
尖峰岭次生林和原始林林下灌木叶氨基酸对氮添加的响应
2021-08-24 09:41:04李修平安丽华倪晓凤吉成均
北京大学学报(自然科学版) 2021年4期
李修平 安丽华 倪晓凤 吉成均,†
1. 北京大学城市与环境学院生态学系, 北京大学地表过程分析模拟教育部重点实验室, 北京 100871;2. 北京大学医药卫生分析中心, 北京 100191; † 通信作者, E-mail: jicj@pku.edu.cn
氮沉降作为全球重大环境问题之一, 直接影响森林生态系统的养分可利用状况和植物的生长[1-2]。近年来, 国内外相继建成多个森林生态系统养分添加控制实验平台, 通过人工氮添加实验来模拟大气氮沉降对森林生态系统的影响[3-6]。在森林植物氮代谢的变化方面, 有研究表明氮添加能促进植物对氮的吸收, 导致叶片含氮量增加, 改变叶片氮分配格局[7-8]。我国也有一些相关报道, 如亚热带常绿阔叶林植物叶片的含氮量在氮添加条件下显著地增加[9]; 虽然热带次生林林下植物叶片总氮含量的响应在统计意义上不显著, 但其叶片氮磷比受到氮添加的显著影响[10]; Liu 等[11]通过研究中国氮沉降的影响发现, 从 20 世纪 80 年代到 21 世纪初, 木本植物和草本植物的叶片含氮量均显著增加, 所有物种的叶片含氮量平均增加 32.8%。
植物体内的氮主要以水解氨基酸(即结合氨基酸)、游离氨基酸和无机氮(铵态氮和硝态氮)等形式贮存[12]。目前, 关于氮添加情景下氮代谢的变化研究多关注总氮或铵态氮、硝态氮的变化, 集中于养分平衡、氮周转速率以及氮利用效率等方面, 而对含氮有机物, 特别是氨基酸的关注较少[1,13-14]。
作为植物吸收同化氮素的初级含氮代谢分子,氨基酸是大气氮沉降进入植物的首要合成物质[15]。在氮沉降情景下, 植物优先选择将多余的氮保存在氨基酸中, 经过酶的催化作用, 参与蛋白质合成,改变植物的生长[16]。可以将氨基酸视为植物与环境氮素之间的枢纽, 其在不同环境条件下的合成策略直接或间接地影响植物生长发育的各个方面, 并作用于氮源的同化以及源/库转运[17]。因此, 研究不同氮添加条件下氨基酸含量的变化, 可以从有机氮分配、氮利用效率和构建成本等新视角了解氮沉降情景下森林生态系统植物的响应与适应机制, 为制定相应的森林保护和改良对策提供科学依据。
目前, 针对植物氨基酸对氮添加的响应, 已有一些初步的研究结果, 发现植物组织中的游离氨基酸含量可以比总氮含量更好地响应大气氮沉降或氮添加[18-19]。但是, 这些研究多针对单个物种进行监测[20-21], 少量对森林生态系统氨基酸响应氮沉降的研究集中在针叶植物[22-26], 国内除在鼎湖山进行的苗圃实验[27]外, 有关热带山地雨林植物氨基酸含量对氮添加的响应方面尚未见报道。
