的人力和财力。目前有关龙舌兰科中报道的SSR引物仅有7对,来自于Agave parryi[11],主要用于龙舌兰属植物遗传多样性研究[11-13],SSR引物偏少,获得的信息量有限,而且也没有关于剑麻SSR通用性研究的报道,张燕梅等[14]利用
‘H.11648转录组EST序列,开发100对SSR引物,其中有18对引物在龙舌兰属中表现较好的多态性。鉴于此,本研究拟利用从剑麻中开发的100对SSR引物,在丝兰麻和中美麻中进行通用性研究,既可以节约开发成本,同时为龙舌兰及其近缘属的资源鉴定、遗传多样性分析以及比较作图奠定基础。
1 材料與方法
1.1 材料
1.1.1 试验材料 选用丝兰麻属的5份种质,中美麻属的4份种质,龙舌兰属的6份种质,共15份材料,每份材料取2年生吸芽3~5株,每株取1 g叶片等量混合后用于提取基因组DNA。所有材料均保存于中国热带农业科学院南亚热带作物研究所剑麻种质资源圃(表1)。
1.1.2 SSR引物 本研究所用100对EST-SSR引物为本团队从‘H.11648转录组EST序列中开发的SSR引物,具体见张燕梅等[14]报道的文献。引物由生工生物工程(上海)股份有限公司合成。
1.2 方法
1.2.1 DNA提取 DNA提取采用天泽公司的柱式植物DNAout试剂盒提取,试验材料用液氮快速研磨后取少量粉末,加入裂解液、离心后取上清,上清液经抽提、漂洗最后过柱,加入100 μL洗脱液将DNA从吸附柱上洗脱下来,4 ℃保存备用,具体操作参照试剂盒说明书。
1.2.2 SSR引物通用性分析 先将每个属的所有DNA样本等量混合后构建DNA池,以每个属的DNA池为模板进行PCR扩增,初步筛选有扩增产物的SSR引物。然后分别以单个样本的DNA为模板,对有扩增产物的SSR引物进行多态性筛选。
PCR反应体系。20 ?L PCR反应体系中各组份的浓度及使用量:10×Buffer(全式金):2 ?L;dNTP:1.6 ?L;Easy-Taq(5 U/?L):0.2 ?L;引物(10 μmol/L)F:0.5 ?L,R:0.5 ?L;DNA:50 ng(1 ?L);灭菌ddH2O:14.2 ?L。所有试剂购自北京全式金生物技术有限公司的Easy Taq DNA polymerase for PAGE kit。
扩增程序。采用Touch-down PCR程序,具体为95 ℃ 15 s,60 ℃ 15 s(=-0.7,即每增加一个循环,退火温度降低0.7 ℃),72 ℃ 30s,16个循环;然后进入下一个扩增阶段,95 ℃ 15 s,50 ℃ 15 s,72 ℃ 30 s,15个循环;最后72 ℃ 60 min,扩增产物4 ℃保存备用。
