
侵蚀-沉积作用对黄土高原土壤中固碳细菌群落分布特征的影响
2021-08-26 09:53彭诗阳钟仁华吴岸松蔡红春张燕茹
湖南林业科技 2021年4期
彭诗阳, 钟仁华, 吴岸松, 蔡红春, 何 勇, 张燕茹
(1.湖南鑫恒环境科技有限公司, 湖南 长沙 410000; 2.湖南鑫恒生态研究院有限公司, 湖南 益阳 413002;3.湖南省林业科学院, 湖南 长沙 410004)
中国西北部的黄土高原区是世界上土壤侵蚀程度最严重的区域。由于该区的人类活动密集,以及降雨集中且量大,导致其土壤可侵蚀性高,致使其中60%以上的土地遭受侵蚀[1]。土壤侵蚀不仅对生态环境和粮食安全造成威胁,还会影响碳循环过程。相关研究主要集中于侵蚀对土壤有机碳流失以及再分布的影响,对于与土壤碳循环紧密相关的土壤固碳微生物在侵蚀影响下的变化规律却报道甚少。土壤固碳是大部分干旱土壤的主要碳源[2],然而,土壤微生物在土壤固碳中的作用往往被忽视。自养细菌可以利用大气中的CO2进行正常的代谢活动,并直接参与大气中CO2的同化[3]。Calvin-Benson-Basham(CBB)循环是主要的CO2固定途径[4],核酮糖-1,5-二磷酸羧化酶/加氧酶(RuBisCO)是调节CBB循环中光合作用的关键酶[5],编码I型RuBisCO大亚基的cbbL基因则是作为分析自养微生物的功能基因[6]。近年来,基于RuBisCO编码cbbL基因的固碳分子生态学研究主要集中在特定的生态环境上。有研究发现,高海拔地区的贫瘠土壤中往往含有光自养细菌群落,其多样性高于其他地区的[2]。也有学者从DNA水平揭示了黄土高原生态系统固碳细菌的群落结构和多样性,并利用测序技术发现了许多新型固碳细菌基因[7]。目前,对黄土高原土壤中CO2同化细菌与非典型土壤条件之间关系,以及土壤微生物,特别是自养细菌对土壤侵蚀响应的研究报道甚少。本研究分析了不同侵蚀-沉积坡面土壤固碳细菌的群落分布特征,以及土壤理化参数与携带cbbL基因细菌群落的关系,以期为土壤侵蚀诱发碳循环微生物机制的研究提供参考。
1 研究区概况
以甘肃省天水市水土保持试验站内罗峪沟流域的主要分支—桥子沟流域(105°30′—105°45′E,105°30′—105°45′N,海拔1 438 m)为研究区域。该流域属于黄土高原丘陵沟壑区第三副区,属于温带季风气候区,以半干旱气候为主要特征。其年平均气温约为10.7 ℃,年平均降水量为 496~628 mm,降雨一般集中在7—9月[8]。分布最广的土壤类型为黑褐土和黄绵土,占整个流域面积的 82%以上,其土壤质地呈粉质。降雨是造成该地区土壤侵蚀的主要自然驱动力,其土壤平均侵蚀率为1 599.50~2 877.76 t· km-2a-1,年产沙量为2 026 mg·km-2[9]。
2 研究方法
2.1 试验设计与样品采集
