
长臂虾科几种重要经济虾类的繁殖生物学研究进展
2021-08-10 09:06:26胡润豪史文军万夕和王李宝杨泽禹吴旭干
海洋渔业 2021年4期
胡润豪,史文军,王 盼,万夕和,沈 辉,黎 慧,王李宝,杨泽禹,吴旭干
(1.上海海洋大学,水产科学国家级实验教学示范中心,上海 201306;2.江苏省海洋水产研究所,江苏南通 226007;3.中国科学院烟台海岸带研究所,山东烟台 264003)
长臂虾科(Palaemonoidea)隶属甲壳纲(Crustacea)十 足 目(Decapoda)真 虾 次 目(Caridea),其下属长臂虾亚科现生种属共计21属373种,隐虾亚科现生种属共计117属604种[1]。常见长臂虾科虾类主要隶属于长臂虾亚科的沼虾属(Macrobrachium)或白虾属(Exopalaemon),其余各属虽在我国也有分布,但群体数量较小或未实现大规模人工培育,如古洁虾属(Palaemonetes)的广东古洁虾(P.tonkinensis)、瘦虾属(Leander)的细脚瘦虾(L.tenuicornis)等[2]。
长臂虾科虾类是我国渔业重要的一类经济虾类[2],主要的养殖或捕捞品种有:日本沼虾(M.nipponense)、罗氏沼虾(M.rosenbergii)、脊尾白虾(E.carinicauda)、秀丽白虾(E.modestus)、葛氏长臂虾(Palaemon gravieri)、海南沼虾(M.hainanense)、细螯沼虾(M.superbum)以及安氏白虾(E.annandalei)[3-4]等,其中养殖规模较大且研究报道较多的有淡水品种日本沼虾、罗氏沼虾和海水品种脊尾白虾。日本沼虾自然分布于亚洲各地,除中国外,日本、朝鲜半岛、越南、马来西亚以及俄罗斯远东地区都有分布[5];罗氏沼虾原产于泰国、印度、缅甸等的河口处,自1976年引入我国后,分别于广东和广西的水产研究所进行试养[6]。2018年我国日本沼虾产量23.4万t、罗氏沼虾产量13.3万t,在淡水养殖虾类产量排行中分别位列第三和第四位,江苏省日本沼虾与罗氏沼虾产量为全国最高,分别为11.5万t和6.4万t[7]。脊尾白虾主要分布于中国大陆沿岸和朝鲜半岛西岸的浅海低盐水域[5],目前全国养殖面积约3.7万hm2,其中江苏省养殖面积最大、产量最高。
1 繁殖生物学特征
1.1 繁殖期
长臂虾科几种重要经济虾类的繁殖期[8-20]见表1。长臂虾科虾类(以下简称长臂虾类)繁殖期较长,一般在5个月以上,繁殖盛期一般在5—8月份,但不同地区的长臂虾类繁殖期和繁殖盛期存在一定差异,这可能主要是由于温度和生物饵料量等因素的差异导致。
1.2 繁殖力