PCR产物检测。PCR产物采用8%非变性聚丙烯酰胺凝胶(丙烯酰胺∶双丙烯酰胺=29∶1)电泳进行分离,具体参考史卫东等[15]的方法并略有调整。20 ?L PCR反应体系加入8 ?L 6loading buffer,混匀后2.5 ?L上样,250 V,电泳3 h后取下胶板,用双蒸水冲洗后,加入固定液(10 mL无水乙醇+5 mL冰乙酸,然后用水补足至100 mL)固定7 min,然后加入AgNO3染色液(0.15% AgNO3)银染10 min,最后加入四硼酸钠显影液(100 mL H2O+1.5 g NaOH+0.756 g四硼酸钠+ 0.75 mL甲醛)显色,直到显出清晰的条带,自来水冲洗后即可拍照。以上操作均在摇床上进行。
1.3 数据处理
采用人工判读数据,某一位点有产物记为“1”,没有产物记为“0”,缺失记为“.”,所得数据输入Excel表格后,绘制每个样本的DNA指纹。
2 结果与分析
2.1 剑麻SSR标记的通用性分析
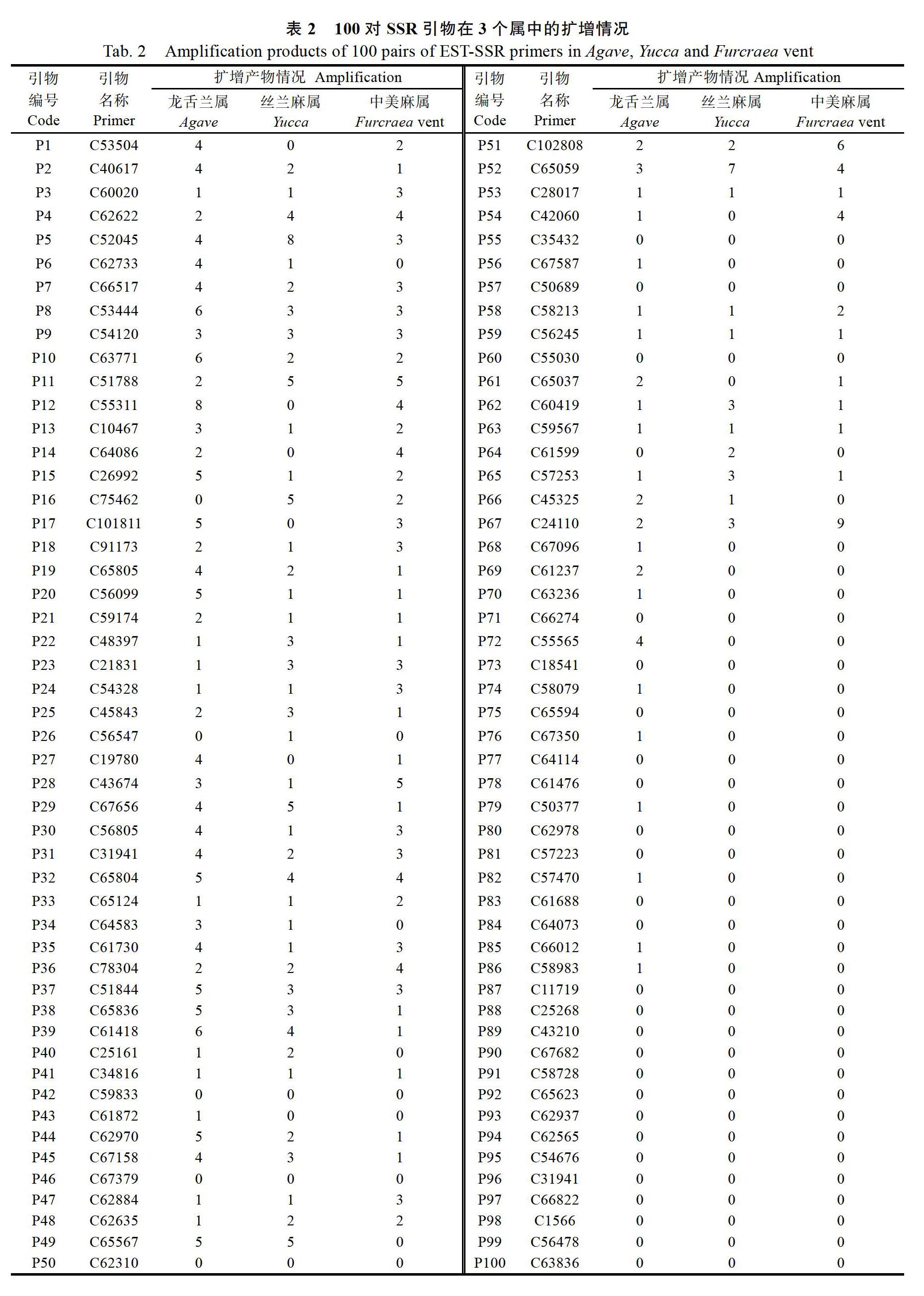
采用聚丙烯酰胺凝胶电泳对100对剑麻SSR引物PCR扩增产物进行检测,有45对SSR引物在3个属中均可扩增出目标产物条带,有68对SSR引物在龙舌兰属中可扩增出目标产物条带,扩增产物所占比例为68%。有52对SSR引物在丝兰麻属中可扩增出目标产物条带,扩增产物所占比例为52%。有52对SSR引物在中美麻属中扩增出目标产物条带,扩增产物所占比例为52%。从而表明从‘H.11648中开发的SSR引物在龙舌兰、丝兰麻以及中美麻等近缘属中是可以通用的,且引物通用性在52%~68%之间(表2)。
2.2 SSR引物多态性分析
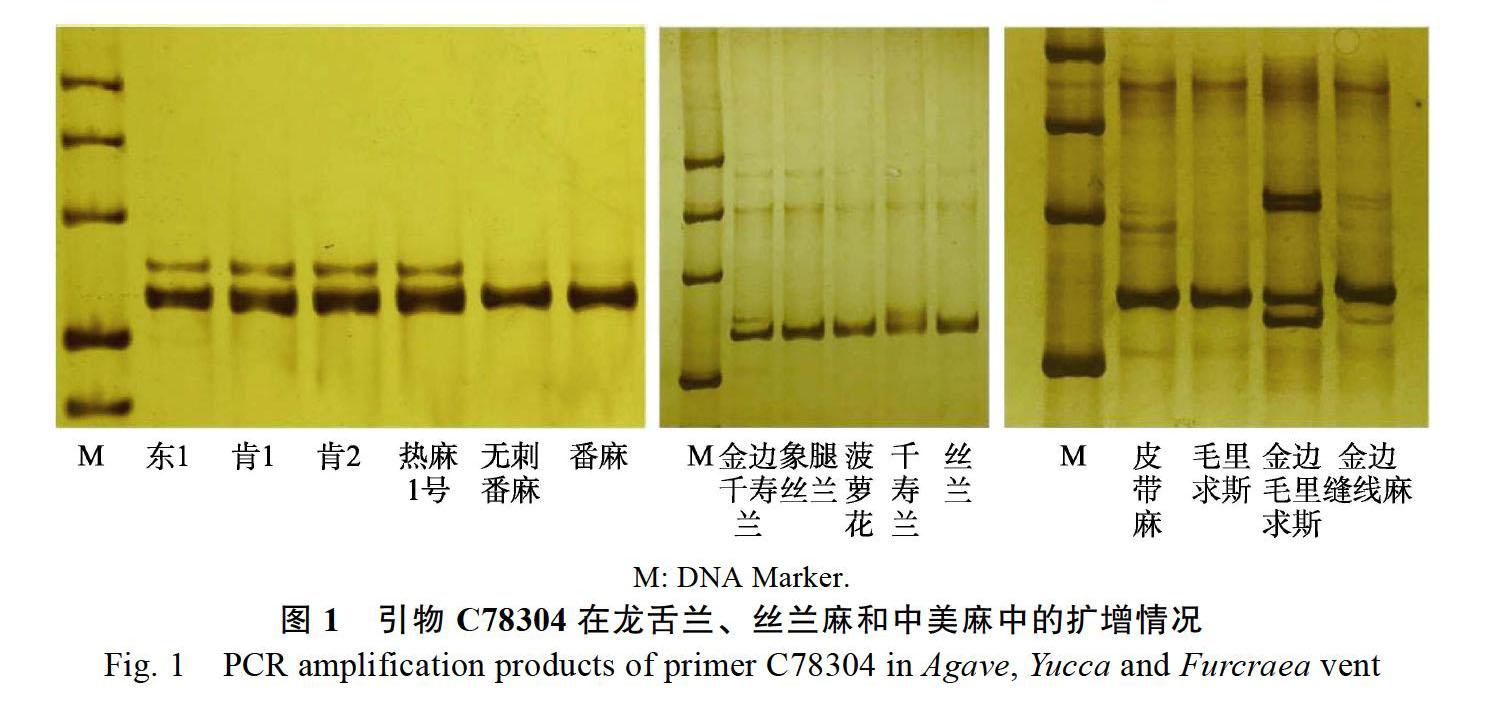
对扩增有目标产物条带的SSR引物进行多态性分析,龙舌兰属68对SSR引物中,有18对引物在龙舌兰属6个个体中表现多态性,多态性引物所占比例为29.51%,18对SSR引物可检测到76个SSR位点,其中每对引物平均检测到4.22个位点,平均多态性位点数为2.67,多态百分比为63.16%(表3)。丝兰麻属52对SSR引物中,有12对引物在丝兰麻属5个个体中表现多态性,多态性引物所占比例为23.08%,12对SSR引物可检测到44个SSR位点,其中每对引物平均检测到3.67个位点,多态百分比为65.91%(表4)。中美麻属52对SSR引物中,有9对引物在中美麻属4个个体中表现多态性,多态性引物所占比例为17.31%,9对SSR引物可检测到40个SSR位点,其中每对引物平均检测到4.44个位点,平均多态性位点数为2.22,多态百分比为50%(表5)。只有引物C78304在龙舌兰、丝兰麻和中美麻中同时表现出多态性(图1)。