分别选择位于桥子西小流域并种植有刺槐的坡地(面积0.23 km2)截面和位于桥子东小流域的未开垦坡地(面积0.21 km2)截面各一个。2个坡地相隔1.65 km,2个坡面均呈“S”型,且平均坡度为 12°。将每个截面分为侵蚀区(上坡)和沉积区(下坡)。分别于未开垦坡面的侵蚀点(UE)、未开垦坡面的沉积点(UD),刺槐坡面的侵蚀点(RE)和刺槐坡面的沉积点(RD)共4个采样点设置土壤侵蚀与沉积处理。为了采集表层(0~10 cm)土样,首先清除石块、生物结皮以及表层土壤约1 cm的可见植物残留物,然后在4个采样点分别设置3个2 cm×2 cm的样方,用内径为 5 cm 的土壤螺旋钻在每个样方中随机采集 5~9 个土样并将土样混合,每种处理重复 3次。采集的土壤样本立刻存于自封袋中低温运输至实验室,然后将一部分样品冷冻干燥(温度为-80 ℃)并保存在50 ml无菌离心管中,用于分子生物学分析;剩余的土壤样品自然风干并过0.25 mm的筛,用于测量土壤理化性质。
2.2 土壤理化性质测定
采用烘干法测定含水率。将新鲜土样中的石头、根系去除,风干并过2 mm筛后,用电极仪测定土壤pH值。采用吸管法测定土壤机械组成。采用油浴重铬酸钾外加热法测定有机碳(SOC)含量。用总有机碳分析仪(TOC.VCPH,日本岛津)测定土壤溶解性有机碳(DOC)含量。土样过0.25 mm 筛后,分别采用凯氏定氮法、氢氧化钠熔融比色法和氢氧化钠熔融火焰光度法测定总氮(TN)、总磷(TP)和总钾(TK)。分别采用碱水解扩散法、Mo-Sb比色法以及火焰光度法分别测定土壤速效氮(AN)、速效磷(OP)和速效钾(AK)。用超纯锂漂移锗探测器和DSPEC多通道γ射线分光光度计(GMX50,PerkinElmer,美国)测量137Cs活性,平均计数时间超过20000s,最后用661.6kev的峰面积计算137Cs含量。
2.3 DNA提取和PCR扩增
使用E.Z.N.A.土壤DNA试剂盒(Omega Bio-tek,Norcross,GA,美国)提取土壤微生物基因组总 DNA,按照试剂盒说明书进行操作。提取物保存于-20 ℃冰箱中备用。
利用cbbL引物(K2f-V2r)扩增自养细菌,其中K2f引物组成为5′-ACCAYCAAG CCSAAGCTSGG-3′ ,V2r引物组成为5′-GCCTTCSAGCTCT CTCACCRC-3′[10]。对cbbL基因进行PCR扩增[11]。PCR反应条件为:先在95 ℃下3 min,然后在95 ℃下35个周期30 s,55 ℃下30 s,72 ℃下45 s,最后在72 ℃下延伸10 min。PCR反应体系为:4 μL 5×快速Pfu缓冲液、2 μL 2.5 mM脱氧核苷酸三磷酸(dNTPs)、每个引物(5 μM)0.8 μL、0.4 μL快速Pfu DNA聚合酶和10 ng模板DNA,加无菌水使反应总体系为 20 μL,每个样品设置3组平行试验。使用ABI GeneAmp®9700(美国)进行PCR扩增。PCR 产物进行琼脂糖电泳,并对 DNA 进行切胶回收,使用QuantiFluor(Promega,美国)检测试剂盒对回收的 DNA 精确定量,等量混合后测序。
