
基于BP神经网络的西北太平洋柔鱼资源丰度时空变化研究
2021-08-10 14:58王雅萌汪金涛陈新军雷林
海洋学报 2021年6期
王雅萌,汪金涛,2,3,4,5*,陈新军,2,3,4,5,雷林,2,3,4,5
(1.上海海洋大学 海洋科学学院,上海 201306;2.农业部大洋渔业开发重点实验室,上海 201306;3.上海海洋大学 国家远洋渔业工程技术研究中心,上海 201306;4.上海海洋大学 大洋渔业资源可持续开发教育部重点实验室,上海 201306;5.农业部大洋渔业资源环境科学观测实验站,上海 201306)
1 引言
柔鱼(Ommastrephes bartramii)是一种生态机会主义的大洋性种类,具有短生命周期,分为秋生群和冬春生群,广泛分布在北太平洋海域,属于高度洄游种类(图1),其资源状况易受海洋环境的影响[1],其中,冬春生群体是我国在西北太平洋重要的商业捕捞对象[2]。中国西北太平洋鱿钓渔业始于1993年,目前,我国每年的柔鱼捕捞量稳定在10万t左右[3]。由于柔鱼类具有重要的生态性和经济性,其资源丰度时空变化及其与环境因子的关系研究越来越受到重视[4–6],研究方法包括了广义可加模型[7]和栖息地指数模型[8–12]等方法,研究认为,柔鱼的资源动态与海表面温度(Sea Surface Temperature,SST)[13–14]、叶绿素(Chlorophyll a,Chl a)浓度[15]、海表面高度(Sea Surface Height,SSH)[16]和海表面盐度(Sea Surface Salinity,SSS)[17]存在明显关系。同时,柔鱼资源丰度变化也与气候和环境因子有关,例如高雪等[18]建立了不同气候和环境因子组合的柔鱼资源丰度灰色系统预测模型,表明影响柔鱼年间资源丰度的因素主要包括了柔鱼产卵场的SST和Chl a 浓度以及太平洋年代际震荡(Pacific Decadal Oscillation,PDO)。另外,机器学习方法,如神经网络也被用于柔鱼资源丰度预测,汪金涛等[19]对比了基于频度统计和BP神经网络预测柔鱼空间分布效果的好坏。

图1 西北太平洋柔鱼冬生群体空间和洄游分布Fig.1 Spatial distribution and migratory distribution of winter cohort of Ommastrephes bartramii in the Northwest Pacific Ocean