热带森林贡献了 70%以上的全球森林总碳汇[28],是陆地生态系统碳循环中重要的组成部分。研究表明, 氮沉降能使热带森林叶片氮含量显著升高[29],但对叶氨基酸含量的响应关注较少。根据受人类活动干扰程度的不同, 可将热带森林分为原始林和次生林两种类型。热带次生林和原始林植物生长对氮沉降的响应存在差异, 次生林生产力对氮沉降的响应更为敏感[30-31]。因此, 热带次生林和原始林植物的氨基酸含量对氮沉降的响应是否存在差异, 这一问题值得研究。探讨热带森林系统中植物叶氨基酸含量在氮添加条件下的变化情况, 有助于对全球不同林型植物响应氮沉降的相关研究进行补充。
本研究的实验依托中国森林养分添加实验平台(Nutrient Enrichment Experiments in Chinese Forests,NEECF)的尖峰岭原始林和次生林施肥样地[6], 利用人工氮添加实验来模拟大气氮沉降对森林生态系统的影响。通过分析林下常见植物的叶氨基酸含量,探讨以下科学问题: 1) 原始林和次生林林下灌木的叶氨基酸含量对氮添加的响应规律; 2) 林下灌木叶氨基酸对氮添加的响应程度与氨基酸含量占比和氨基酸含氮量的关系。
1 研究区概况
研究区位于海南尖峰岭自然保护区的热带山地雨林, 经纬度为 18°43′N, 108°53′E, 海拔 870 m。研究区属低纬度热带岛屿季风气候, 雨热同期, 降水丰富, 干湿两季明显, 年均气温为 24.7℃[32], 年均降水量为 2449 mm, 年均蒸发量为 1249 mm[33], 本地氮沉降速率为 25 kg N/(ha·a)[6]。
研究区物种多样性强, 成分复杂, 乔木层、灌木层和草本层层次分明, 层间多分布附生蕨类和气生兰类等植物, 具有典型的热带森林特征[34-35]。常见林下植物有多刺鸡藤(Calamus tetradactyloides)、射毛悬竹(Ampelocalamus actinotrichus)、大叶白颜树(Ginonniera subaequalis)和棕竹(Rhapis excelsa)等, 多为灌木。
研究区原始林是长期未受到人类活动干扰的天然成熟林, 次生林则是在 20 世纪 60—70 年代原始林皆伐迹地上发育的天然更新林。两种林型的气候条件相似, 但其森林结构和功能、植物物种组成和土壤特性(尤其是土壤含氮量)存在差异(表 1)。
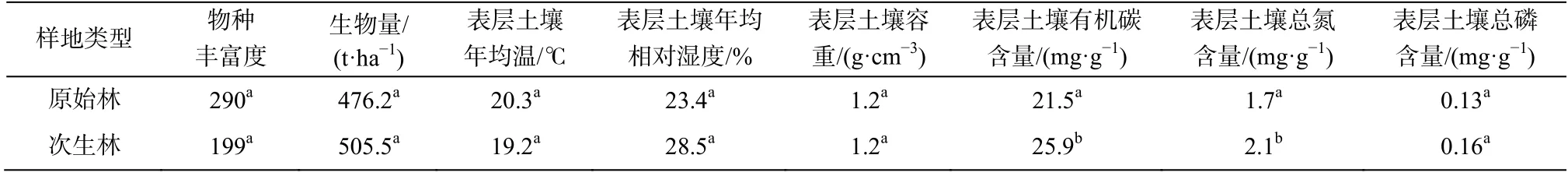
表1 原始林和次生林样地基本信息[36]Table 1 Basic information of primary and secondary forests[36]
2 研究方法
2.1 实验设计
实验样地为 NEECF 项目养分添加实验平台的实验样地之一, 建立于 2010 年 9 月, 包含原始林和次生林两种林型, 均设置对照 CK (0 kg N/(ha·a))、中氮添加 M (50 kg N/(ha·a))和高氮添加 H (100 kg N/(ha·a)) 3 个氮浓度梯度, 且按照随机试验的方式,将样地划分为 3 种氮添加处理 ×3 个重复的 9 个小样方, 每个小样方大小为 20 m × 20 m, 样方之间设置至少 10 m 的缓冲带。
