
Taz及Runx2在大鼠下颌第一磨牙发育中的表达研究
2021-08-11 03:22张冰,徐欣
口腔医学 2021年7期
张 冰,徐 欣
PDZ结合基序的转录辅激活物 (transcriptional coactivator with PDZ-binding motif, Taz)是Hippo信号通路下游重要的效应因子,作为一个包含WW结构域的转录共激活因子,其在哺乳动物的多种组织器官发育中发挥着重要的调控作用[1-3]。Taz可以通过上调Runt相关转录因子2 (Runt-related transcription factor 2,Runx2),激活Smad信号通路及骨钙蛋白的表达,此外,Taz与14-3-3蛋白相互作用可以调控干细胞的分化[4]。上述研究提示Taz可能在骨生成或者某些硬组织形成过程中起重要作用。Runx2是成骨细胞分化过程中起重要作用的转录因子[5-6],参与调控骨和牙齿的发育及牙齿的矿化[7-8]。有研究发现Runx2与Taz在促进成骨细胞分化的过程中存在相互作用关系[9],但在牙齿发育中二者是否具有相关性尚不明确。本研究采用免疫组织化学的方法观察Taz、Runx2在大鼠下颌第一磨牙发育过程中的表达,以明确其时空特异性表达规律,为深入探究二者在牙齿发育中的作用机制奠定基础。
1 材料与方法
1.1 材料
Wistar大鼠(10周龄)(山东大学实验动物中心),Taz多克隆抗体(Santa Cruz公司,美国),Runx2多克隆抗体(Abcam公司,美国),免疫组织化学SABC二抗试剂盒(北京中杉金桥生物技术有限公司),DAB显色试剂盒(北京中杉金桥生物技术有限公司),其余所需试剂及设备均由山东省口腔组织再生重点实验室提供。
1.2 方法
1.2.1 获取标本 10周龄Wistar大鼠(由山东大学动物实验中心提供,实验设计经山东大学口腔医院动物伦理委员会批准通过,伦理审批号:GD202036)雌雄2∶1交配,次日检栓,发现阴栓视为受孕,当天中午即为受孕0.5 d。分别取胎龄14.5(E14.5)、E16.5、E18.5天大鼠胚胎头颅标本(各4例)及出生后0(PN0)、PN3及PN7、PN14、PN21、PN28天大鼠下颌骨标本(各4例),4 ℃条件下常规组织固定24 h(4%多聚甲醛,pH=7.2)、脱钙(10% EDTA,pH=7.2)、脱水(梯度乙醇)及石蜡包埋。大鼠胚胎头颅标本行冠状面5 μm石蜡连续切片,大鼠下颌骨标本行矢状面5 μm石蜡连续切片。经苏木精-伊红(HE)染色及免疫组织化学染色分别从形态学上观察磨牙的发育状况,从蛋白水平上检测Taz、Runx2的表达。