2.4 Illumina MiSeq测序和数据处理
在Illumina MiSeq平台上进行配对末端测序(2×250)。测序步骤大致如下:①根据 barcode 序列将各样本区分开并去除 barcode;②使用QIIME(版本1.91)对原始的FASTQ文件进行解复用和质量过滤; ③利用Uclust软件根据序列之间的相似度(97%)将cbbL基因序列归为不同的操作分类单元(OTU)[12]; ④使用BLAST程序[13],对照GeneBank(国家生物技术信息数据库中心)中已知的cbbL序列,对序列进行分类鉴定,得到自养细菌的分类学信息。所有原始序列都保存在GenBank中,数据编号为PRJNA350577。
利用mothur软件计算土壤细菌 α 多样性指数,包括Chao1丰富度指数、ACE丰富度指数、Shannon多样性指数以及Simpson多样性指数。通过群落生态学软件包R-forge进行主坐标分析(PCoA)。采用 SPSS 18.0 进行方差分析(Duncan 检验法)和主成分分析,用Origin 软件(8.5版本)作图。差异显著性水平设为α=0.05。
3 结果与分析
3.1 土壤理化性质及土壤养分含量
表1结果显示:4个样点的土壤质地相似,均以粉粒为主,其次是砂粒,最后是黏粒。在2个侵蚀坡面中,沉积区的粉粒含量均高于侵蚀区的。4个样点的土壤含水率和容重均无显著差异。未开垦坡面的土壤pH值虽整体高于刺槐坡面的,但是侵蚀-沉积扰动没有对未开垦坡面的pH值造成影响,UE和UD点的土壤pH值无显著差异。刺槐坡面上,侵蚀区的土壤pH值显著高于沉积区的。

表1 四个样点土壤的理化性质Tab.1 Physical and chemical properties of soil of four sample sites样点砂粒/%粉粒/%黏粒/%含水率/%容重/(g·cm-3)pH值UE35.26±0.96 ab43.71±0.79 a21.03±0.32 c13.20±0.20 a1.17±0.07 a8.69±0.03 aUD36.00±0.39 a43.98±0.08 ab20.02±0.37 c13.17±0.48 a1.28±0.03 a8.71±0.02 aRE30.39±0.85 b44.27±0.22 c25.34±0.64 a13.04±1.39 a1.26±0.04 a8.44±0.02 bRD32.35±1.18 c44.53±0.71 d23.12±0.48 b12.07±0.67 a1.22±0.16 a8.35±0.04 c注: UE为未开垦坡面的侵蚀点;UD为未开垦坡面的沉积点;RE为刺槐坡面的侵蚀点;RD为刺槐坡面的沉积点。