产卵量和产卵次数是虾类繁殖力的主要体现,长臂虾类产卵量从几百粒到几万粒不等,产卵次数也存在较大差异。长臂虾类产卵量不仅受 到 卵 巢 发 育 状 况[11]、体 长[15,21-22]和 体 质量[23-24]等内在因素影响,也容易受到外界环境因素的影响,如温度[13]、盐度[25]和季节变化[26]等。多数长臂虾类一年内可以多次产卵[15,22,27]。长臂虾类产卵量一般少于对虾类而多于螯虾类(除秀丽白虾外)[18,20,23,26,28-34],这可能是由于雌虾个体大小以及卵粒直径差异造成的(表2)。

表2 长臂虾科虾类与其他科虾类产卵量及卵径对比Tab.2 Comparison of spawning number and egg diameter of Palaemonoidea and other families
1.3 雌雄性比
雌雄性比是动物群体构成的重要评价指标,也是繁殖生物学关注的群体特征之一,长臂虾科几种重要经济虾类雌雄性比见表3。卢敬让等[21]认为,繁殖季节雌虾多于雄虾是因为抱卵雌虾繁殖洄游,以及越冬期间雄虾死亡率高于雌虾,从而导致越冬后群体中雌虾占比较大,这与何绪刚等[8]和孙建贻等[35]对日本沼虾性比变化的研究结果一致。另外,在长臂虾类繁殖期,部分成虾寿命已经达到上限,群体中雄虾在交配完成生物学使命后很快死亡,雌虾则要等待受精卵孵化后死亡[8,36];而雌虾死亡后,大量当年幼虾加入群体,缓冲了雌虾死亡导致的性比变化趋势。与其他野生长臂虾类群体相比,引进物种罗氏沼虾主要为池塘养殖,虽然雌雄性比均大于1,但相对变化较小,这可能是由于自然群体的捕捞在时间和空间上跨度较大,导致数据统计的波动性大,而池塘养殖时间和空间跨度小,数据统计的波动较小。
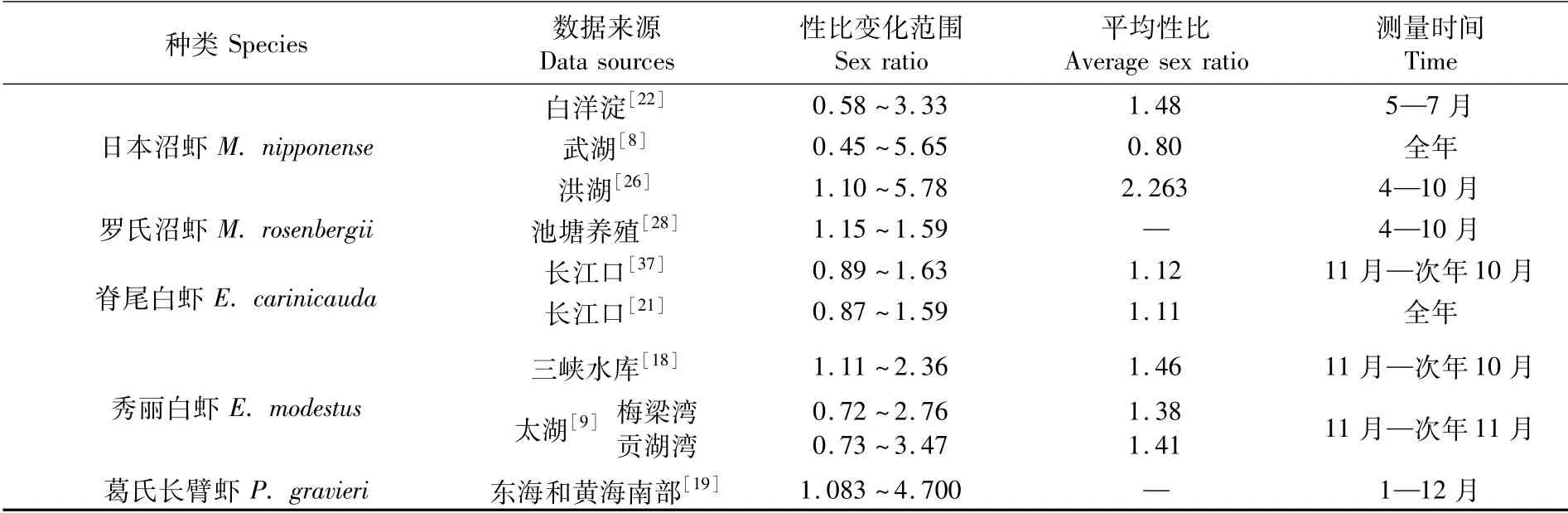
表3 长臂虾科几种重要经济虾类雌雄性比Tab.3 Sex ratio of some important economic kinds of Palaemonidea
2 生殖系统发育
2.1 雌性生殖系统
2.1.1 雌性生殖系统结构
长臂虾类雌性生殖系统组成基本相似,都有卵巢与输卵管,但不同种之间存在一定差异,主要表现在卵巢左右两叶的连接方式上[36],目前还未见关于这种结构差异可能导致的影响的报道。脊尾白虾雌性生殖系统由1对卵巢和2根输卵管组成,整个卵巢呈“O”字型,前后端愈合在一起,中 间 形 成 一 个 空 隙[36],与 粗 糙 沼 虾(M.asperulum)[38]卵巢相似。罗氏沼虾卵巢左右两叶呈桑葚状,前端愈合,后端分离,卵巢两侧分别有短的输卵管与生殖孔相连[39-40],与哈氏仿对虾(Parapenaeopsis hardwickii)[41]卵巢相似。日本沼虾的卵巢呈椭圆形,前端略尖,后端圆钝,前部成对,后部合二为一,在卵巢的两侧各有一条短而直的输卵管[42],与克氏原螯虾(Procambarus clarkii)[43]的卵巢结构类似。
