
北盘江光唇裂腹鱼种群基因组DNA遗传多样性的AFLP分析
2021-08-10 09:06代应贵安丹丹韩虎峰
海洋渔业 2021年4期
杨 伟,代应贵,2,安丹丹,韩虎峰
(1.贵州大学动物科学学院,贵阳 550025;2.贵州大学特种水产研究所,贵阳 550025)
光唇裂腹鱼(Schizothorax lissolabiatus),隶属于鲤形目(Cypriniformes)鲤科(Cyprinidae)裂腹鱼亚科(Schizothoracinae)裂腹鱼属,分布于元江、澜沧江、怒江和珠江水系南北盘江等地[1-2],为产地名优野生经济鱼类。近年来,光唇裂腹鱼栖息地遭到破坏,人类活动对其造成了不利影响[3]。
遗传多样性通常是指种内的遗传多样性,即种内个体之间或一个群体内不同个体遗传变异的总和[4]。遗传多样性是评价物种资源状况的一个重要指标,是物种适应周围环境变化、维持生存和进化的物质基础[5]。以DNA为模板的分子标记技术,通过对物种基因的扫描,能够直接反映DNA序列的差异(多态性),具有稳定、高效和准确等优点而被广泛用于物种资源评价、遗传变异规律分析、辅助育种等[6-8]。AFLP分子标记结合了RFLP技术的可靠性和PCR技术的高效性,多态性强,分辨率高,获得信息量大,用较少的引物组合即可获得准确的遗传信息,尤其适用于研究背景模糊、材料来源广泛等物种遗传资源的标记分析[8]。该技术现已被广泛应用于动植物的遗传图谱构建、种质鉴定、遗传多样性分析等研究[8-10]。
北盘江是珠江水系西江上源红水河的主要支流。目前,光唇裂腹鱼在北盘江仅见于上游可渡河[3]。本研究通过对北盘江光唇裂腹鱼种群全基因组DNA进行AFLP分析,以期揭示该种群遗传结构及遗传多样性的现状,为开展其野生资源保护和种质资源开发利用提供参考依据。
1 材料与方法
1.1 样品的采集
本研究所用试验鱼于2010年7月采自珠江水系北盘江可渡河自杨柳至都格河段,用刺网每隔7 km捕捞7~8尾光唇裂腹鱼,共计30尾鱼。试验鱼体长108~258 mm,体质量23.6~273.0 g。取鱼体背部肌肉用无水乙醇保存并带回实验室备用。
1.2 模板DNA的提取
1.3 AFLP分析
1.3.1 引物接头与引物组合信息
本次实验所用的引物组合由作者自行设计并委托上海生物工程技术服务有限公司合成,经过初筛,选用8个引物组合对北盘江光唇裂腹鱼种群全基因组DNA进行AFLP分析(表1)。
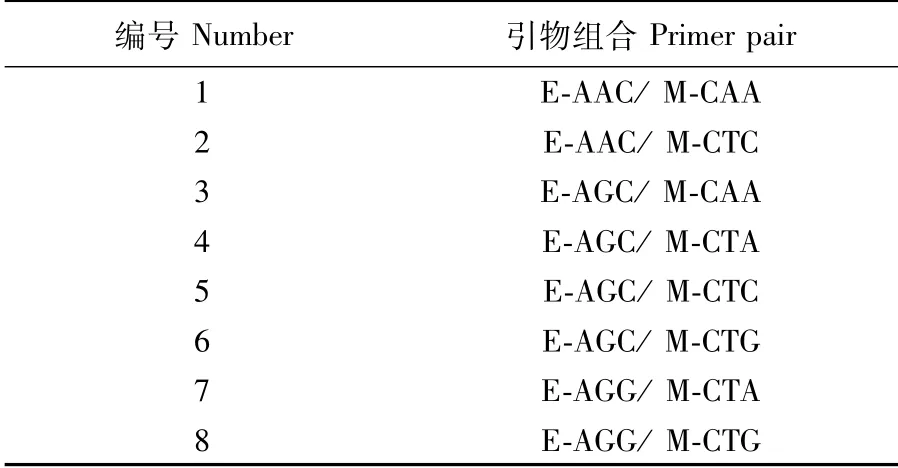
表1 北盘江光唇裂腹鱼种群AFLP分析所用引物组合Tab.1 Primer pairs used in AFLP analysis for Schizothorax lissolabiatus population from the Beipan River