不同样点土壤养分含量结果(表2)显示:UE的土壤有机碳含量显著低于UD的,RE的土壤有机碳含量显著低于RD的。说明侵蚀扰动对土壤有机碳含量有明显影响。UD的可溶性有机碳含量显著高于UE的;RD的可溶性有机碳含量显著高于RE的。由此可见,侵蚀-沉积扰动对于土壤中碳含量的影响也明显。RE的TP含量略低于RD的, UE的TP含量与UD的无明显差异;UD的 OP含量显著低于UE的,UD的AK含量显著高于UE的;TP和OP的含量在RE和RD间无明显差异。同一坡面的侵蚀区和沉积区的TN、TK或AN含量均无显著差异。

表2 四个样点土壤的养分含量Tab.2 Nutrient content of four sample sites样点TN/(g ·kg-1)TP/(g · kg-1)TK/(g ·kg-1)AN/(mg · kg-1)OP/(mg ·kg-1)AK/(mg ·kg-1)SOC/(g · kg-1)DOC/(mg ·g-1)UE1.00±0.02 a0.92±0.02 a19.21±0.06 a51.07±1.05 a4.80±0.00 a211.35±0.87 b7.29±0.72 b36.38±0.71 aUD1.05±0.04 a0.91±0.01 a19.33±0.03 a55.19±1.57 a3.77±0.01 b259.41±7.06 a7.91±0.88 a38.62±1.86 bRE1.07±0.03 a0.82±0.00 b19.79±1.01 a62.49±2.63 a3.76±0.00 b262.00±1.99 a8.10±0.57 a36.35±1.41 aRD1.06±0.01 a0.84±0.03 b19.70±1.63 a58.14±0.48 a3.75±0.00 b169.96±3.97 c8.94±0.00 c37.15±2.19 c
3.2 土壤自养细菌的多样性和丰度
表3结果显示:从4个样点土壤样品中共提取了63 291个自养细菌cbbL基因的有效序列,同时,在UE、UD、RE和RD样点的土壤样品中分别检测到349、337、419和372个自养细菌OTU。4个样点的cbbL基因文库有效序列为13 344~18 888条,Reads(测序读长)个数为9 141~11 406条。覆盖率是指对各类文本库的覆盖率,其值越大,在样本中测量序列的概率就越大。表3结果中所有样品的覆盖率均达到99%以上,表明其序列的结果准确地描述了样品中微生物的情况。
表3结果还表明:4个样点土壤自养细菌的ACE无明显差异;以Chao1指数为尺度,2个坡面侵蚀区土壤自养细菌的丰富度均明显低于沉积区的。以Shannon、Simpson指数为尺度,侵蚀区土壤自养细菌的多样性均显著低于沉积区的。以上结果说明,侵蚀扰动对自养细菌的丰富度和多样性均有消减作用。