2.1.2 卵巢发育
研究人员主要根据卵巢颜色、大小或相对位置及内部卵细胞成熟情况将日本沼虾[42]和脊尾白虾[15]的卵巢发育分为5个时期,罗氏沼虾[40]的卵巢发育分为4个时期,见表4。总结3种长臂虾类卵巢各时期的特点,笔者认为脊尾白虾卵巢5个发育时期的划分和命名更为合理。

表4 3种长臂虾类卵巢发育时期及特征Tab.4 Ovarian development stages and characteristics of three species of Palaemonidae
2.2 雄性生殖系统
2.2.1 雄性生殖系统结构
长臂虾类雄性生殖系统的研究主要集中在精巢结构、精子发生等方面。长臂虾类的雄性生殖系统由精巢和输精管组成[44-45],与对虾类雄性生殖系统存在明显差异,没有分泌管和精囊,而这两种结构往往在对虾生殖系统中发挥重要作用[47](图1)。精巢内为无数的生精小管,其管壁由位于基膜上的生精上皮和管壁上皮构成,生精上皮细胞主要为精原细胞,经有丝分裂,一部分精原细胞发育成不同阶段的雄性生殖细胞,形成生殖带[45]。同一区段的生精小管内精子发生的同步率较高,可以保证在较短时间内产生出大量成熟精子;不同区段的精子的发生不同步,使得生殖带能在较长时期内源源不断地产生精子,适应于当年生雄虾繁殖期的多次繁殖[46]。长臂虾类输精管末端膨大为壶腹,肌肉层发达,可以暂时贮存包裹着精子的精荚,同时发达的肌肉层能在射精时给予所需的力量,其作用可能类似于对虾的精囊,起到储精和射精的双重功效[44-46]。
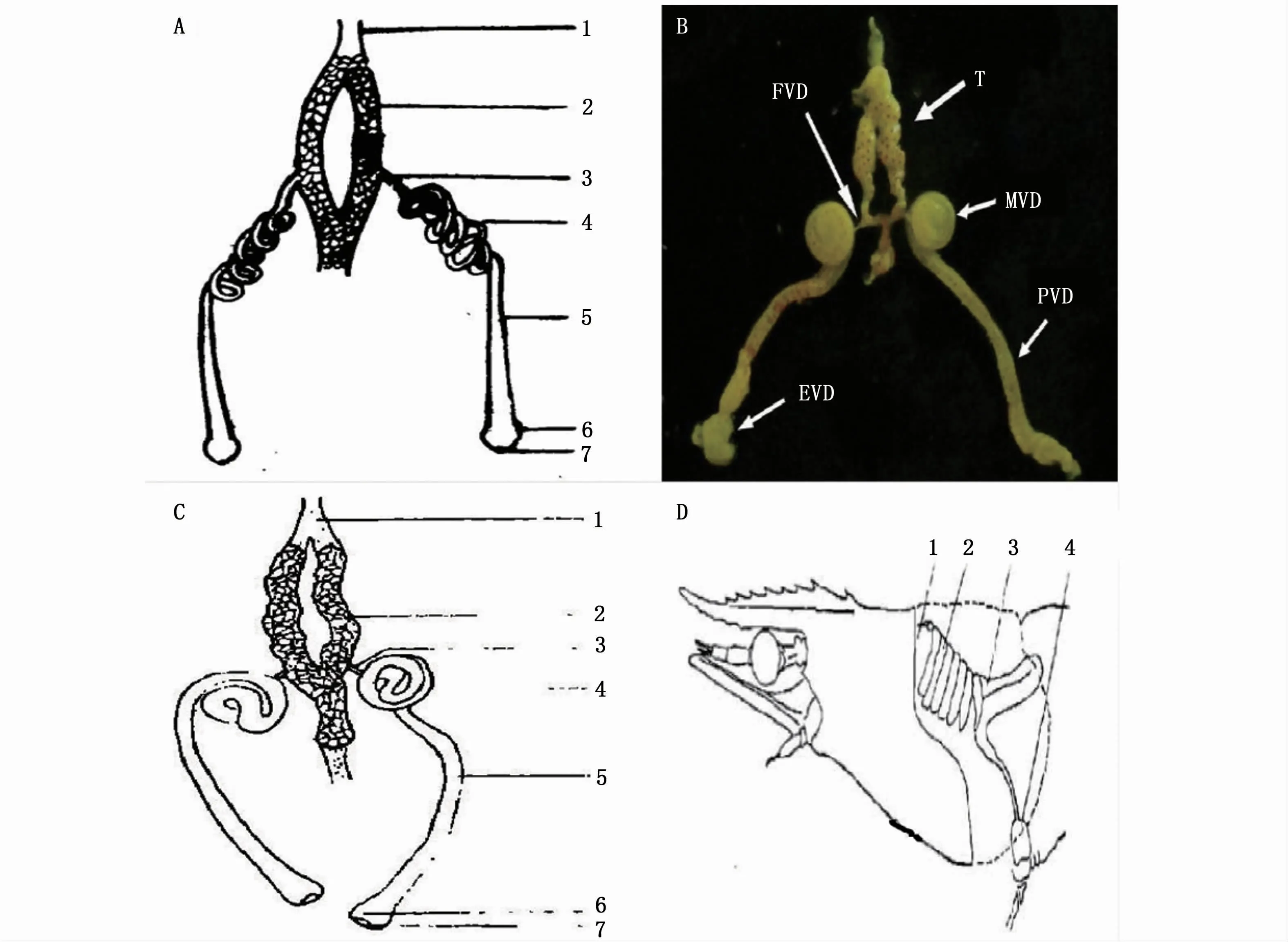
图1 日本沼虾[45](A)、脊尾白虾[44](B)、秀丽白虾[46](C)、中国明对虾[47](D)雄性生殖系统Fig 1 Male reproductive system of M.nipponense[45](A),E.carinicauda[44](B),E.modestus[46](C)and Fenneropenaeus chinensis[47](D)
2.2.2 精子发生
长臂虾类的精荚呈索状,由中央的精子群、粘液团及包被在其外的精荚壁组成[48-49]。早期的精荚主要由精荚基质构成,由输精管上皮分泌并逐渐积累到精子团外部;随后输精管上皮分泌形成精荚壁,最终产生完整精荚[50]。日本沼虾、脊尾白虾、秀丽白虾的精荚形成过程见表5。