1.3.2 AFLP分析
反应体系中所用的内切酶为EcoRI和MseI。酶切与连接反应同时进行。预扩增反应体系共计25μL,包括模板DNA 2μL,Pre-ampmix 1μL,dNTPs 1μL,10×PCR buffer 2.5μL,Taq酶0.5 μL,ddH2O 18μL,PCR反应程序:94℃2 min;94℃30 s,56℃30 s,72℃80 s(30个循环);72℃5 min。将预扩增产物1∶20稀释后作为选择性扩增模板,选择性扩增反应体系共计25μL,包括2μL稀释后的预扩增产物,双蒸水17.5μL,10×PCR buffer 2.5μL,EcoRI引物1μL,MseI引物1μL,dNTPs 0.5μL和Taq DNA聚合酶0.5μL,PCR反应程序:95℃预变性5 min;95℃变性30 s,50~60℃退火45 s,72℃延伸1 min,32个循环;72℃终延伸10 min,12℃冷却10 min。PCR产物4℃保存。
采用贝克曼库尔特公司CEQ8000遗传分析系统,运用荧光标记技术和毛细管电泳分离技术,将PCR产物在毛细管中电泳分离,以激光激发荧光采集数据,电泳结果通过软件自动统计分析并转换成“0、1”矩阵,保存于EXCEL表格供数据处理。
1.4 数据处理
利用POPGENE3.2软件统计位点总数、多态位点数、多态位点比例(P),并计算观测等位基因数(Na)、有效等位基因数(Ne)、Nei遗传多样性指数(H)、Shannon信息指数(I)。用NTSYS2.10软件计算30尾个体间遗传相似性系数(S),根据NEI和LI[11]的方法计算遗传距离:D=1-S。基于个体间遗传距离矩阵,采用Mega6软件构建种群个体的UPGMA、NJ系统树。用Excel表统计显性位点数据,进行显现基因型频率构图分析。
2 结果与分析
2.1 AFLP扩增结果
北盘江30尾光唇裂腹鱼中,每个引物组合扩增片段数在8~73之间,扩增片段长度为69~500 bp(图1),个体基因型数为28~30个、平均29.5个(表2)。

图1 引物组合8扩增的北盘江光唇裂腹鱼种群DNA指纹图谱Fig.1 Electrophoretogram of PCR products by AFLP primer pair 8 in Schizothorax lissolabiatus population from the Beipan River
2.2 种群遗传多样性