表3 四个样点土壤自养细菌测序结果以及α多样性指数Tab.3 Sequencing results of four sample sites and α diversity index样点有效序列读长OTU丰富度指数多样性指数ACEChao1ShannonSimpson覆盖率/%UE16 84511 314349377 ba387 b 4.41 c0.030 6 b99.60UD13 3449 141337376 ba391 ab4.63 b0.020 5 c99.40RE18 88811 406419436 a 411 a 4.27 a0.041 1 a99.70RD14 21410 102372407 ba438 ab4.74 d0.029 7 b99.43
3.3 土壤自养细菌群落在门、属分类学水平上的构成
将系统发育分析得到的cbbL基因序列分为不同的系统发育类群,探讨微生物群落的动态变化。在门分类学水平上的细菌群落相对总丰度如图1所示。图1显示,所有样点土壤中的自养细菌主要隶属于3个细菌门,即变形菌门(Proteobacteria)、放线菌门(Actinobacteria)和芽单胞菌门(Gemmatimonadetes)。这也是本研究中鉴定出的已知仅有的3个主要自养细菌门,分别占cbbL基因总序列的7.8%~10.9%、0.72%~1.5%和0.11%~1.42%。约有10%的自养细菌群落尚未划分为门级,有80%以上的自养细菌在环境样品中被鉴定为无等级,意味着这些自养细菌群落在门的层次上没有明确的分类信息和名称。
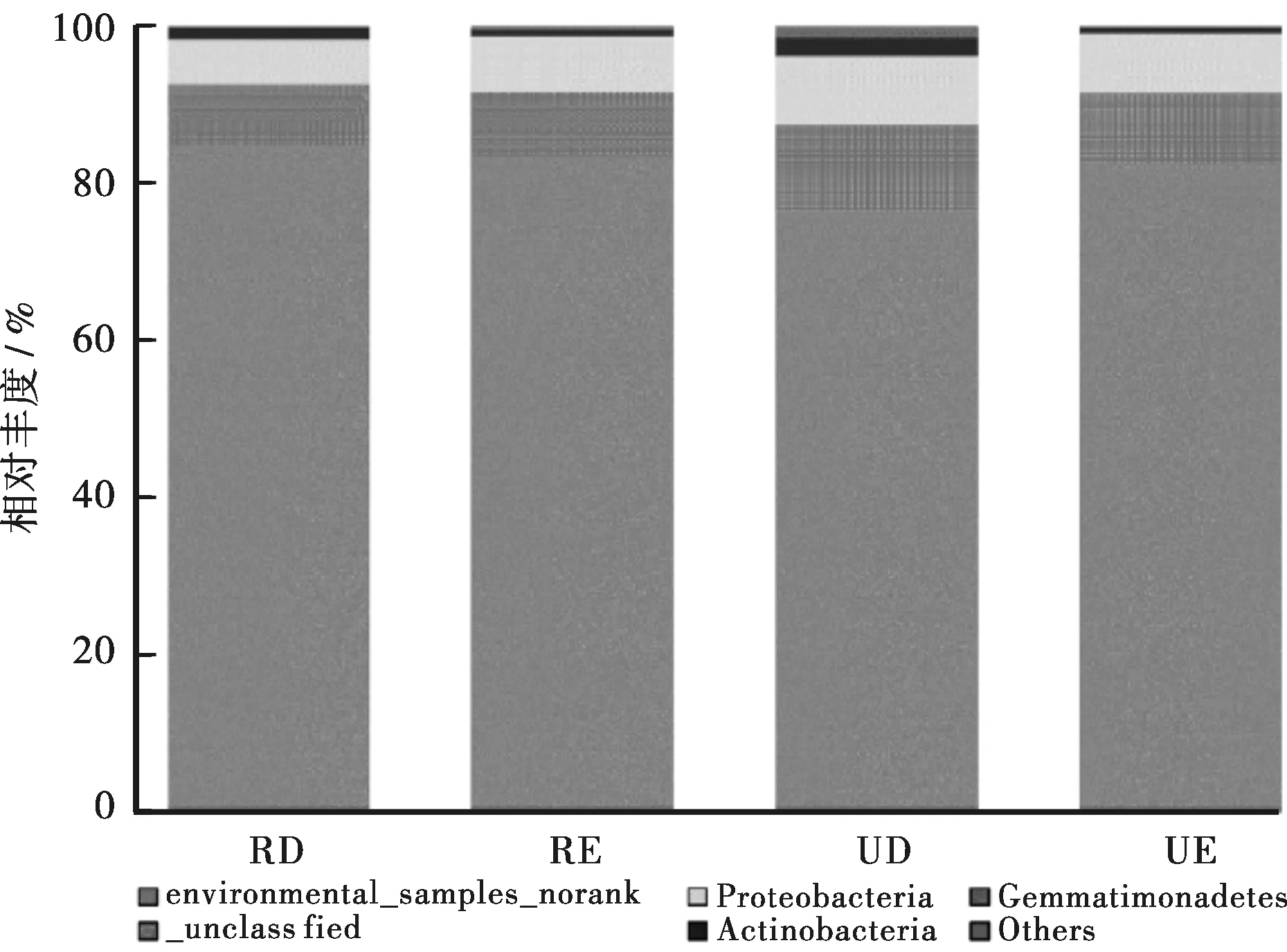
图1 四个样点土壤自养细菌在门分类学水平上的物种相对丰度Fig.1 The relative abundance of autotrophic bacteria at the phylum taxonomic level of the four sample sites
在属分类学水平下自养细菌群落的相对丰度见图2。图2显示:在所有土壤样品中,共鉴定出21个携带cbbL固碳功能基因的自养细菌属,其中斯塔克亚菌属(Starkeya)为优势自养细菌属,平均占所有土壤样品的3.68%;多态菌属(Polymorphum)是第二优势属,在UD和RD样品中分别占1.64%和1.3%。变形杆菌属(Ramlibacter)和贪噬菌属(Variovorax)分别为UE和RE的第二优势属,分别占2.19%、0.96%。值得关注的是,4个样点中,只有在UE样点发现了雷氏菌属(Ralstonia)和红假单胞菌属(Rhodopseudomonas)。

图2 四个样点土壤自养细菌在属分类学水平上的物种相对丰度Fig.2 The relative abundance of autotrophic bacteria at the genus taxonomic level of the four sample sites