使用硝酸铵(NH4NO3)进行施氮处理, 施氮方式如下: 将硝酸铵水溶液喷洒至实验样地的森林地面上, 在控制样地则喷洒相同体积不含氮的水, 施氮时间为全年, 频率为每月一次。
2.2 样品采集和处理
采样时间为 2019 年 1 月, 与下次施氮时间相隔1 周。采集叶片时, 优先选择两种林型的共有种。在每个样方, 每种植物选取 3 株进行采样, 采集植株北面或东北面的健康成熟叶片, 避免光照影响。本次采样所选择的林下灌木物种信息见表 2。
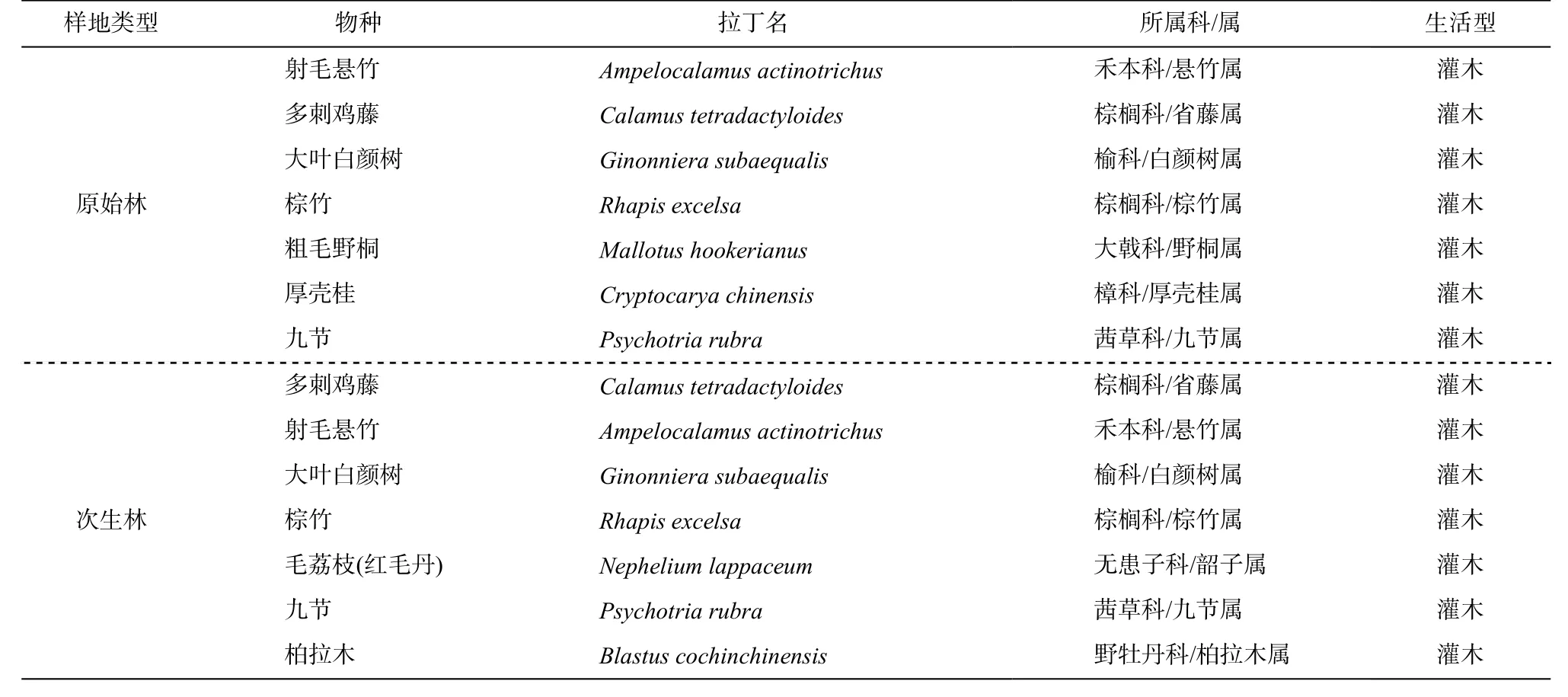
表2 尖峰岭样地林下灌木采集物种Table 2 Species collection of understory shrubs in Jianfengling site
将采集的植物叶片样品用蒸馏水反复冲洗至无泥土, 并剔除杂质, 装入信封中带回实验室, 40℃烘干至恒重。用 ZM 200 型研磨仪磨碎, 过 100 目筛,混合均匀后密封保存。
水解氨基酸样品处理: 在充氮密封条件下, 将样品用 6 mol/L 的盐酸在 110℃水解 22 小时。进行显色预实验(取 20 μL 水解液加入 200 μL 茚三酮反应液中, 混匀, 100℃水浴 3 分钟, 与氨基酸标样(含氨基酸 100 ng/mL)进行颜色对比), 确定适当的稀释倍数。取适量水解液放入离心管中, 用浓缩仪蒸干盐酸。向离心管中加入 0.02 mol/L 盐酸 1 mL, 溶解样品。以 10000 rpm 的转速离心 15 分钟, 析出沉淀物,上清液经 0.45 μm 滤膜过滤后上机测试。
游离氨基酸样品处理: 将适量样品置于离心管中, 加入适量 0.05 mol/L 盐酸, 室温下超声波振荡萃取 30 分钟。加入浓度为 5%~10%的磺基水杨酸,充分混匀, 静置 1 小时, 去除溶液中的蛋白质。进行显色预实验(步骤同水解样品), 确定适当的稀释倍数。取适量游离液入离心管中, 加入 0.02 mol/L盐酸稀释至 1 mL, 以 10000 rpm 的转速离心 15 分钟,上清液经 0.45 μm 滤膜过滤后上机测试。
使用全自动氨基酸分析仪(日立 L8900)进行氨基酸溶液测定, 通过阳离子交换柱和茚三酮反应柱进行氨基酸离子交换和显色反应。氨基酸分析仪内置可见光分光光度计, 进行两种波长(570 nm (用于测定除脯氨酸外的其余氨基酸)和 440 nm (用于测定脯氨酸))下的测定, 得到样品溶液的色谱图, 其中曲线与横坐标(基准线)所围成的面积即为溶液中的氨基酸含量(ng/20μL), 经过换算得到对应叶片样品的氨基酸含量(mg/g)。测定的氨基酸包括天冬氨酸、苏氨酸、丝氨酸、谷氨酸、脯氨酸、甘氨酸、丙氨酸、半胱氨酸、缬氨酸、甲硫氨酸(蛋氨酸)、亮氨酸、异亮氨酸、苯丙氨酸、酪氨酸、赖氨酸、组氨酸和精氨酸等 17 种。