北盘江30尾光唇裂腹鱼总计扩增出975个有效位点,其中多态位点数897个,多态位点占总位点数的比例为92.00%(表3)。每个引物组合总扩增位点数为98~151个,平均121.88个;扩增多态位点为93~140个,多态位点比例为79.67%~95.76%,每个引物组合平均多态位点比例为91.23%。8个引物组合扩增共有位点数为5~25个,平均10.88个。
3.流程设计缺乏严谨性。内部控制体系的核心设计师流程设计,企业根据在运营过程中所遇到的风险,制定科学合理有效的控制活动,这有利于企业设计合理的可以预防风险的业务流程。但是,目前我国公立医院的运营现状来看,医院内部各部门之间权责不明、权责一体化。公立医院为了方便业务的执行,使得各部门之间未能相互制约,导致医院在业务执行过程中工作人员有徇私舞弊的现象出现。所以,医院在经营发展过程中要完善流程设计,根据实际所遇到的风险制定科学有效的控制活动。
基于8个引物组合的扩增结果,北盘江光唇裂腹鱼种群观测等位基因数(Na)、有效等位基因数(Ne)、Nei遗传多样性指数(H)和Shannon信息指数(I)平均值分别为1.912 3、1.369 0、0.227 1和0.357 5(表3)。
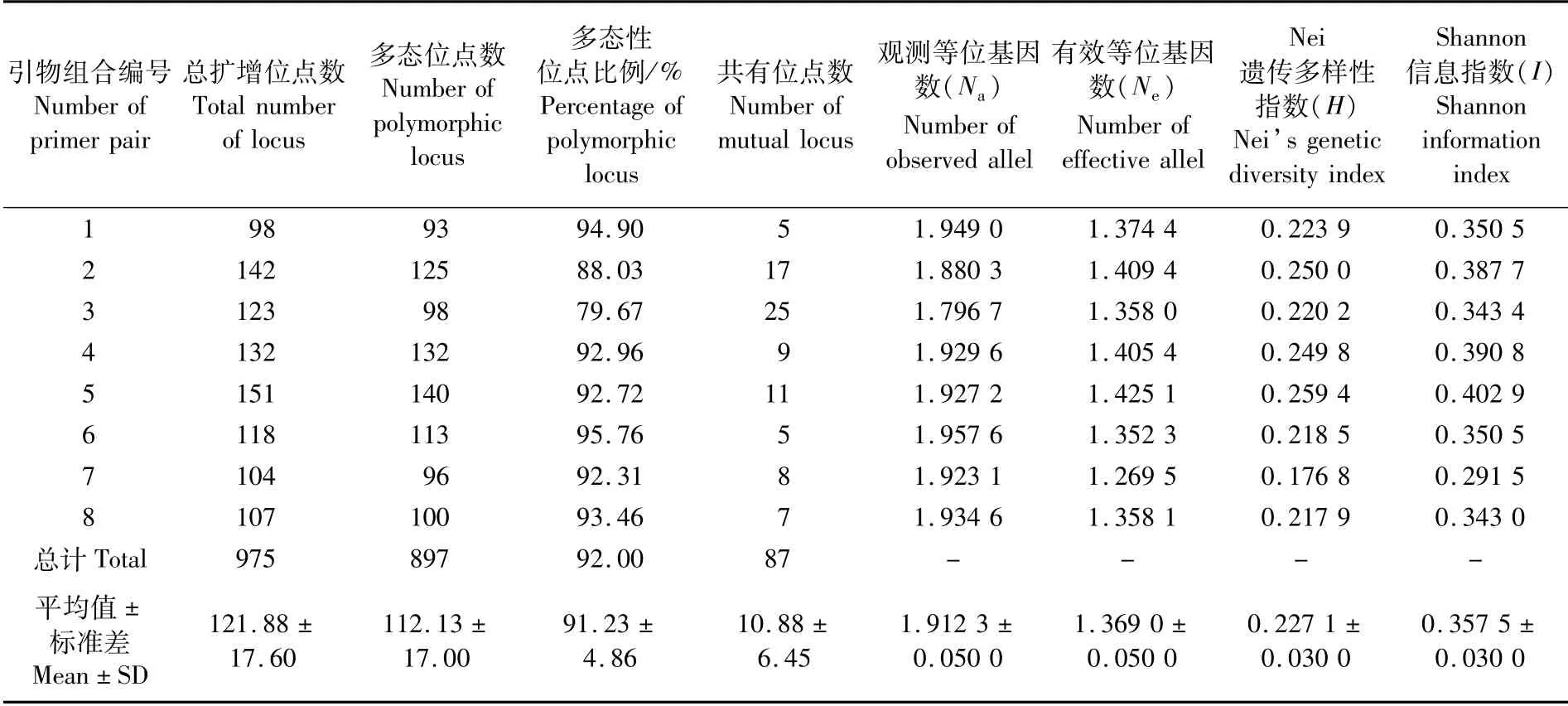
表3 光唇裂腹鱼遗传多样性参数Tab.3 Genetic diversity of Schizothorax lissolabiatus population from the Beipan River
根据8对扩增引物组合扩增出的975个位点,将30尾光唇裂腹鱼扩增位点的显性基因型频率以10%为单位划分为10个区间:1%~9%、10%~19%、20%~29%、30%~39%、40%~49%、50%~59%、60%~69%、70%~79%、80%~89%、90%~99%和0、1两个关键点(图2)。在关键点0扩增位点数出现峰值,在60%~69%区间扩增位点数呈现最低值。

图2 扩增位点数在不同显性基因频率区间内的分布Fig.2 Number of AFLP amplified locus in different intervals of occurency frequency of locus in Schizothorax lissolabiatus population from the Beipan River
2.3 遗传距离和系统树
北盘江光唇裂腹鱼种群30尾个体间遗传距离为0.214 7~0.410 9,平均为0.304 0。
基于个体间遗传距离对北盘江30尾光唇裂腹鱼进行聚类分析,结果显示采用UPGMA法和NJ法构建的系统树具有相似的拓扑结构,即30尾光唇裂腹鱼均聚为两支(图3,图4)。在UPGMA系统树中,一支由25个个体组成,另一支由5个个体组成;在NJ系统树中,一支代表24个个体,另一支代表6个个体。