BP神经网络模型是由Rumelhart和Mc-Celland首先提出的具有多层结构的前向神经网络[20]。BP神经网络具有非线性映射能力,其优化算法用于非线性函数拟合,可以提高拟合精度,使模型具有较强应用性[21–22];并且模型可以通过合理的自组织网络的拓扑结构,改变神经元的激活特性以及在必要时调整网络的学习参数等方法,提高了信息处理的能力[23]。但是,现有的研究仅是利用神经网络模型对柔鱼的资源丰度进行预测,并没有应用到渔场分析中。如能否利用神经网络模型提取柔鱼资源丰度的时空变化规律并用于其标准化?能否探究柔鱼资源丰度变化模式与环境因子的响应关系?为此,本研究以时空因子(年、月、经度、纬度)和与柔鱼资源丰度变化密切相关的环境因子作为输入变量,以柔鱼单位捕捞努力量渔获量(Catch Per Unit Effort,CPUE)为输出变量建立神经网络空间分布模型,估计2000−2015年柔鱼资源丰度的时空分布,分析环境因子的重要性,研究结果将为柔鱼的资源评估以及资源可持续开发与养护提供科学依据。
2 材料和方法
2.1 材料来源
中国远洋渔业数据中心收集的2000−2015年7−10月西北太平洋的渔业捕捞数据,空间范围为40°~50°N,150°~170°E,数据内容包括作业时间、作业船数、作业位置及捕捞产量,时间分辨率为天。
环境数据包括 SST、Chl a浓度、SSH和 SSS。其中,SST、Chl a浓度和 SSH数据来源于 Oceanwatch(http://oceanwatch.pifsc.noaa.gov);SSS数据来源于哥伦比亚大学环境数据中心(http://iridl.ldeo.columbia.edu),数据时间分辨率为月,空间分辨率为 0.5°×0.5°。
CPUE作为西北太平洋柔鱼的资源丰度指标,生产数据按空间分辨率0.5°×0.5°进行统计,时间分辨率按月进行统计,计算CPUE的公式为[24]

2.2 BP 神经网络模型
研究表明,海洋环境因子 SST[13–14]、Chl a 浓度[15]、SSH[16]和SSS[17]与柔鱼资源空间分布变化密切相关,因此将这4个环境因素和时空因子作为输入因子;为了确定不同环境因子的重要性,采用向前选择法将环境因子逐个加入模型中,根据拟合优度确定环境因子重要程度,构建4种模型方案(表1)。对于每种方案,随机选取70%的数据集作为训练样本集,剩下30%作为测试样本集,设定学习速率为0.1,动量参数为0.5,最大训练批次为1 000次,最大误差为0.001,隐含层节点数按经验公式选取[25],公式为