2.3 DNA指纹图谱构建
SSR标记在指纹图谱构建中的效率与其在研究对象中的多态性密切相关,一般多态性高的标记效率高。龙舌兰属18对多态性引物分别可检测2~6个位点,83.33%的引物检出位点在4个以上,引物C67656和C61418可将龙舌兰属的6份种质区分开。丝兰麻属12对多态性引物分别可检测2~5个位点,33.33%的引物检测位点数在4个以上,其中引物C51788、C75462和C75462可同时
将4份种质区分开,引物C65059可将5份种质区分开。中美麻属有8对多态性引物可检出4个以上位点,多态性位点数在1~5个之间,引物C24110可将4份种质区分开。同时根据分析结果绘制15份种质的SSR-DNA指纹图谱(图2)。
3 讨论
近年来,利用植物EST序列开发EST-SSR引物,并在近缘或远缘物种中进行通用性分析已成为一种较为常见的引物开发手段,无论在单子叶植物还是双子叶植物中均有许多成功的报道。本研究对来自于‘H.11648的100对EST-SSR引物分别在龙舌兰属、丝兰麻属和中美麻属中的通用性进行分析显示‘H.11648EST-SSR引物通用性在52%~68%之间,多态性引物比例在17.31%~29.15%之间,表明剑麻‘H.11648EST-SSR引物在龙舌兰及其近缘属中的通用性较高。这与Holton等[16]报道大麦EST-SSR在小麦中的通用性为55%,Malay等[17]研究显示高羊茅EST-SSR对其他草类通用性为57%,徐磊等[18]报道高粱SSR在割手密中的通用性为70%基本一致。此外,研究也发现利用‘H.11648EST-SSR开发的SSR引物,在龙舌兰及其近缘属的通用性和多态性引物比例明显比Lindsay等[11]报道的基因组SSR通用性高。引物通用性高低既与SSR侧翼序列的保守程度和SSR基序本身在进化过程中的稳定性有关[19],同时也与SSR标记来源有关。
如Xu等[20]发现北美鹅掌楸SSR标记在中国鹅掌楸中的通用性达到85%。此外,郑丽珊等[21]研究发现,双子叶植物棉花的SSR可在单子叶植物香蕉中通用,且EST-SSR的通用性比基因组SSR高。同样地,杨彦伶等[22]研究也表明杨树的EST-SSR在柳树中的通用性比基因组SSR高,与本研究结果一致。