图3 30尾光唇裂腹鱼UPGMA系统树Fig.3 UPGMA phylogenetic tree of 30 individuals of Schizothorax lissolabiatus from the Beipan River
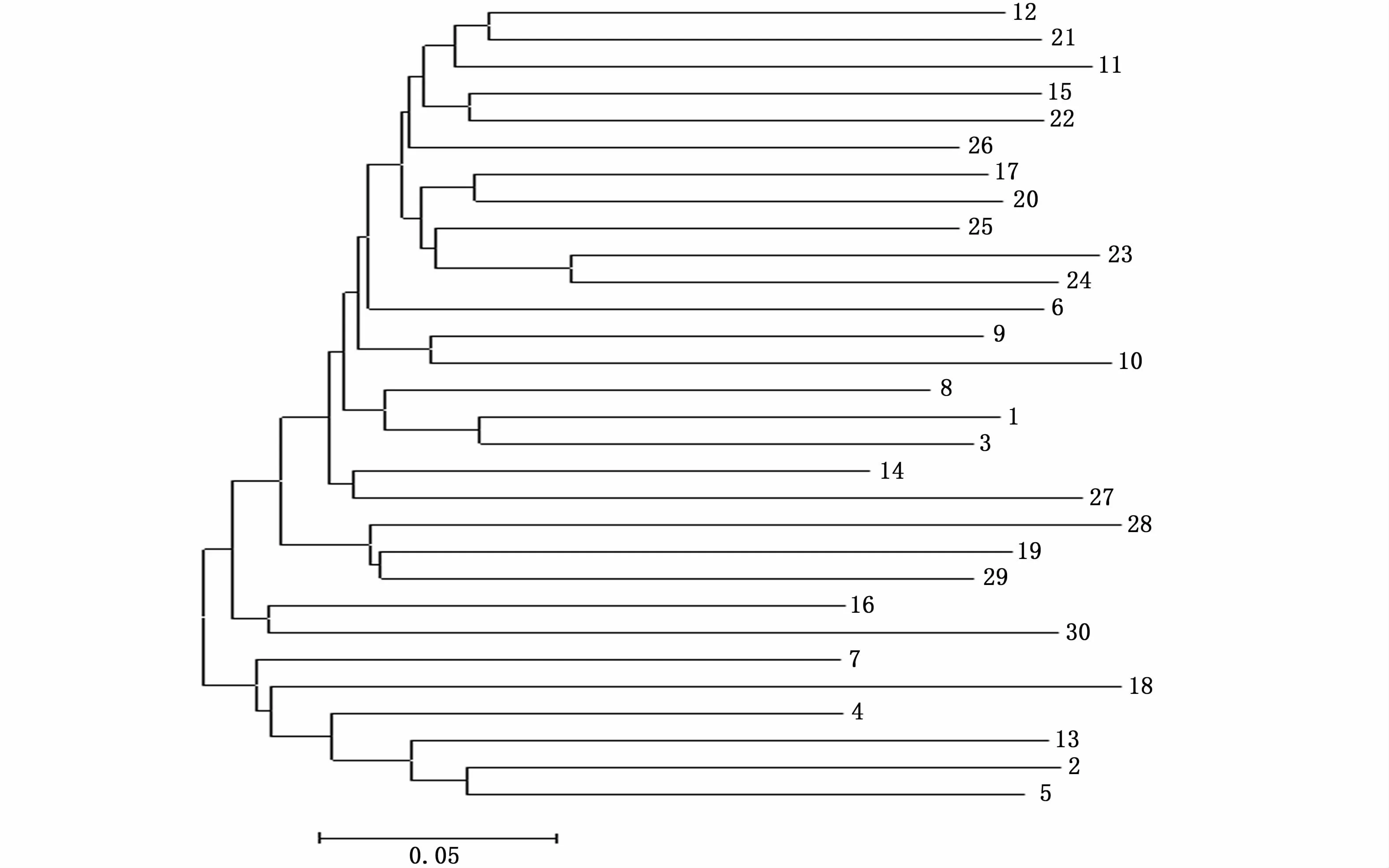
图4 30尾光唇裂腹鱼NJ系统树Fig.4 NJ phylogenetic tree of 30 individuals of Schizothorax lissolabiatus from the Beipan River

表2 8个AFLP引物组合的扩增结果Tab.2 AFLP amplification results by 8 primer pairs in Schizothorax lissolabiatus population from the Beipan River
3 讨论
3.1 北盘江光唇裂腹鱼种群的遗传多样性与保护
一个群体(或物种)遗传多样性越高或遗传变异越丰富,表明其对环境变化的适应能力越强,越容易扩展其分布范围和开拓新的环境[5]。群体遗传多样性每损失10%,就会对其繁殖能力、存活率、生长等重要生理性状产生极大的负面影响[12]。因此,对遗传多样性的研究,不仅可以了解物种的进化历史,也可以为分析物种的进化潜力和预测物种发展方向提供重要依据。
多态位点比例(P)和Nei遗传多样性指数(H)是评价物种遗传多样性的重要指标[13]。本研究中,北盘江光唇裂腹鱼种群的多态位点比例(P)、观测等位基因数(Na)、Nei遗传多样性指数(H)、Shannon信息指数(I)分别为91.23%、1.912 3、0.227 1和0.357 5(表3),高于同属的齐口裂腹鱼(Schizothorax prenanti)大渡河种群、雅砻江种群、青衣江雅安多营种群和宝兴种群(P=67.95%~77.94%)[14]、昆 明 裂 腹 鱼(Schizothorax grahami)六冲河群体(P=60.63%)[15],略低于澜沧江中上游光唇裂腹鱼4个地理群体(Na=2,H=0.286 8~0.324 8,I=0.426 9~0.481 1)[16],而与已被证实遗传多样性较高的乌江四川裂腹鱼(Schizothorax kolzovi)群体(P=92.99%,H=0.212 2)[17]处于同一水平。可见,北盘江光唇裂腹鱼种群具有较为丰富的遗传多样性,种质资源较佳。