表5 3种长臂虾类的精荚形成Tab.5 Spermatophore formation of three species of Palaemonidae
3 胚胎和幼体发育
3.1 胚胎发育过程
胚胎发育过程是繁殖生物学重要的研究内容,胚胎发育过程中,卵粒的大小、体积和含水量随胚胎发育而增加。长臂虾类胚胎发育过程基本一致,以雌雄虾抱对受精产生的受精卵为起始,卵内仅1个细胞;卵裂期受精卵内的细胞开始分裂,形成分裂球;细胞经过第8次分裂数量增加到256个,进入囊胚期,此时细胞核全部移近受精卵表面,构成囊胚层;受精卵动物极细胞密集,植物极内陷形成原口,标志着原肠期的开始;胚胎卵黄含量减少,进入前无节幼体期,两对触角原基和一对大颚原基的形成是前无节幼体期的主要事件;后无节幼体期,形成两对颚足原基和两对小颚原基;进入前溞状幼体期的胚胎,透明区内出现一对由复眼原基内色素细胞分泌的黑色颗粒状物;复眼原基形成体积更大的椭圆形复眼,此时称为溞状幼体期或后溞状幼体期,胚胎头胸部13对附肢发育基本完成,随后幼体破膜而出,胚胎期结束[51-54]。3种虾类的胚胎发育的划分方法见表6。

表6 胚胎发育分期Tab.6 Stages of embryonic development
目前根据胚胎发育的进程,长臂虾类胚胎发育分期主要有7期法[52]、8期法[51,53]和12期法[54]。12期法对卵裂期的划分过于详细、工作量大且实用性较低;7期法以受精卵卵裂开始为胚胎发育第1期,这与事实不相符。与前两者相比,8期法更加简洁且实用,其第6期命名为“后无节幼体期”,与第5期“前无节幼体期”可以鲜明对比;第8期命名为“膜内溞状幼体期”,相比“后溞状幼体期”的命名更能概括这一时期的特征。
3.2 卵裂方式
几种重要长臂虾类的卵裂方式基本一致,但与对虾科、螯虾科虾类(以下简称对虾类、螯虾类)的卵裂方式存在差异,3科虾类的卵裂方式及区别见表7[51-56]。卵裂方式主要受到卵黄含量影响,3科虾类的卵主要属于中黄卵(日本沼虾为多黄卵),但实际卵黄含量仍存在差异:对虾类单次产卵量大,卵黄含量较少,分裂面可将卵完全分割开;长臂虾类产卵量较少,卵黄含量较多,分裂面不能将卵完全分割开;螯虾类产卵量极少,卵黄含量较多,卵裂以不完全卵裂为主。此外,相关文献虽未明确指出脊尾白虾的卵裂方式,但结合日本沼虾和罗氏沼虾卵裂方式的描述及笔者的观察研究,认为脊尾白虾卵裂方式应属于完全卵裂和不完全卵裂之间的过渡类型。
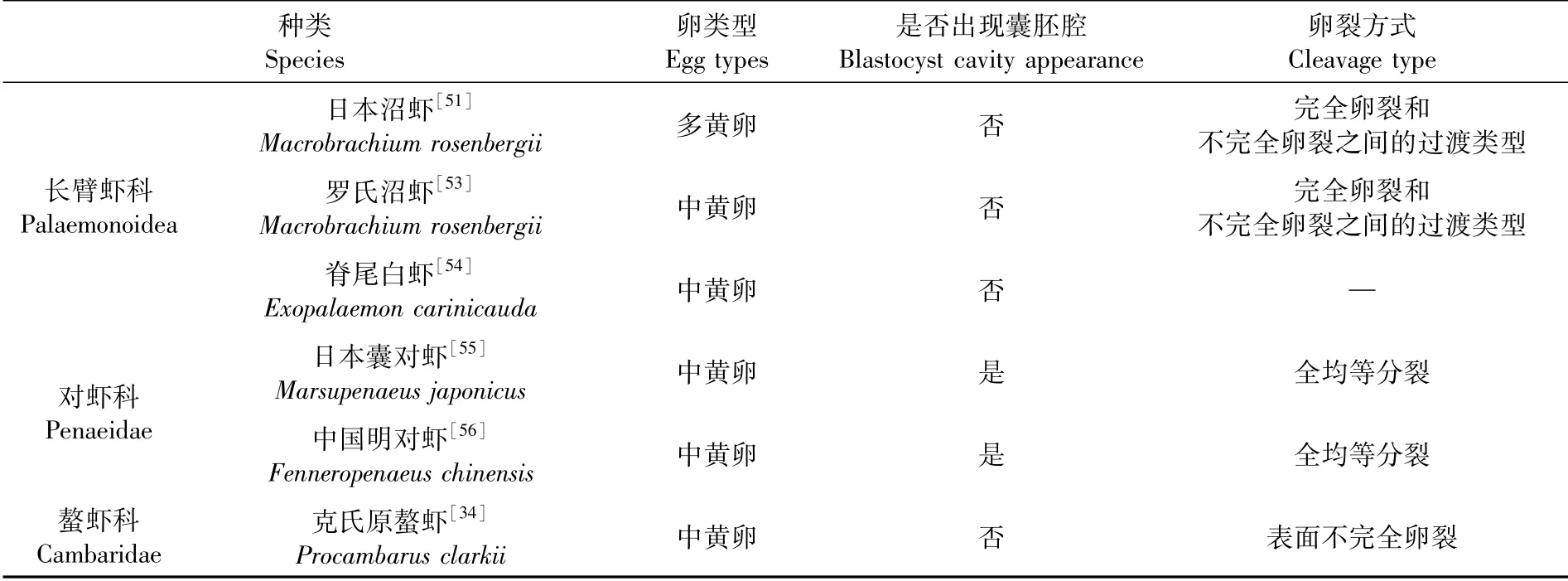