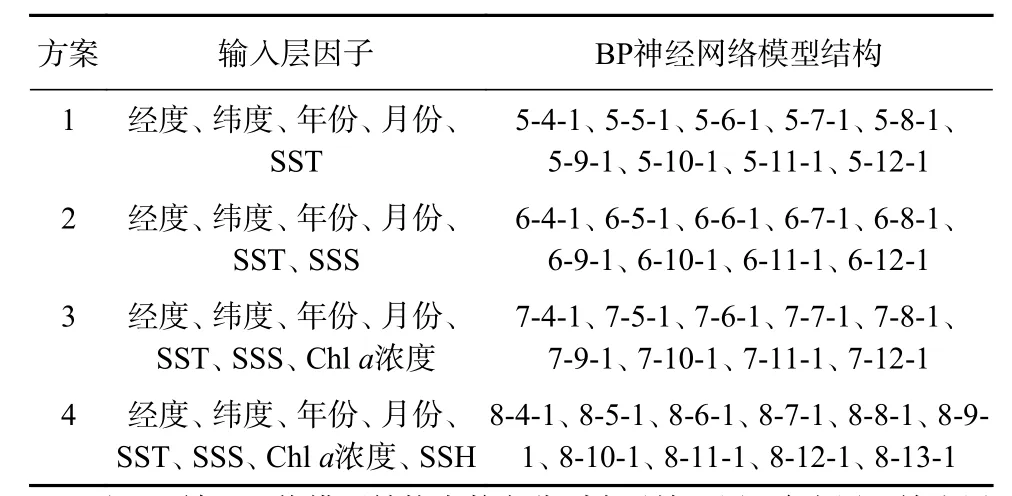
表1 西北太平洋柔鱼空间分布BP神经网络模型Table 1 Spatial BP neural network models for Ommastrephes bartramii in the Northwest Pacific Ocean

式中,n1为隐含层神经元个数;n为输入层神经元个数;m为输出层神经元个数;a为[1,10]之间的整数。
2.3 模型验证
性能指标均方误差(Mean Squared Error,MSE)最小化作为判断和选择最优模型的标准[26]。

式中,MSE为均方误差;N为样本个数;t表示第t个样本;为第t个样本输出层实际观测值;为第t个样本输出层预测值。MSE越小,BP神经网络预测的结果就越准确,BP神经网络模型越优。
2.4 柔鱼资源丰度的估计
利用建立的最优模型估计2000−2015年7−10月40°~50°N,150°~170°E 海域范围内的柔鱼资源丰度,空间平均CPUE表示月平均柔鱼资源丰度。
3 结果
3.1 最优模型
根据均方误差大小(表2,表3),方案1的最优模型结构为5-9-1,训练集样本的均方误差为0.038 9;测试集样本的均方误差为0.038 8;方案2的最优模型结构为 6-8-1,训练集样本的均方误差为 0.038 2(表 2),测试集样本的均方误差为0.038(表3);方案3的最优模型结构为7-9-1,训练集样本的均方误差为0.047 1,测试集样本的均方误差为0.041 4;方案4的最优模型结构为8-9-1,训练集样本的均方误差为0.042 6,测试集样本的均方误差为0.042 3。根据均方误差越小,模型越优原则,确定方案2为最优方案,最终模型结构为6-8-1,其训练样本集和测试样本集拟合状态如图2和图3所示,其输入层为经度、纬度、年份、月份、SST、SSS,隐含层数为 8(图 4)。

图2 西北太平洋柔鱼空间 BP 神经网络最优模型训练样本集(PCPUE)拟合状态Fig.2 Training dataset fitting of optimal BP neural network(PCPUE) for Ommastrephes bartramii in the Northwest Pacific Ocean