从cbbL属水平测序得到的OTU中,82.37%属于no_rank,无法被鉴定。从4个样点土壤中获得的自养细菌克隆序列,有一些是与碳氮循环相关的功能细菌,例如具有光营养、固氮(蓝藻)、硝化和共氧化功能的细菌(Ralstoniaeutropha)。此外,本研究鉴定的细菌属多为兼性自养细菌,包括芽生殖菌属(Gemmatirosa,丰度范围为0.11%~1.42%)、贪铜菌属(Cupriavidus,丰度范围为0.04%~0.11%)和富养产碱菌属(Ralstoniaeutropha,丰度范围为0.001%~0.009%)。
3.4 自养细菌群落的结构
基于定量非加权UniFrac度量的PCoA揭示了4个样点土壤自养细菌群落结构的差异。图3结果表明:RE和RD样点的自养细菌群落在系统发育上比UE和UD样点的类群更接近,即在未开垦坡面上,沉积区和侵蚀区的自养细菌群落结构差异更为明显。第一轴显示细菌群落构成的累积变化量为60.62%,第二轴显示的为26.97%,横纵2个PCoA轴显示了87.59%的总变异。在同一坡面上,相似性的MANOVA分析表明,自养细菌群落在侵蚀部位和沉积部位存在差异(P>0.05)。

图3 基于Unifrac距离对四个样点土壤自养细菌群落结构的主坐标分析(PCoA)Fig.3 Principal Coordinate Analysis (PCoA) of soil autotrophic bacteria community structure at four sample sites based on unifrac distance
基于cbbL基因型属分类学水平上细菌群落剖面聚类热图和多个样本的相似性树分析揭示了同一侵蚀坡面上的自养细菌群落具有更高的相似性(见图4a)。图4b结果表明,根据自养细菌群结构的相似性,4个菌群可分为2类,一类是UE和UD菌群,另一类是RE和RD菌群,且这2类菌群之间也有很好的分离,表明2个坡面的自养细菌群落结构存在明显的差异。UE样点的土壤自养细菌群落结构和UD的相似,RE样地的土壤自养细菌群落结构与RD的相似。
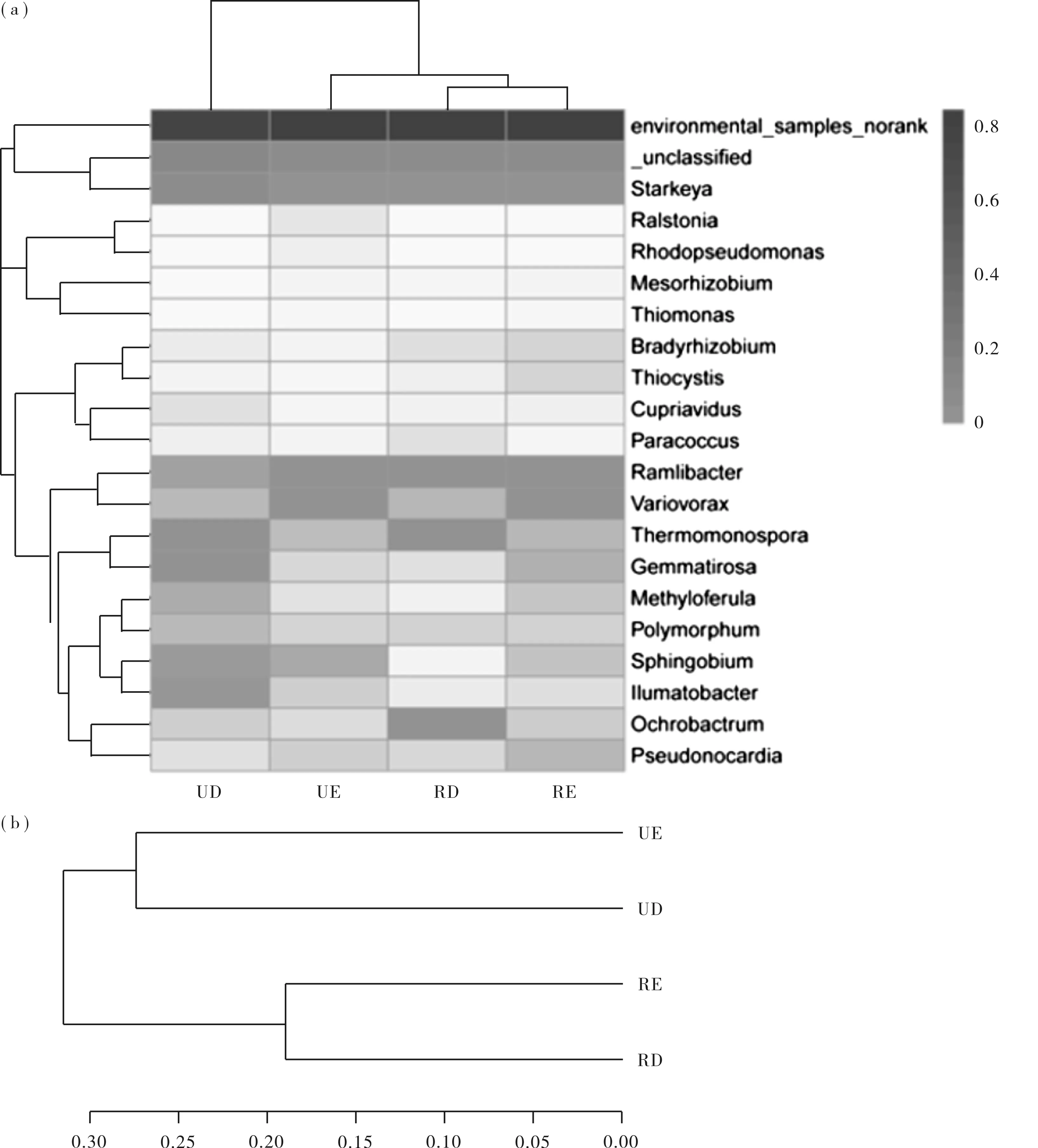
图4 四个样点土壤自养细菌群落的层次聚类分析Fig.4 Hierarchical cluster analysis of autotrophic bacterial communities in four sample sites
3.5 自养细菌群落与土壤理化性质的关系
自养细菌前50位OTU与14个土壤理化性质指标之间的相关性热图(图5)显示:土壤理化性质聚类为2类(第一类为土壤有机碳、粉粒含量、黏粒含量、TK、TN和AN;第二类为土壤pH、含砂量、TP、有机磷、AK、水分和容重)。2类菌群与自养菌的OTU的相关性正好相反;OTU 135和OTU 16是所有土壤样品中自养细菌的优势OTU。第一类土壤理化性质均与这2个优势OTU呈负相关,而第二类土壤理化性质均与这2个优势OTU呈正相关。其中,土壤含水率、沙粒含量和TP含量与前23位OTU呈正相关;土壤粉粒含量、黏粒含量和TN含量与其余OTU呈正相关。