表7 长臂虾类、对虾类、螯虾类的卵裂方式Tab.7 Cleavage patterns of Palaemonidae,Penaeidae and Cambaridae
3.3 幼体发育过程
不同种虾类在胚胎内度过的时间长短不同,这导致幼体出膜形态以及后续发育进程呈现出不同的类型。对虾类在无节幼体期前就破膜而出[57],长臂虾类无节幼体期在胚胎中度过,出膜的幼体一般是溞状幼体[58]。长臂虾类刚出膜的溞状幼体运动能力较弱,自然条件下,早期溞状幼体为植食性,主要滤食各种藻类来获取营养,人工养殖条件下会投喂卤虫无节幼体等为其提供营养[58]。虾类的幼体发育为变态发育,不同时期的幼体之间形态特征差异较大,幼体的形态特征及生活习性与成虾有很大不同[36]。日本沼虾幼体发育存在中间蜕皮现象,从Z5(经过4次蜕皮后的溞状幼体)开始,凡需蜕二次皮方可完成一期形态递进的幼体,其第一次蜕皮后,第二触角外鞭形态无递进变化,幼体形态、大小变化不大,即为中间蜕皮状态,则需再蜕皮一次[58];其他长臂虾类中未见相关报道,因此日本沼虾蜕皮次数通常高于其他长臂虾类。罗氏沼虾、日本沼虾与脊尾白虾的幼体发育过程见表8[58-62]。研究人员对脊尾白虾的蜕皮次数和发育时间存在2种观点[60-61],这主要是由于对进入仔虾期的判定不同,一种观点认为幼体需经过6次蜕皮才能发育完好进入仔虾期[60];而另一种观点则认为经5次蜕皮后,幼体即发育完好进入仔虾期[61]。

表8 长臂虾类幼体发育过程Tab.8 Larval development of Palaemonidae
3.4 环境因子对胚胎和幼体发育的影响
环境因子对长臂虾类幼体孵化、发育、存活等有很大的影响[63-66]。温度对于水生生物生长和存活有明显影响,在适温范围内,温度越高,水产动物胚胎与幼体发育速率越快,生长速度亦相应加快。当环境温度超过动物自身的调节能力,例如,高于适温范围时,其代谢强度加大,能量无法积累或已经积累的能量被消耗;低于适温范围时,其代谢活动降低,都会导致生长速率减慢[63-66]。徐慈浩等[53]在对罗氏沼虾的研究中指出,温度的升高有利于加快胚胎发育的速度,但是过快的发育速率可能导致胚胎结构分化不够完整,最终影响罗氏沼虾幼体的孵化率和孵化质量。脊尾白虾各期幼体在16~36℃范围内,温度越高发育速度越快,发育持续时间越短,幼体体长日增长量越大[67]。日本沼虾[68]、罗氏沼虾[69]的幼体发育也受温度影响,这种影响主要是通过影响虾类幼体有效积温的累计来影响幼体发育的速率。