图3 西北太平洋柔鱼空间 BP 神经网络最优模型测试样本集(PCPUE)拟合状态Fig.3 Testing dataset fitting of optimal BP neural network(PCPUE) for Ommastrephes bartramii in the Northwest Pacific Ocean

图4 西北太平洋柔鱼最优空间 BP 神经网络模型Fig.4 Optimal spatial BP neural network model for Ommastrephes bartramii in the Northwest Pacific Ocean
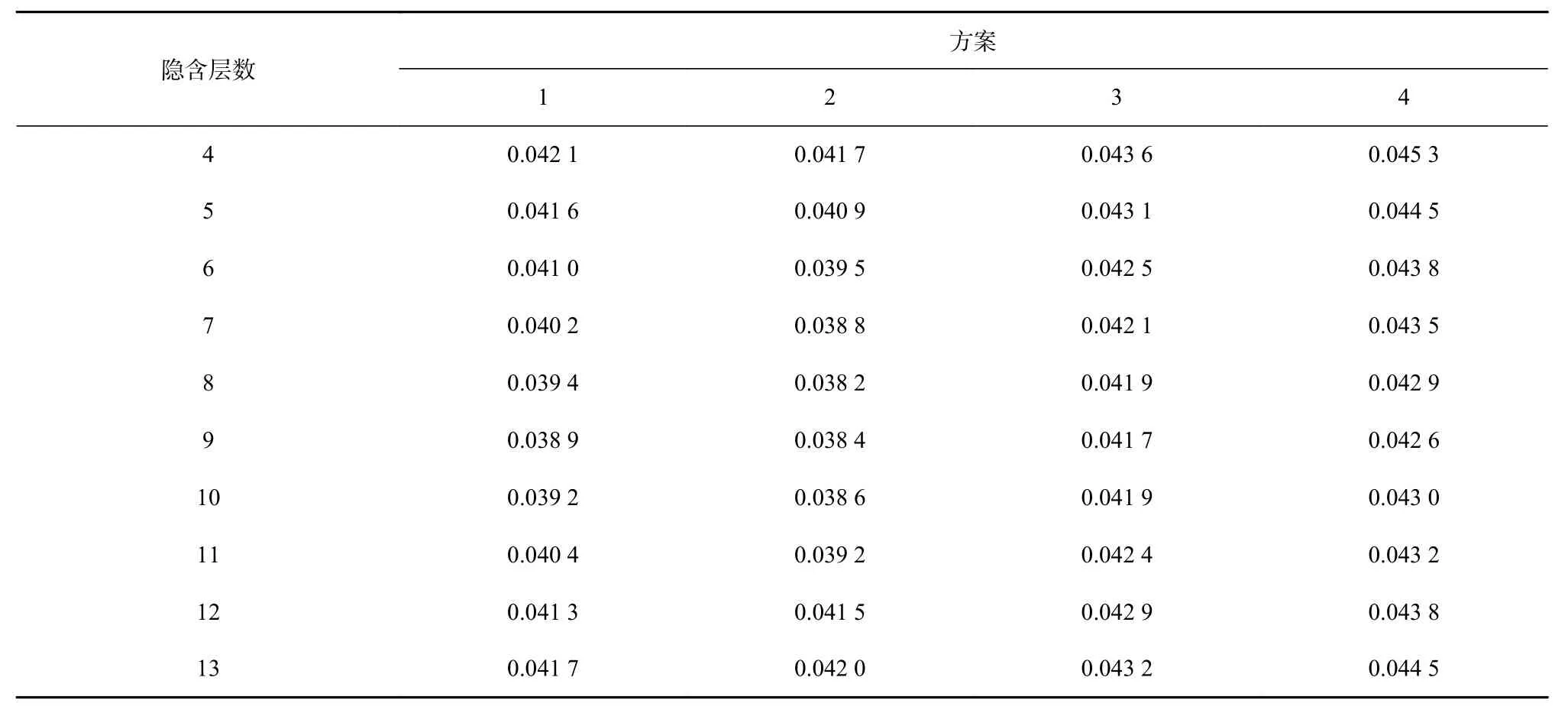
表2 不同方案不同隐含层下训练集CPUE的均方误差Table 2 Mean square error of training dataset CPUE for different scenarios
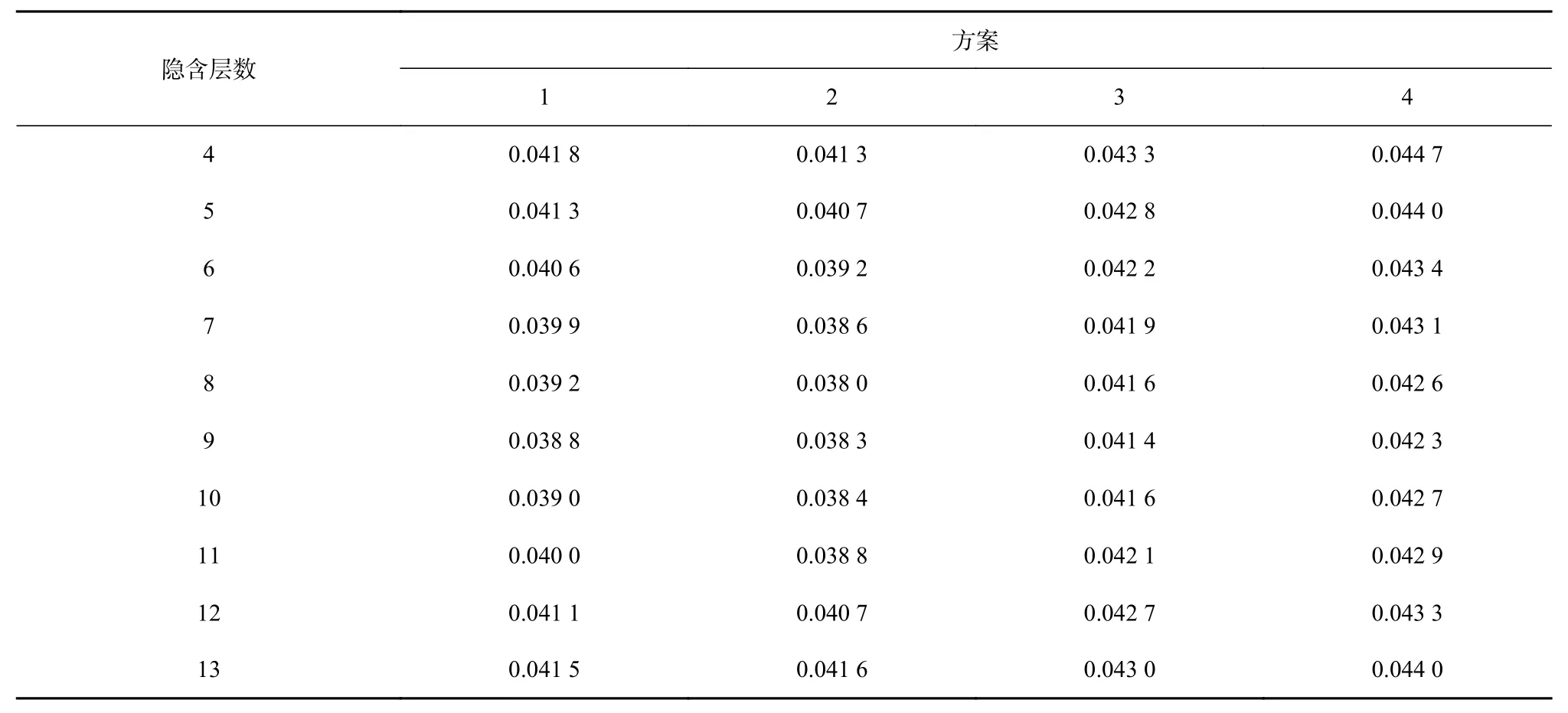
表3 不同方案不同隐含层下测试集CPUE的均方误差Table 3 Mean square error of testing dataset CPUE for different scenarios
3.2 柔鱼资源丰度空间分布
柔鱼CPUE空间分布在2000−2015年间变化明显。在8−9月,CPUE集中分布比较明显,CPUE较大值大多分布在155°~160°E范围内;各年最高CPUE纬度范围集中在 41.5°~43.5°N 范围内。2000−2010年,2000年8月、2003年8月、2004年和2005年的10月、2006年 7−9月、2007年 9月和 2008年 8月中心渔场范围较大;大部分年份的7月、10月渔场范围较为分散,并且中心渔场范围非常小。2011−2015年各月份的CPUE整体分布较少,柔鱼资源丰度小(图5,图6)。

图5 2000−2008 年西北太平洋柔鱼预测 CPUE 分布Fig.5 Simulated CPUE distribution of Ommastrephes bartramii from 2000 to 2008 in the Northwest Pacific Ocean