图5 自养细菌前50位OTU与土壤理化性质的相关性热图Fig.5 The correlation heat map between the top-fifty OTUs of autotrophic bacteria and the physical and chemical properties of soil
4 结论与讨论
测序结果表明,甘肃省罗峪沟流域的主要分支—桥子沟流域的土壤固碳细菌以兼性自养菌为主,优势菌株主要为变形菌门、放线菌门和芽单胞菌门。在未开垦坡面和刺槐坡面中,侵蚀区的自养细菌多样性明显低于沉积区的。此外,PCoA分析和系统发育树分析表明,在同一坡面,侵蚀区的土壤自养细菌群落结构与沉积区的也存在一定的差异。土壤质地(沙粒、粉粒和黏粒含量)与自养细菌群落结构显示出较强的相关性。综合以上结果,黄土高原土壤侵蚀-沉积扰动对土壤自养细菌群落结构的异质性有一定影响。
甘肃省罗峪沟流域的主要分支—桥子沟流域的土壤颗粒细小,随雨水径流易迁移。因此,随着土壤侵蚀量的增加,土壤中粉粒和黏粒的含量会减少,砂粒的比例则会相应增加。同一坡面侵蚀点和沉积点的大部分养分(如TN、TK、TP和AN)含量没有显著差异。这可能是因为刺槐坡面和未开垦坡面的土壤质地以粉粒为主,因此土壤侵蚀不会对含量少的成分造成显著影响。侵蚀区植被恢复减少了养分的流失,这可以解释同一坡面上部分养分含量差异不显著的原因。除此之外,植物残渣、根系和根系分泌物也使土壤中有机质有所增加[14]。
在刺槐坡面和未开垦坡面中,均发现沉积区的自养细菌多样性高于侵蚀区的。土壤侵蚀引起的选择性运输导致轻质部分有机碳优先运输到沉积地,并且大部分丰富的不稳定有机碳和养分沉积物的积累有利于沉积地自养细菌群落的生长[15]。同时,侵蚀地的土壤微生物群落也可能直接受到水文过程的影响,雨滴和径流引起的飞溅、碰撞和剪切力可以直接诱导土壤表面微生物的死亡,特别是对于团聚体破碎后释放的微生物[16]。因此,沉积区应该更有利于微生物的累积,导致了丰度的增长。
基于RuBisCO基因的分子方法是探索这种系统发育上非常多样化的细菌群特征的有力方法[17]。RuBisCO作为CBB循环中的关键基因,广泛存在于多种分类中,包括α-变形杆菌、β-变形杆菌和γ-变形杆菌、厚壁菌和氯屈曲菌[18]。本研究根据测序结果发现的许多细菌与碳循环有关。比如革兰氏阴性细菌(尤其是变形杆菌)与土壤中的无机碳固定密切相关[19],属于厚壁菌门的克氏杆菌属(Ktedonobacter)其相对丰度随着CO2浓度梯度增加而显著增加[20]。厚壁菌门是黄土高原丘陵沟壑区土壤中的优势菌种之一,能产生降解底物的胞外酶(如纤维素酶、脂肪酶或蛋白酶)。它们的相对丰度与土壤水分呈负相关,这表明它们适应了干燥的土壤[21]。此外还有一些细菌种,如本研究中发现的慢生根瘤菌(Bradyrhizobium)、红假单胞菌(Rhodopseudomonas)和罗氏菌(Ralstonia)在现有研究中很少发现[13]。说明黄土高原侵蚀环境中自养细菌种群数量丰富,自养细菌群落结构组成和形成是与环境条件长期相适应的结果。