SSR标记具有丰富的多态性,利用该标记,可较好地区分不同品种资源[23]。如引物C65059可较好地将丝兰麻的5份种质区分开。但研究发现,‘肯1和‘肯2几乎有相同的指纹,‘肯1和‘肯2均来自肯尼亚,根据表型很难将二者分开,说明这2份种质亲缘关系较近,存在几乎相同的指纹,一方面可能存在重复引种,另一方面,可适当增加标记数量来进一步检测。此外,尽管100对引物中有43对在龙舌兰属、丝兰麻属和中美麻属中都有扩增,但最后只检测到1对引物在这3个属中同时具有多态性,究其原因,一方面由于每个属的个体数较少,SSR数量不多,若个体间亲缘关系近则会使多态性引物减少,另一方面在数据分析时,龙舌兰属、丝蘭麻属和中美麻属是分开分析的,从而导致在某一个或两个属中有多态性的引物未被列入其中,因此可以适当增加引物数量,优化反应体系,从而达到更好的检测效果。
此外,龙舌兰、丝兰麻和中美麻同属龙舌兰科植物,近缘关系较近,通过本研究开发出一套通用的EST-SSR标记,不仅可以有效弥补丝兰麻和中美麻中分子标记不足的现状,丰富标记数量,减少标记开发成本和时间,同时通过不同物种间的比较作图,揭示其进化关系,为遗传育种研究奠定基础。
参考文献
[1] 庄雪影. 园林树木学(华南本)[M]. 广州: 华南理工大学出版社, 2014: 289.
[2] Escalante A, López Soto D R, Velázquez Gutiérrez J E, et al. Pulque, a traditional Mexican alcoholic fermented beverage: historical, microbiological, and technical aspects[J]. Frontiers in Microbiology, 2016, 7: 1026.
[3] Pereira G M, Ribeiro M G, da Silva B P, et al. Structural characterization of a new steroidal saponin from Agave angustifolia var. Marginata and a preliminary investigation of its in vivo antiulcerogenic activity and in vitro membrane permeability property[J]. Bioorganic & Medicinal Chemistry Letters, 2017, 27(18): 4345-4349.