1.2.2 免疫组织化学染色 石蜡切片二甲苯脱蜡3次(10 min/次),梯度乙醇下行水化处理,磷酸盐缓冲液(PBS,0.01 mol/L,pH=7.2)漂洗3次(5 min/次),0.1%胰酶37 ℃孵育10 min进行抗原修复,PBS漂洗3次,3%过氧化氢37 ℃孵育20 min,以阻断内源性的过氧化氢酶。正常山羊血清封闭30 min后去除多余血清勿漂洗,滴加Taz抗体(1∶300)或Runx2抗体(1∶400)湿盒内4 ℃条件下过夜。次日,切片于室温下复温30 min,PBS漂洗3次,滴加生物素标记二抗,37 ℃孵育25 min,PBS漂洗3次,滴加酶结合物,37 ℃孵育25 min,PBS漂洗3次,由显微镜下控制DAB显色。免疫组织化学苏木素复染,梯度乙醇上行脱水、二甲苯透明,中性树胶封片。阴性对照采用PBS代替Taz、Runx2抗体4 ℃过夜,其余条件相同。
2 结 果
2.1 HE染色
牙齿的发育始于口腔上皮的增厚,E14.5天时牙板向间充质细胞中内陷形成花蕾状(图1)。随着细胞进一步增殖、分化,E16.5天时成釉器分化为3层细胞即内釉上皮层、外釉上皮层和星网状层,此期成釉器为帽状期(图1)。釉结作为一个临时性的结构呈密集的细胞团状,参与多种信号的表达调控,在牙齿形态的发生中起关键作用。在PN7天可见双层结构的上皮根鞘,上皮根鞘开始向根方增殖、延伸启动了牙根发育的形态学信号(图2)。PN14至PN28天,由牙囊的间充质细胞分化而形成的牙骨质、牙周膜、牙槽骨等结构完成了牙周组织的发育,根尖孔逐渐缩窄,牙根的发育逐步完成(图2)。
2.2 Taz、Runx2在胚胎期大鼠下颌第一磨牙牙胚中的表达与定位
在E14.5天即蕾状期, Taz在口腔上皮及其周围的间充质细胞中均有阳性表达,Runx2在蕾状期表达广泛(图1)。在E16.5天,随着牙胚的发育成釉器形状似帽子,在釉结、牙乳头及牙囊部分细胞中都存在Taz、Runx2的阳性表达,此外,Taz在口腔上皮中仍呈阳性着色(图1)。至E18.5天的钟状早期,Taz的阳性表达主要集中于内釉上皮层、外釉上皮层、中间层及牙囊处,在牙乳头及星网状层中的表达较弱,而Runx2的表达在钟状早期则主要集中于内釉上皮层、中间层、牙乳头及牙囊中(图1)。
2.3 Taz、Runx2在出生后大鼠下颌第一磨牙发育中的表达与定位
当牙胚发育至PN7天时,牙冠的发育即将完成,在成釉细胞、成牙本质细胞层、牙本质基质、中间层及上皮根鞘中存在Taz及Runx2的阳性表达, 此期成牙本质细胞在牙尖部位为高柱状,向根尖方向逐渐呈现扁平状(图2)。至PN14天时,牙冠的发育已经完成,在成釉细胞层中Taz的表达逐渐减弱;在此期的成牙本质细胞、牙本质小管及上皮根鞘处仍可见到Taz与Runx2两种蛋白的阳性表达(图2)。