图6 2009−2015 年西北太平洋柔鱼预测 CPUE 分布Fig.6 Simulated CPUE distribution of Ommastrephes bartramii from 2009 to 2015 in the Northwest Pacific Ocean
3.3 柔鱼资源丰度时间变化
时间上,2000−2015年柔鱼预测CPUE呈现年间不断波动的趋势(图 7),2000−2003年柔鱼 CPUE 大致逐年增加,2004−2007年保持较高值,柔鱼资源丰度大;2008−2015年CPUE逐年波动并有减小趋势,CPUE最小值在2013年7月,为0.8 t/d,资源丰度最小;各年7−10月CPUE峰值主要出现在8月或者9月,2004年8月、2007年9月和2008年8月峰值最为明显且分别为 4.97 t/d、5.75 t/d 和 4.40 t/d。各年 7−11月平均CPUE在2007年9月最高为5.75 t/d,柔鱼资源丰度最大。
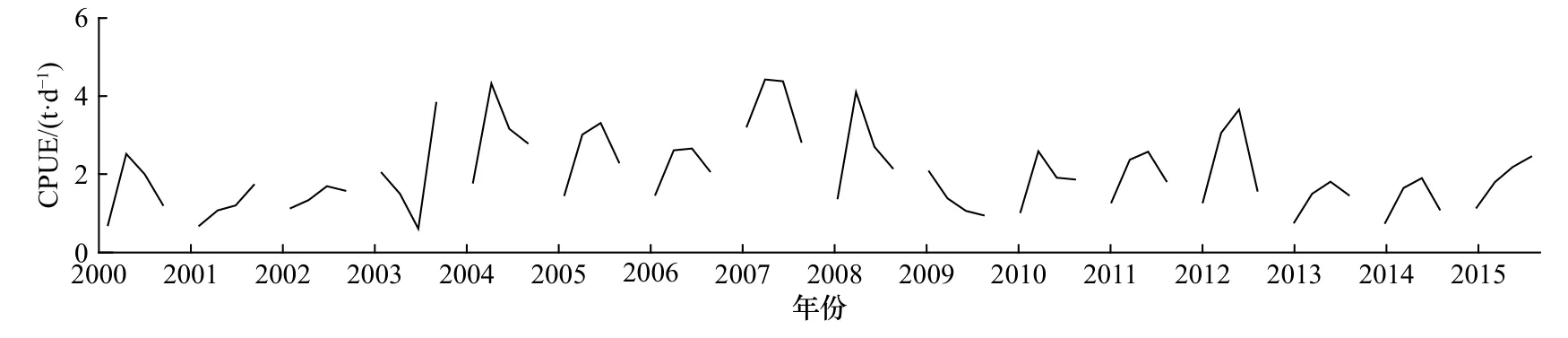
图7 2000−2015年7−10月份柔鱼资源丰度时间趋势Fig.7 Temporal trend of the abundance of CPUE of Ommastrephes bartramii from July to October during 2000 to 2015
4 讨论与分析
根据变量向前选择法和模型精度可知,环境因子SST和SSS对柔鱼的资源丰度空间分布模型的贡献率大于Chl a浓度和SSH。Alabia等[27]利用最大熵模型分析渔场发现,相比SSH、海表面风应力旋转和涡动,SSS和SST对模型的贡献更大;魏广恩[28]对柔鱼CPUE标准化的研究中也发现,SST对CPUE的偏差解释率要大于Chl a浓度,且柔鱼渔场的形成与黑潮和亲潮的动态密切相关[29];余为等[30]认为,渔场SST能够代表黑潮和亲潮强弱的变化,是影响柔鱼资源丰度动态的关键因子,SSS和SST相结合能够表征海水的水团变化。本研究结论SST和SSS对柔鱼资源丰度时空变动有较大影响与前人基本一致。