2.3 参数选用与数据统计分析
响应程度指氮添加样地叶氨基酸含量与对照样地叶氨基酸含量的相对变化量(relative variation),用来表征叶氨基酸的响应情况, 计算公式为
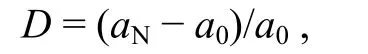
其中,D表示响应程度,aN表示氮添加样地叶氨基酸含量,a0表示对照样地叶氨基酸含量。对照样地的D值为 0, 将 0 作为对照值, 若计算得出的响应程度与对照样地的 0 值有显著差异, 表明该氮添加条件下叶氨基酸存在显著响应, 反之则无显著响应。
用氨基酸含量占比表示该氨基酸含量占总氨基酸含量的比例。因水解氨基酸与游离氨基酸的含量(mg/g)的数量级差异过大, 为使横坐标轴相对一致,且考虑到响应程度(纵坐标)的数值多在 0~1 之间,故使用含量占比替代真实含量来建立关系。用氨基酸含氮量表示该氨基酸分子中氮元素占总分子量的比例。使用 Excel 2016 进行数据处理和绘图, 使用SPSS 18.0 进行独立样本 t 检验和单因素方差分析(one-way ANOVA)。
3 实验结果
3.1 次生林和原始林林下灌木叶氨基酸的本底值
比较原始林与次生林对照样地林下灌木的叶氨基酸含量(表 3)可知, 次生林的水解氨基酸总含量(94.6478 mg/g)略高于原始林(86.6151 mg/g), 但差异不显著; 游离氨基酸则表现为原始林总含量(1.9442 mg/g)显著高于次生林(1.0513 mg/g) (p<0.05)。由于水解氨基酸总含量远高于游离氨基酸, 故氨基酸总含量也呈现次生林(95.6991 mg/g)略高于原始林(88.5593 mg/g)的结果, 但差异不显著。
从表 3 看出, 次生林中 17 种水解氨基酸的含量均略大于原始林, 但差异不显著; 次生林中 17 种游离氨基酸含量多数显著低于原始林(p<0.1)。
3.2 次生林和原始林林下灌木叶氨基酸对中氮添加的响应
比较游离氨基酸总量与水解氨基酸总量(图 1)可知, 在中氮添加条件下, 次生林水解氨基酸的响应程度显著大于原始林(次生林D=0.0998, 原始林D=0.0696,p<0.05), 游离氨基酸的响应情况与之相似(次生林D=0.4978, 原始林D=0.2228,p<0.05)。
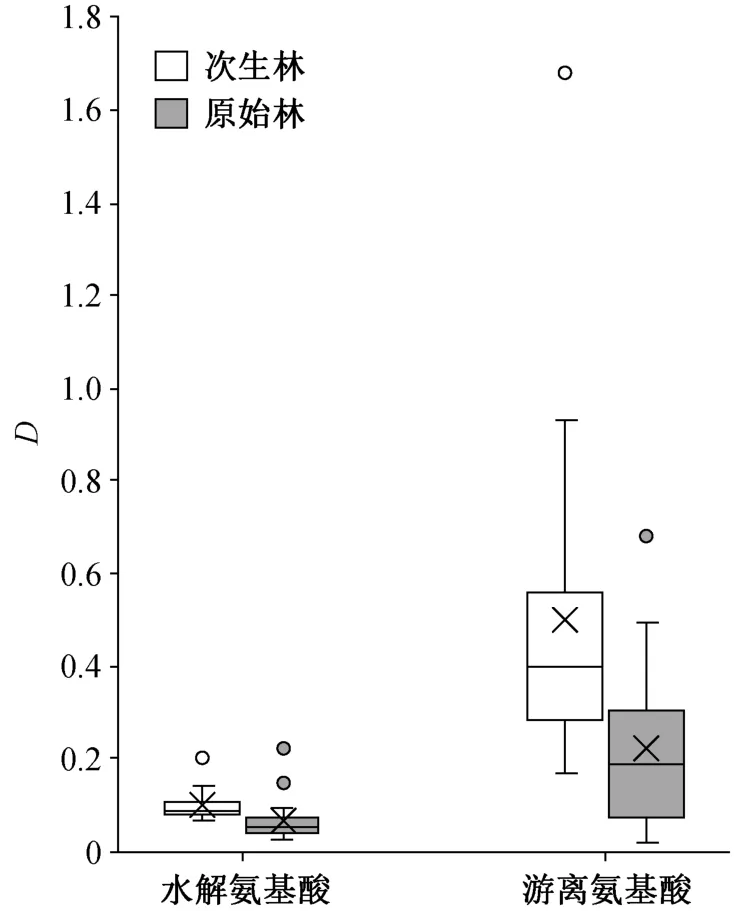
图1 次生林和原始林中林下灌木叶氨基酸对中氮添加的响应程度Fig. 1 Relative variations of amino acid contents for middle nitrogen addition in leaves of understory shrub species in primary forest and secondary forest