[4] Mason A S. SSR Genotyping[M]// Batley J. Plant Geno-typing. Springer, New York, NY, 2015: 77-89.
[5] Bibi A C, Gonias E D, Doulis A G. Genetic diversity and structure analysis assessed by SSR markers in a large collection of vitis cultivars from the island of Crete, Greece[J]. Biochemical Genetics, 2020, 58(2): 294-321.
[6] Zhao Y, Wang Y, Wang L, et al. Molecular identification of mung bean accessions (Vigna radiate L.) from Northeast China using capillary electrophoresis with fluorescence- labeled SSR markers[J/OL]. Food and Energy Security, 2019, 9(20), DOI:10.1002/fes3.182.
[7] Wang Y, Jia H M, Shen Y T, et al. Construction of an anchoring SSR marker genetic linkage map and detection of a sex-linked region in two dioecious populations of red bayberry[J]. Horticulture Research, 2020, 7(1): 53.
[8] Daware A, Das S, Srivastava R, et al. An efficient strategy combining SSR markers- and advanced QTL-seq-driven QTL mapping unravels candidate genes regulating grain weight in rice[J]. Frontiers in Plant Science, 2016, 7: 1535.
[9] 王瑞晶, 李培英. 小麦EST-SSR标记在偃麦草中的通用性分析[J]. 分子植物育种, 2016,14(6): 1516-1523.
[10] 文明富, 陈 新, 王海燕, 等. 木薯基因组SSR和EST-SSR在麻疯树和橡胶树中的通用性分析[J]. 作物学报, 2011, 37(1): 74-78.
[11] Lindsay D L, Edwards C E, Jung M G, et al. Novel microsatellite loci for Agave parryi and cross-amplification in Agave palmeri (Agavaceae)[J]. American Journal of Botany, 2012, 99(7): e295-e297.
[12] Cabrera-Toledo D, Vargas-Ponce O, Ascenclo-Ramírez S, et al. Morphological and genetic variation in monocultures, forestry systems and wild populations of Agave maximiliana of western Mexico: Implications for its conservation[J]. Frontiers in Plant Science, 2020, 11: 817.
[13] Trejo L, Limones V, Pe?a G, et al. Genetic variation and relationships among agaves related to the production of Tequila and Mezcal in Jalisco[J]. Industrial Crop & Products, 2018, 125: 140-149.
[14] 張燕梅, 李俊峰, 杨子平, 等. 基于转录组的剑麻SSR标记开发与鉴定[J]. 热带作物学报, 2021, 42(5): 1261-1266.
[15] 史卫东, 罗海玲, 康红卫, 等. 基于非变性聚丙烯酰胺凝胶电泳的菜心SCoT分析[J]. 亚热带植物科学, 2020, 49(1): 1-8.
[16] Holton T A, Christopher J T, McClure L, et al. Identifi-cation and mapping of polymorphic SSR markers from ex-pressed gene sequences of barley and wheat[J]. Mo-lecular Breeding, 2002, 9(2): 63-71.
[17] Saha M C, Mian R, Zwonitzer J C, et al. An SSR- and AFLP- based genetic linkage map of tall fescue (Festuca arundinacea Schreb.)[J]. Theoretical and Applied Genetics, 2005, 110(2): 323-336.
[18] 徐 磊, 邢淑莲, 姚艳丽, 等. 高粱SSR和EST-SSR标记在割手密中的通用性分析[J]. 中国农学通报, 2015, 31(27): 164-171.
[19] 程小毛, 黄晓霞. SSR标记开发及其在植物中的应用[J]. 中国农学通报, 2011, 27(5): 304-307.
[20] 胥 猛, 李火根. 鹅掌楸EST-SSR引物开发及通用性分析[J]. 分子植物育种, 2008(3): 615-618.
[21] 郑丽珊, 石玉真, 王静毅, 等. 棉花EST-SSRs在香蕉中的通用性[J]. 中国农学通报, 2008, 24(1): 33-37.
[22] 杨彦伶, 张亚东, 张新叶. 杨树SSR标记在柳树中的通用性分析[J]. 分子植物育种, 2008, 6(6): 1134-1138.
[23] 王源秀, 徐立安, 黄敏仁. 杞柳和簸箕柳候选杂交亲本SSR指纹图谱分析[J]. 南京林业大学学报(自然科学版), 2008, 32(2): 1-5.
责任编辑:谢龙莲