盐度是影响长臂虾类胚胎和幼体发育的另一重要环境因子。脊尾白虾胚胎发育时间与发育时的盐度成正相关[70]。通常虾类胚胎和幼体对盐度变化更为敏感[71],罗氏沼虾幼体发育的适宜盐度为10~14[69],成体则能在盐度0~40的水中生存[72]。日本沼虾幼体在盐度为12时生长速度最快,成活率最高[73]。梁俊平等[74]认为,不同盐度下(2~30)脊尾白虾溞状幼体的变态率和仔虾存活率均无显著差异,但盐度15和20下仔虾干质量显著大于其他盐度组,盐度对仔虾个体干质量影响显著。
水体环境的pH是虾类繁殖的重要环境因素之一,最为直观的是影响幼体的存活率,过高或者过低的pH值都会降低幼体的存活率[69,75]。浦蕴惠[76]报道,脊尾白虾胚胎适宜的pH在7.0,碱性环境对胚胎的影响更大,胚胎更适宜偏酸性的环境。pH值的变化能够引起虾体内酶活性的改变,从而影响虾的各种生理活动,幼体处于身体迅速发育、各种组织逐渐形成与成熟的状态,pH值变化的影响更加明显。KAWAMURA等[77]研究了低pH水环境对罗氏沼虾幼体触觉的影响,指出较低pH值下,幼体触觉刚毛中的甲壳素可能被降解,表面机械感受器被抑制。
此外,硫代硫酸钠脱氯的自来水饲养罗氏沼虾幼体会导致其存活时间缩短,发育迟缓[78];随着饲料中蛋白含量的升高,斑节对虾增重率先升高后下降,但不影响存活率[79],而饲料中添加β-胡萝卜素能够促进脊尾白虾的生长,β-胡萝卜素浓度为0.5 g·kg-1时脊尾白虾的生长效果最佳[80]。
4 繁殖相关调控基因
长臂虾类的繁殖过程受到多种基因调控,包括调控性腺发育、性别分化、胚胎发育、幼体发育等生理过程的基因,已见报道的长臂虾类中发现的参与繁殖过程调控的基因见表9[82-87,90-107]。

表9 长臂虾类繁殖相关调控基因Tab.9 Regulation genes related to reproduction of Palaemonidae
性腺发育相关基因的研究报道较多,传统繁殖生物学观点认为,甲壳纲动物的性别决定是由性染色体主导的,成熟的性腺能够分泌各种性激素作用于身体中的其他组织,从而引起非生殖组织产生性别差异。最近的研究否定了这种观点,研究人员发现中枢神经系统性别分化的出现早于性激素的分泌[81],这说明在发育初期,性别分化由基因单独决定的,随后激素开始分泌,性别分化才会逐渐受到激素的影响。目前已发现如卵巢特 异 基因[79]、叉头盒蛋 白L2基 因[83]、vasa[84]等能够影响性腺发育,参与虾类发育的性别分化。
胚胎与幼体的生长速率以及存活率也是虾类繁殖能力的重要反映。这一过程受多种基因调节,目前在长臂虾科已见报道的EcR[85]、WNT4[86]、TRY[87]等都对胚胎发生、幼体生长或者这两者的发育有着调控作用。幼体生长过程中,需要经历多次蜕皮,蜕皮速率一定程度上会影响生长速率。