在牙齿发育至PN21及PN28天时,在近牙冠处的成牙本质细胞中Taz的表达较强,向根尖方向表达逐渐减弱,在根部成牙本质细胞及牙本质基质中Taz的表达呈弱阳性(图2)。Runx2在成牙本质细胞及牙本质小管中表达呈阳性。牙周膜即牙周韧带,是牙周组织中起支持作用的重要结构,为连接牙骨质和牙槽骨的结缔组织,在PN21及PN28天的免疫组织化学染色中发现Taz、Runx2在牙周膜及牙槽骨中也存在阳性着色(图2)。

eo: 成釉器; me: 间充质细胞; ek: 釉结; oee: 外釉上皮层; dp: 牙乳头; iee: 内釉上皮层; df: 牙囊; oe: 外釉上皮
3 讨 论
牙齿的发育是一个连续、复杂的上皮与间充质相互作用的过程,许多信号分子参与调控细胞增殖、分化及牙齿形态的发生[10-12],转录共激活因子Taz是一个14-3-3结合蛋白[2,13],为Yap的旁系同源蛋白,二者在蛋白质结构上有45%的相似性[2]。研究发现Yap/Taz是Hippo信号通路下游重要的效应因子,参与多种组织器官发育,二者在间充质干细胞的分化中也发挥重要作用[3]。有研究报道在小鼠正常切牙发育过程中Yap参与调控干细胞的增殖及分化[14],此外,Yap过表达可以影响细胞极性及牙齿形态的发生[15]。作为Yap的同源蛋白,Taz在牙齿发育过程的调控作用及机制目前报道尚不多见。Runx2是成骨细胞标记基因(例如骨桥蛋白、骨涎蛋白、骨钙蛋白等)的一个关键调控因子,为Runt家族的一员,具有高度保守的DNA结构域。研究发现Runx2不仅参与成骨细胞分化和骨形成的过程,同时在牙胚形成、牙齿矿化及牙周组织的发育中起重要的调控作用。近年的研究证实,Runx2的转录共激活功能是通过Taz实现的,在成骨细胞分化过程中Taz作为一个转录共激活剂在细胞核中的定位可激活Runx2的表达,从而促进骨的形成[16]。本研究通过免疫组织化学的方法观察Taz与Runx2在牙齿发育中的表达与定位,探讨二者在牙齿发育中的作用。
研究结果显示,在蕾状期,Taz、Runx2两种蛋白在口腔上皮及周围的间充质细胞中呈阳性表达;当牙胚发育至帽状期,Taz、Runx2的阳性着色存在于牙囊、牙乳头及釉结,此外,Taz在口腔上皮中仍存在表达。随着牙胚的发育Taz的表达逐渐增强,至钟状早期,整个成釉器的各层细胞及牙囊中均呈阳性表达,Runx2的表达多集中于内釉上皮层、中间层、牙乳头及牙囊。有研究报道Runx2可能调控多个信号通路的多个基因,在由蕾状期向帽状期过渡的牙胚早期发育中发挥重要作用[17],基于Taz与Runx2在牙胚早期相似的特异性表达与定位,我们推测Taz可能在牙胚早期发育过程中也发挥着重要的作用。