自养细菌群落结构的相似性和差异性分析结果表明,在同一侵蚀坡面上,侵蚀区的自养细菌群落结构与沉积区的有一定差异。其原因是,在同一坡面上,侵蚀区的土壤有机碳和可溶性碳含量与沉积区的均存在一定差异。本研究中的自养细菌分专性自养细菌和兼性自养细菌。其中,专性自养细菌在生长过程中只能将二氧化碳作为唯一碳源,而兼性自养细菌还可以使用有机物质作为其替代碳源,代谢方式灵活。因此,兼性自养细菌在营养物质丰富的环境中有优势,专性自养细菌则更能适应有机质和其他营养物质含量较低的土壤环境[17,22]。黄壤土壤颗粒组成以粉粒为主,而红壤土壤颗粒组成以粘粒为主,其土壤水分含量更为丰富,为有机物质提供了稳定的环境[23]。同时,黏粒相比于砂粒能携带更多表层土壤的轻有机物质,其反复堆积会促进大量有机物质的埋藏,导致氧气减少以至没有足够的有机物质供给,有机物质的周转速率降低,有机物质含量高[24]。因此,有机碳相较于其他土壤养分的含量,对细菌群落结构的影响更为显著。
土壤微生物群落组成能够调节或改变生态系统的功能,同时与土壤理化特性有着密切的关系[25-26]。土壤pH值和含水率可以影响不同类型土壤中微生物群落的组成,对自养细菌群落中基质(如氨和二氧化碳)的有效性有显著影响,从而强烈影响土壤C循环。土壤pH值对土壤中的CO2浓度有着显著的影响,因此,不同的CO2固定途径受到土壤pH值的限制[27]。氮被认为是微生`物生存最基本的元素之一,氮的浓度与某些特定分类群有关[28]。研究发现,土壤总氮通过影响有机碳和pH值的有效性在一定程度上影响土壤微生物群落,进而影响土壤生物量[29]。cbbL基因主要存在于粉粒和粘粒中,而在不同管理的农业土壤中,沙土中几乎没有cbbL基因[30]。以上研究结果表明,植被覆盖和土壤处理都会影响土壤细菌群落,因为土壤性质控制着生物地球化学过程和生态系统功能。
猜你喜欢
当代水产(2022年8期)2022-09-20
水土保持学报(2022年4期)2022-08-16
科海故事博览·下旬刊(2022年4期)2022-05-07
昆明医科大学学报(2022年2期)2022-03-29
农业工程学报(2022年1期)2022-03-25
山西农业大学学报(自然科学版)(2021年4期)2021-09-23
党员生活·下(2021年2期)2021-04-14
排灌机械工程学报(2021年1期)2021-01-19
农家致富顾问·下半月(2020年2期)2020-05-11
学校教育研究(2020年7期)2020-04-09