对 17 种氨基酸而言, 在中氮添加条件下, 次生林(除甲硫氨酸外)和原始林水解氨基酸的响应程度与对照样地的 0 值相比均无显著差异, 即对氮添加的响应不显著; 次生林(除半胱氨酸和酪氨酸外)和原始林(除丙氨酸外)游离氨基酸的响应程度与对照样地的 0 值相比亦均无显著差异, 即对氮添加的响应不显著。从图 2 可以看出, 植物叶氨基酸含量对氮添加响应的方向存在差异, 使得整体上对中氮添加的响应程度不高。
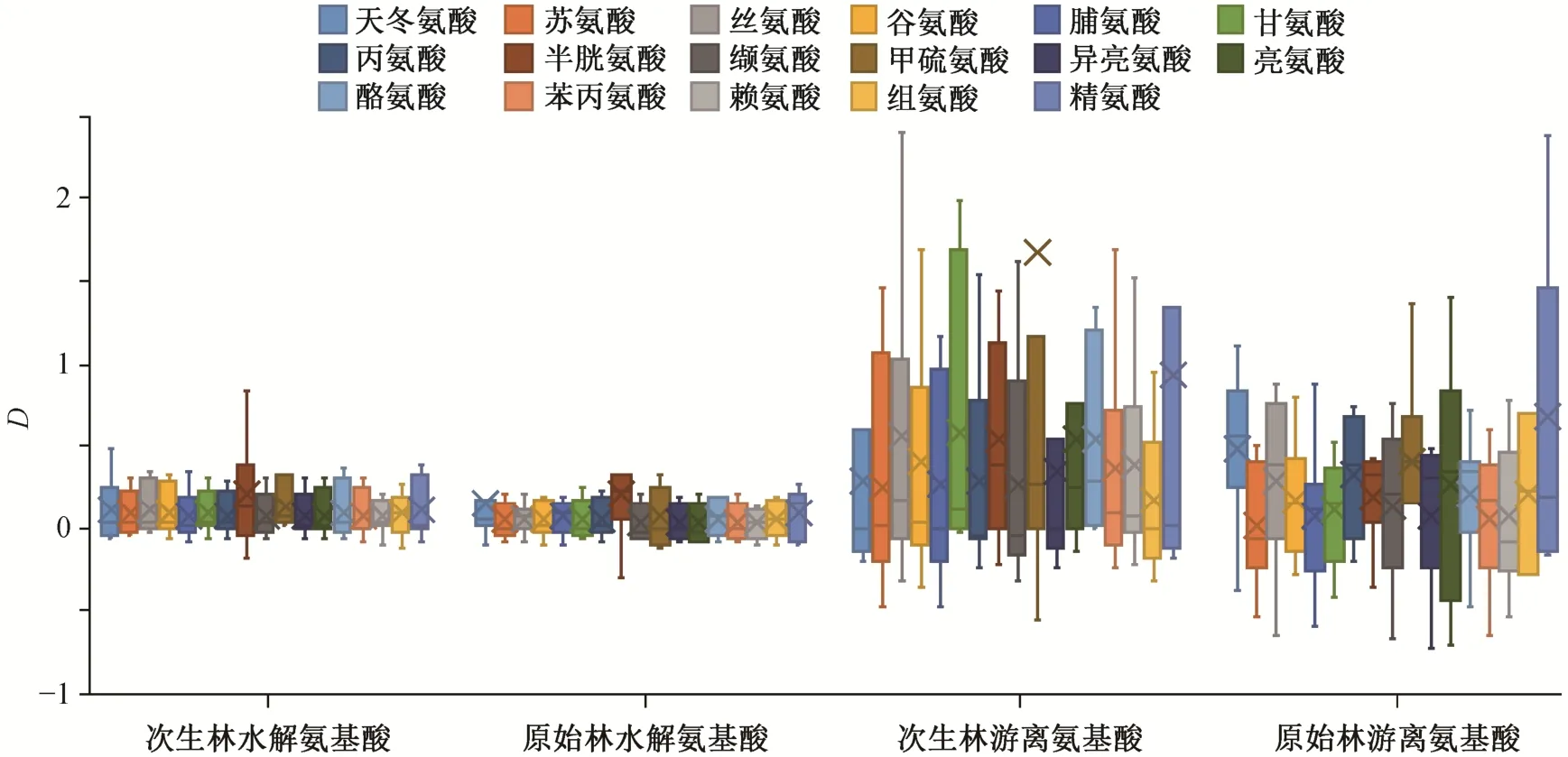
图2 17 种氨基酸对中氮添加的响应程度Fig. 2 Relative variations of 17 amino acid contents for middle nitrogen addition
分析氨基酸响应程度与氨基酸含量占比的关系(图 3)可知, 次生林水解氨基酸对氮添加的响应程度随氨基酸含量占比的增加而显著减小(p=0.033),原始林也有相同的趋势, 但不显著; 次生林和原始林游离氨基酸对氮添加的响应程度均不显著, 次生林有随氨基酸含量占比增加而减小的趋势, 原始林则有微弱的相反趋势。
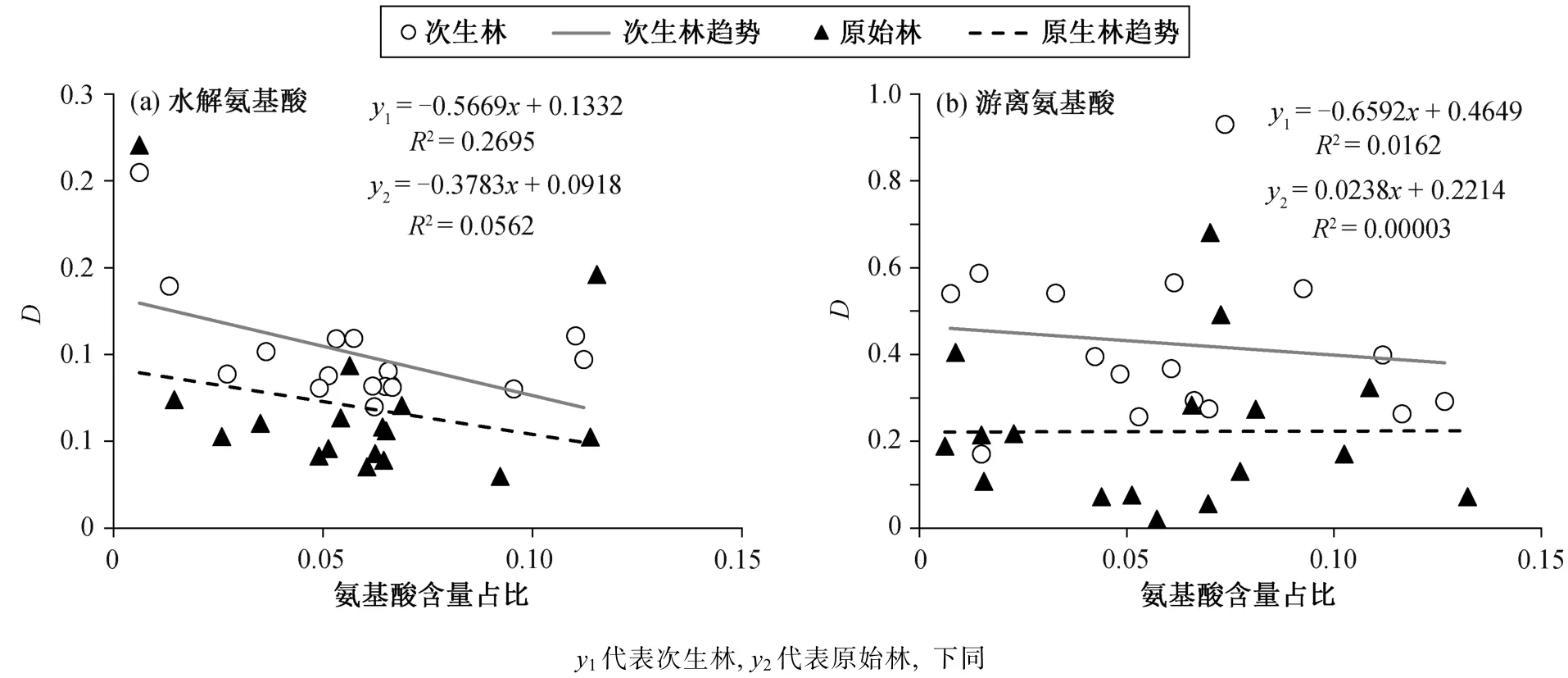
图3 氨基酸对中氮添加的响应程度与氨基酸含量占比的关系Fig. 3 Relationship between relative variations and proportions of amino acid contents for middle nitrogen addition
从氨基酸对氮添加的响应程度与氨基酸含氮量的关系(图 4)可知, 次生林与原始林氨基酸含量占比的变化趋势基本上一致。水解氨基酸的响应程度有随氨基酸含氮量增加而减少的趋势, 游离氨基酸的响应程度有随氨基酸含氮量增加而增加的趋势。
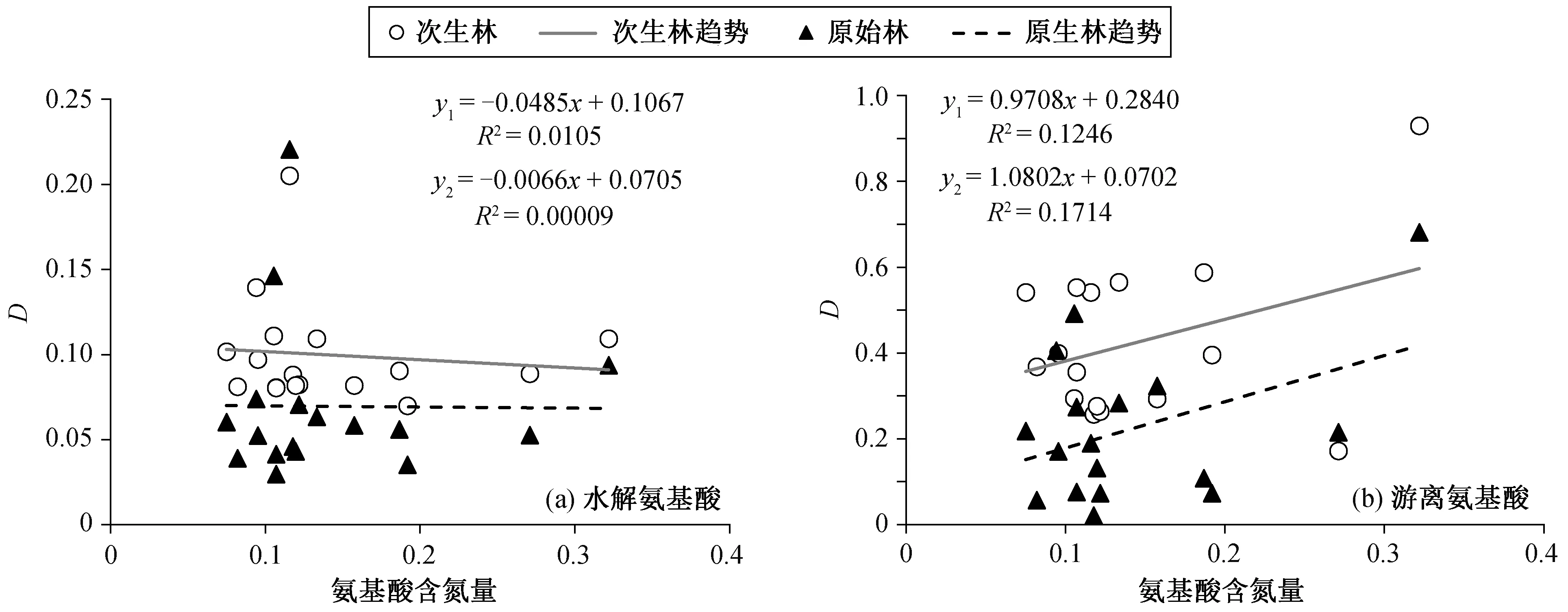
图4 氨基酸对中氮添加的响应程度与氨基酸含氮量的关系Fig. 4 Relationship between relative variations and nitrogen contents of amino acids for middle nitrogen addition
3.3 次生林和原始林林下灌木叶氨基酸对高氮添加的响应
从图 5 可以看出, 在高氮添加条件下, 水解氨基酸的响应程度在次生林与原始林之间无显著差异(次生林D=0.0951, 原始林D=0.1009), 游离氨基酸的响应程度则次生林显著低于原始林(次生林D=0.3995, 原始林D=0.6806)(p<0.05)。
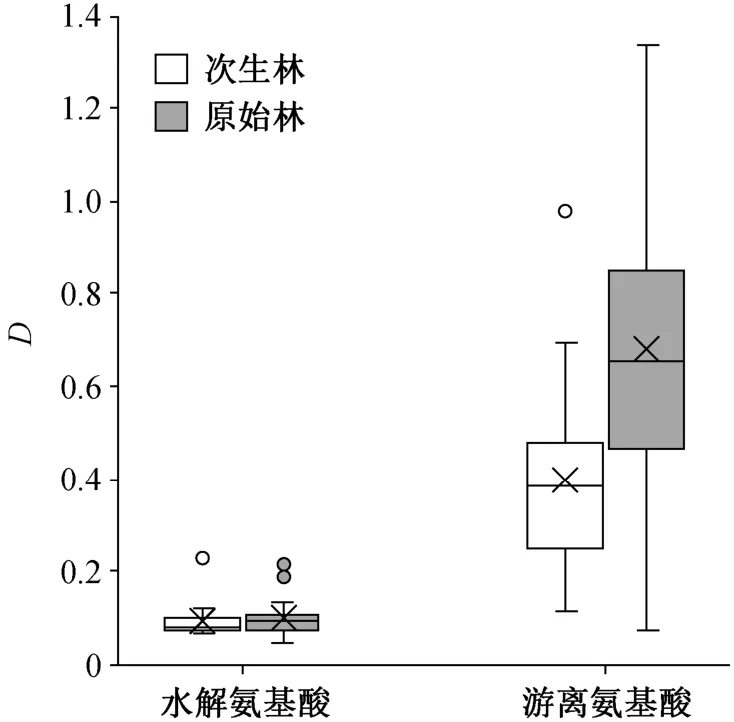