此外,釉结呈密集的细胞团状,是调控牙齿形态重要的临时性结构[18-20]。Li 等[21]的研究报道釉结是蕾状期形成的重要结构,釉结的主要功能是分泌一系列的信号分子,共同调节牙齿形态的发生。釉结的形成主要依赖于αE-catenin通过Yap及其同源物Taz的信号传递功能。αE-catenin在牙齿发育过程中的粘附功能与其在釉结形成中的作用是分离的,釉结形成涉及到Yap/Taz信号通路的调控。本研究中免疫组织化学染色显示Taz在釉结处存在阳性着色,提示Taz可能参与牙齿形态的发生过程中信号通路的调控,与Li等的研究相近,但本研究尚未进行深入机制的研究及探讨。大鼠下颌第一磨牙牙胚的免疫组织化学染色发现,在PN7天牙冠的发育即将完成,在成釉细胞、成牙本质细胞层及牙本质基质中Taz的阳性着色最为明显,除此之外,在中间层及上皮根鞘处也检测出Taz的表达,表明Taz可能参与成釉细胞和成牙本质细胞的分化及釉质和牙本质的形成,同时在牙根的发育中也起到一定的调控作用。随着牙冠发育的完成,Taz在成釉细胞中的表达减弱,在成牙本质细胞中的阳性着色由牙冠向根尖方向逐渐降低。至PN21、PN28天,在牙周膜及牙槽骨等牙周组织中可发现Taz与Runx2均存在特异性表达。根据以往的研究发现,Taz与Runx2的相互作用可以促进成骨因子的表达,因此,我们推测Taz可能通过对Runx2的调控实现对牙槽骨、牙周膜等牙周组织发育的调节。
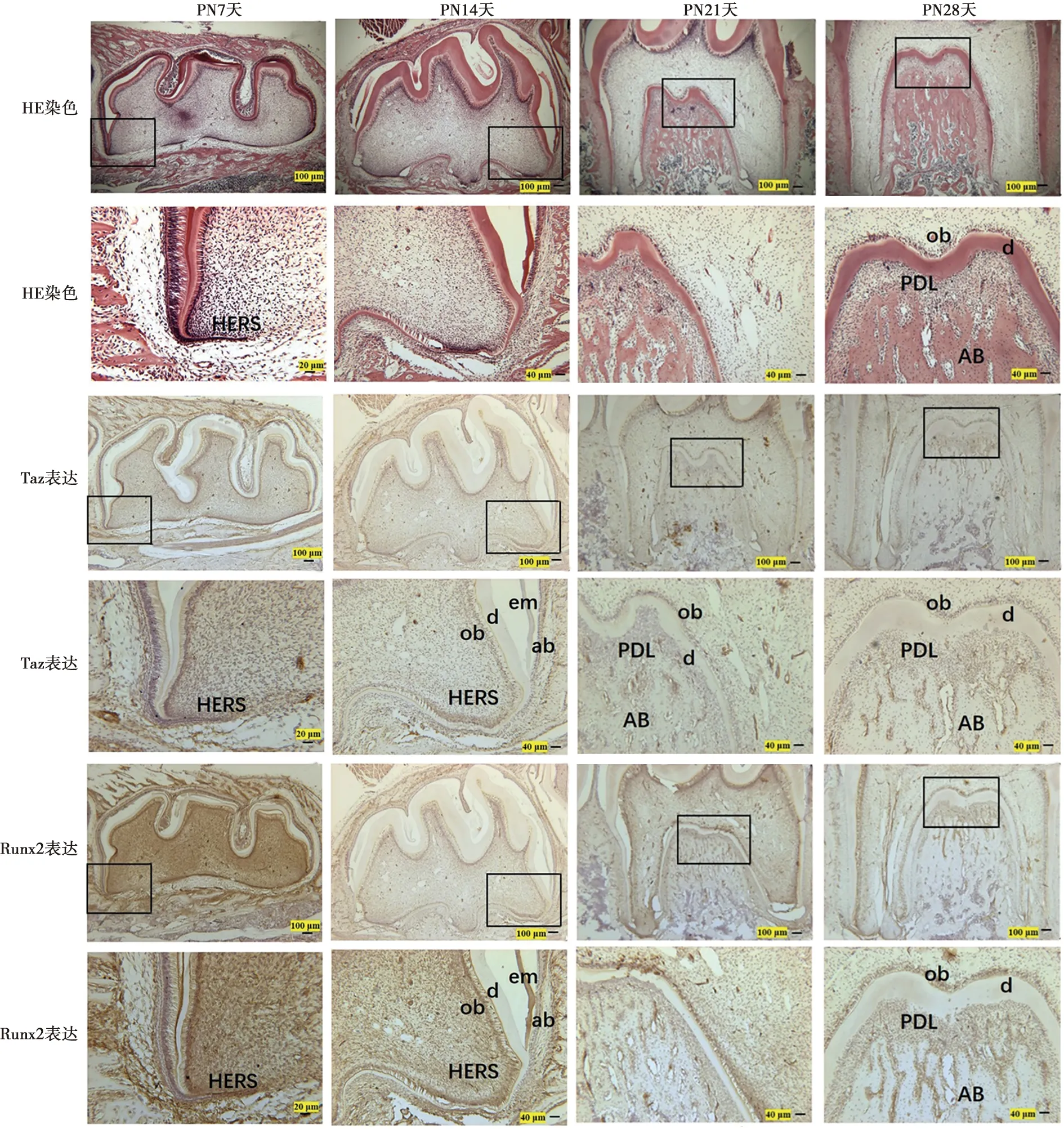
ab:成釉细胞;ob:成牙本质细胞;em:釉质基质;d:牙本质;PDL:牙周膜;AB:牙槽骨;HERS:上皮根鞘
本实验通过免疫组织化学染色观察Taz、Runx2在大鼠下颌第一磨牙发育中的时空特异性表达,结果发现Taz与Runx2的表达与定位具有一定的一致性,推测Taz可能参与牙齿发育的调控,此外,Taz可能通过调控Runx2的表达参与成釉细胞和成牙本质细胞的分化及牙根和牙周组织的发育。然而,对于具体的调控机制尚不明确仍需进一步的探索研究。
猜你喜欢
基层中医药(2022年5期)2022-10-24
大众科学(2022年7期)2022-08-19
中国典型病例大全(2022年11期)2022-05-13
江苏农业科学(2021年22期)2021-12-08
影像诊断与介入放射学(2021年5期)2021-11-29
西南医科大学学报(2021年2期)2021-06-13
英美文学研究论丛(2018年2期)2018-08-27
安徽农业科学(2015年7期)2015-07-13
华西口腔医学杂志(2013年5期)2013-11-11
华西口腔医学杂志(2013年1期)2013-05-10