但是,渔场中SSH和Chl a浓度也反映了渔场当下水文特征(如涡流)和饵料特征[31],也能够影响柔鱼空间分布[30],在后续的研究中可以用于辅助分析。同时,气候变化也影响柔鱼的资源丰度,沈建华等[32]认为,当黑潮较弱或亲潮较强时出现的黑潮大蛇形和表温过低现象导致了2001年太平洋柔鱼产量下降;余为等[16]认为,PDO暖期时其气候条件有利于柔鱼生长和繁殖,且气候事件会对CPUE产生影响,这与本研究得到的 CPUE时间序列一致(图 6):2003−2006年处于PDO暖期,CPUE较高,2000−2002年和2009−2015年处于PDO冷期,CPUE较低。
西北太平洋柔鱼为全年产卵生殖的短生命周期物种,以月时间尺度的研究能够发现更丰富的柔鱼阶段性生态习性变化[1]。柔鱼早期幼体生活在35°N以南的黑潮逆流区,以后向北洄游,8−10月性未成熟和性成熟的柔鱼主要分布在40°~46°N亲潮前锋区及其周边海域,10月开始南下进行洄游[33]。在我国柔鱼渔业传统作业海域,8−10月冷暖水交汇,等温线密集,柔鱼渔场较密集,柔鱼丰度较高,7月柔鱼尚未洄游至捕捞海域,资源丰度较低;8月到达索饵场,资源丰度较高;10−11月柔鱼性成熟,开始南下进行产卵洄游,资源丰度减小[34],这与本研究得到的结果一致(图6);7−8月柔鱼的CPUE增加,9−11月柔鱼的CPUE减少。
CPUE作为资源丰度的指标,捕捞努力量空间分布、时空尺度的选择会对CPUE标准化产生影响[35]。人工神经网络具有很好的自学习能力和很强的泛化和容错能力[36],但BP神经网络是典型的“黑盒模型”,在解释CPUE与输入因子之间的关系上缺乏直观性[37],且在收敛速度上不足,所以学习算法和网络结构优化还有改进空间[38],今后可研究明确CPUE和环境因子在神经网络模型中具体关系。
本文利用柔鱼渔业数据和其渔场环境数据,构建了BP神经网络空间分布模型,拟合了柔鱼资源丰度,发现神经网络模型能够模拟柔鱼资源丰度的时空变化,探究不同环境因子对柔鱼资源丰度时空变化的重要性,并可用于资源丰度标准化,结果可为柔鱼生产和管理提供参考。
猜你喜欢
现代电力(2022年2期)2022-05-23
科技风(2021年19期)2021-09-07
趣味(作文与阅读)(2021年5期)2021-08-19
小哥白尼(军事科学)(2020年12期)2021-01-18
今日中国·法文版(2020年7期)2020-07-04
电子制作(2019年19期)2019-11-23
电子制作(2019年24期)2019-02-23
科学中国人(2017年35期)2017-06-08
哈尔滨理工大学学报(2016年3期)2016-11-05
探测与控制学报(2015年4期)2015-12-15