图5 不同林型的林下灌木叶氨基酸对高氮添加的响应程度Fig. 5 Relative variations of amino acid contents for high nitrogen addition in leaves of understory shrub species in primary forest and secondary forest
图 6 显示, 对 17 种氨基酸而言, 在高氮添加条件下, 次生林(除半胱氨酸外)和原始林(除甘氨酸、甲硫氨酸、亮氨酸、酪氨酸和组氨酸外)中水解氨基酸的响应程度与对照样地的 0 值有显著差异(p<0.1), 次生林(除天冬氨酸、苏氨酸、谷氨酸、脯氨酸、缬氨酸、甲硫氨酸、异亮氨酸和精氨酸外)和原始林(除谷氨酸、脯氨酸、甘氨酸、酪氨酸、苯丙氨酸、赖氨酸和精氨酸外)中游离氨基酸的响应程度与对照样地的 0 值有显著差异(p<0.1)。上述结果表明, 与中氮添加相比, 更多种类的叶氨基酸对高氮添加表现为正响应。
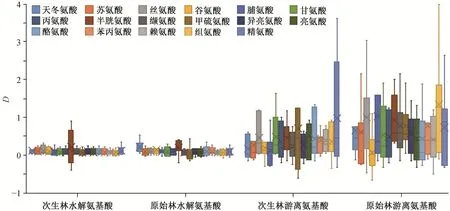
图6 17 种氨基酸对高氮添加的响应程度Fig. 6 Relative variations of 17 amino acid contents for high nitrogen addition
从氨基酸对氮添加响应程度与氨基酸含量占比之间的关系(图 7)可以发现, 对水解氨基酸而言, 次生林的响应程度呈现随其含量占比增加而减小的趋势, 原始林则相反; 对游离氨基酸而言, 次生林和原始林的响应程度均呈现随其含量占比增加而减小的趋势。
通过分析响应程度与氨基酸含氮量的关系(图8)可知, 次生林与原始林的变化趋势较为一致。水解氨基酸对氮添加的响应程度有随氨基酸含氮量增加而减少的趋势, 但不显著; 游离氨基酸对氮添加的响应程度有随氨基酸含氮量增加而增加的趋势,并在次生林中表现显著(p=0.036)。

图8 氨基酸对高氮添加的响应程度与氨基酸含氮量的关系Fig. 8 Relationship between relative variations and nitrogen contents of amino acids for high nitrogen addition
3.4 不同氮添加条件下次生林和原始林林下灌木叶氨基酸响应的差异
通过对比氨基酸对中氮添加和高氮添加的响应程度(图 9)可知, 次生林中两种条件下氨基酸响应程度无显著差异(水解氨基酸: 中氮添加时D=0.0998,高氮添加时D=0.0951; 游离氨基酸: 中氮添加时D=0.4978, 高氮添加时D=0.3995); 原始林中氨基酸对高氮添加的响应程度显著高于中氮添加(水解氨基酸: 中氮添加时D=0.0696, 高氮添加时D=0.1009; 游离氨基酸: 中氮添加时D=0.2228, 高氮添加时D=0.6806) (p<0.05)。
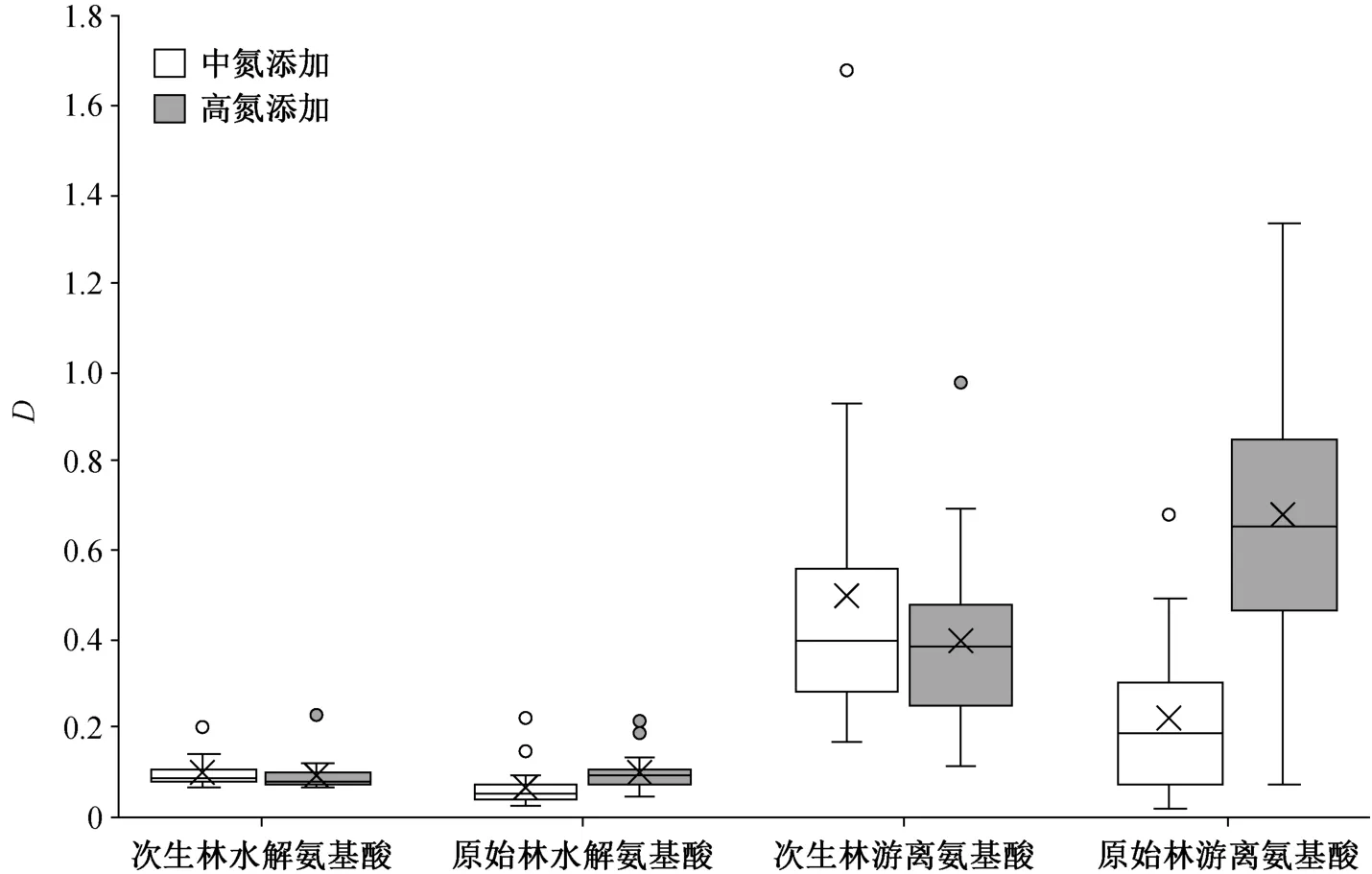