无脊椎动物缺乏真正的抗体和特异性的免疫细胞,机体防御反应依靠非特异的固有免疫结构[88],因此,虾类仅能通过被动的先天免疫抵御外界影响,幼体期发育经过多次生长蜕皮,蜕皮时幼体先天性免疫能力减弱,且幼虾身体结构发育不成熟,对外界影响耐受能力较弱,免疫能力是幼体生长尤其蜕皮时期存活率的关键影响因素之一。一些与长臂虾类免疫功能相关的基因可能也在幼体发育过程中发挥作用,如α2-巨球蛋白(alpha2-macrogloblin,α2M)、C-型凝集素(C-type lectins,CTL)[89]等,这些基因对幼体生长期间免疫功能的影响有待进一步研究。
5 总结与展望
长臂虾科几种重要经济虾类的繁殖期一般为5个月以上,繁殖盛期一般在5—8月份,一般有多次繁殖盛期。繁殖力方面,罗氏沼虾的单次产卵量最高,而秀丽白虾单次产卵量较少。大部分长臂虾类自然群体年平均雌雄性比高于1,但波动较大。
长臂虾类胚胎发育过程包括多种生理活动,将长臂虾类的胚胎发育过程分为8个时期的分期方法较为合理。长臂虾类孵出的溞状幼体需发育至Ⅲ期溞状幼体方具备运动捕食的能力,因此该阶段需注意保证幼体的营养供应以及提供合适的开口饵料。胚胎和幼体发育过程受到温度、盐度、pH等多种环境因子的影响,提供适宜的环境条件能够显著提高胚胎孵化率、幼体存活率以及生长速率等。
多种基因参与调控长臂虾类繁殖过程,其中一些基因或参与早期的性别分化,或参与性腺的发育。长臂虾类雌雄个体的大小、生长速率存在明显差异,通过研究性别分化基因,使得人工控制种群性别成为可能,单性化养殖可能是性别分化基因研究未来的发展方向。
目前养殖规模较大且经济价值高的日本沼虾、罗氏沼虾和脊尾白虾等已有较为全面的繁殖生物学研究。秀丽白虾、葛氏长臂虾、海南沼虾、细螯沼虾等有一定的产量,但主要依靠野外捕捞,且相关研究较少。繁殖相关基因与繁殖过程的速率与成功率密切相关,同时其研究也是性成熟、胚胎与幼体发育、生长蜕皮等过程的重要理论基础。
猜你喜欢
当代水产(2022年8期)2022-09-20 06:47:02
学苑创造·A版(2021年10期)2021-10-30 10:34:43
农民致富之友(2020年27期)2020-09-12 14:07:33
当代水产(2020年3期)2020-06-15 12:02:58
农家之友(2018年7期)2018-01-31 15:01:00
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15 12:47:46
世界热带农业信息(2014年11期)2015-01-05 18:01:03
应用海洋学学报(2014年4期)2014-11-22 07:43:48
现代农业科技(2009年17期)2009-03-08 01:47:48