图9 不同林型林下灌木叶氨基酸对氮添加的响应程度Fig. 9 Relative variations of amino acid contents for nitrogen addition in leaves of understory shrub species
也就是说, 原始林中氨基酸对高氮添加的响应程度更大, 而次生林中氨基酸对两种氮添加的响应程度基本上一致。在响应程度与氨基酸含量占比的关系方面, 原始林与次生林的趋势相对较为一致,均有响应程度随氨基酸含量占比增加而减小的趋势。在响应程度与氨基酸含氮量的关系方面, 次生林与原始林的趋势相对一致, 但水解氨基酸与游离氨基酸的趋势相反, 即随着氨基酸含氮量的增加,水解氨基酸响应程度有减小的趋势, 游离氨基酸响应程度有增加的趋势。
4 讨论
4.1 次生林和原始林林下灌木叶氨基酸含量对氮添加响应的差异
中氮添加时, 次生林林下灌木叶氨基酸含量对中氮添加的响应程度高于原始林, 原因可能是次生林与原始林林下灌木对土壤氮的需求存在差异以及这两种林型的叶氨基酸含量存在差异。通常认为,氮添加对大多数热带原始森林的生长没有促进作用, 而对某些热带次生森林的生长有促进作用[30,37-41],如马素辉[36]通过实验发现, 尖峰岭次生林中部分物种在中氮添加时就表现出叶片氮元素的积累现象,即次生林的生长受到氮相对不足的限制。本研究对无氮添加样地叶氨基酸含量的研究表明, 原始林游离氨基酸总含量显著地高于次生林, 或许暗示尖峰岭的次生林更缺氮, 使得该地次生林植物对中氮添加比原始林更敏感, 因此在中氮添加条件下, 次生林林下灌木叶氨基酸含量的响应程度高于原始林。
高氮添加时, 原始林水解氨基酸的响应程度与次生林无显著差异, 而游离氨基酸的响应程度高于次生林。由于尖峰岭的本底氮沉降值(25 kg/(ha·a))较高[6], 长期的高氮添加可能导致土壤氮过量, 我们推测这种游离氨基酸的响应差异可能与氮过量有关。大量的研究表明, 一定程度的氮添加可以促进植物生长, 过量的氮添加则会抑制植物生长[14,16,37,42-43]。氮过量时, 植物可以通过将多余的氮储存在氨基酸中来避免含氮物质(如氨)过高带来的生理伤害[16]。除作为合成蛋白质的原料库外, 游离氨基酸在防御病虫害和抵抗逆境方面也具有重要的功能[15]。由于尖峰岭原始林林下灌木游离氨基酸的总含量显著高于次生林, 原始林林下灌木的氮过量现象可能更明显, 因此表现为原始林游离氨基酸的响应程度高于次生林。
本研究还发现, 次生林和原始林各氨基酸的响应程度与该氨基酸含量的关系基本上一致, 原因可能是本次实验选择的次生林和原始林物种基本上相同, 所以这两种林型的氨基酸对氮添加响应在一定程度上具有相似性。
4.2 植物叶氨基酸对氮添加响应随氨基酸含氮量及氨基酸含量占比的变化规律
研究表明, 部分含量较高或含氮量较高的氨基酸对氮添加较为敏感[44-47]。本文发现, 水解氨基酸对氮添加的响应程度随含氮量的变化规律不明显,游离氨基酸对氮添加的响应程度则表现出随含氮量增加而增大的规律。在氨基酸含量占比方面, 游离氨基酸对中氮添加的响应程度随氨基酸含量占比的变化规律不明显, 对高氮添加的响应程度则表现出随氨基酸含量占比增加而减小的趋势。有关氨基酸对氮添加的响应程度随氨基酸含氮量的变化规律,本文结论与文献[44-47]基本上一致, 说明植物倾向于选择含氮量高的氨基酸, 以便提高贮存效益。有关氨基酸对氮添加的响应程度随氨基酸含量占比的变化规律, 本文结果与文献[44-47]存在差异, 可能是由于文献[44-47]中探讨氨基酸含量的影响时主要关注某些具体的氨基酸, 如精氨酸和赖氨酸等,而未讨论氨基酸响应程度与氨基酸含量的关系。本文通过综合研究 17 种氨基酸, 发现氨基酸含量占比与含氮量之间并非显著正相关, 如精氨酸含氮量最高, 赖氨酸含氮量也相对较高, 但氨基酸含量占比最高的为谷氨酸、脯氨酸和丙氨酸。因此, 相比于较高的本底含量, 植物可能更倾向于选择含氮量较高的氨基酸来贮存氮。
本研究的施氮样地没有出现北方针叶林中常见的精氨酸含量剧增现象。精氨酸具有较低的 C/N 值(1.5), 是植物移除多余氮素的最有效的氨基酸[19],由于北方森林长期处于氮限制状态[48], 因此当有外源氮添加时, 植物会大量吸收土壤中的氮, 而精氨酸的剧增可以保证植物吸收的氮得以有效地储存。但是, 热带森林多处于磷限制状态, 土壤氮相对充足, 故植物对氨基酸合成的分配与北方森林有一定的差异, 这也说明热带森林植物对氮的适应采取了不同于北方森林的策略。
5 结论
本文研究结果表明, 未施氮时, 尖峰岭两种林型林下灌木的总氨基酸含量和水解氨基酸含量无显著差异, 原始林游离氨基酸总含量显著高于次生林;中氮添加时, 次生林林下灌木叶氨基酸含量的响应程度高于原始林; 高氮添加时, 两种林型水解氨基酸的响应程度无显著差异, 游离氨基酸的响应程度则原始林显著高于次生林; 在氮添加条件下, 相比于本底含量占比较高的氨基酸, 植物更倾向于选择含氮量较高的氨基酸来进行氮的贮存。
致谢 感谢北京大学城市与环境学院博士研究生马素辉同学提供尖峰岭试验站背景数据, 并在样品采集过程中提供帮助。
猜你喜欢
乡村科技(2018年8期)2018-02-13 12:16:18
现代农业研究(2017年11期)2018-01-12 21:03:03
辽宁林业科技(2017年4期)2017-06-22 10:23:36
辽宁林业科技(2017年4期)2017-06-22 10:23:36
诗潮(2017年2期)2017-03-16 10:42:30
湖南林业科技(2017年1期)2017-02-06 05:29:17
现代园艺(2017年20期)2017-02-03 18:49:29
现代食品(2016年24期)2016-04-28 08:12:06
中国资源综合利用(2016年12期)2016-01-22 02:02:26
合成化学(2015年9期)2016-01-17 08:57:21