然而,韩虎峰和代应贵[3]基于线粒体DNA控制区序列分析表明,珠江水系光唇裂腹鱼种群遗传多样性较贫乏,本文研究的结果与之相反。类似地,基于RAPD[18]、线粒体控制区[19]、Cyt b基因[20]及AFLP[21]等不同分子标记,对大弹涂鱼(Boleophthalmus pectinirostris)群体遗传多样性的研究也得出了相反的结论,推测这可能是因为不同标记的DNA分子大小、进化速率及多态位点信息等存在差异的结果。本研究中,珠江水系北盘江光唇裂腹鱼种群采用AFLP检测显示了较高的遗传多样性,而韩虎峰和代应贵[3]采用线粒体DNA控制区测序方法检测却表明该群体遗传多样性贫乏。其原因可能是由于本研究检测的是种群全基因组DNA的变异水平,而韩虎峰和代应贵[3]则仅仅检测了该种群线粒体DNA控制区部分序列(481 bp)的变异结果,显然本文的研究结果更为可信。
有效等位基因数(Ne)是衡量群体遗传变异程度的指标。有效等位基因数(Ne)与观测等位基因数(Na)之间的差异,能说明某等位基因在群体内分布的均匀程度。Ne与Na之间的差异越小,表明该等位基因在群体内分布的均匀度越高[7]。本研究中,8个引物组合Ne的范围为1.269 5~1.425 1、平均为1.369 0,Na的范围1.796 7~1.957 6、平均为1.912 3(表3),显示两者之间差异较大,表明该群体中等位基因分布的均匀度较低,可能存在着等位基因丢失的现象,也表明北盘江光唇裂腹鱼群体个体间基因交流较弱。Shannon信息指数(I)是反映群体离散程度的一个重要指标,Shannon信息指数的变化范围通常在1.5~3.5[7]。本研究中,北盘江光唇裂腹鱼群体Shannon信息指数(I)的平均值为0.357 5(表3),显著低于正常值范围,说明光唇裂腹鱼北盘江种群遗传变异离散程度较低。
光唇裂腹鱼为偏r-型物种[22],这类鱼类种群结构简单,世代交替快,更新能力强;但易受环境影响,资源稳定性较差[23]。多年来,北盘江干流梯级水电开发建坝[24],使河流变成了静水或缓流,阻断了鱼类的洄游通道,从而对适应于流水洄游生活的光唇裂腹鱼产生了不利影响。韩虎峰和代应贵[3]研究表明,光唇裂腹鱼在北盘江干流因支流革香河洗煤污水汇入而仅见于上游可渡河,种群数量减少。可见,水质污染是破坏北盘江光唇裂腹鱼资源的重要因素。同时,北盘江沿岸渔民的酷渔滥捕也严重影响了该河流的野生鱼类资源,导致其鱼类种群个体明显小型化[25],进而对河流中光唇裂腹鱼种群造成了威胁。
综上所述,尽管北盘江光唇裂腹鱼种群目前仍具有较丰富的遗传多样性,但因其自身的生态类型以及梯级电站建坝、水域污染和过度捕捞等因素的影响,该种鱼类在北盘江分布区已缩小,资源受到了威胁,遗传变异离散程度较低,存在着等位基因丢失的现象。因此,需要开展北盘江水域污染治理、加强渔政执法实行禁渔、减少北盘江流域水电站的修建及设置水电站过鱼通道、加大对北盘江光唇裂腹鱼资源监测的力度、实施北盘江光唇裂腹鱼人工增殖放流、设立北盘江光唇裂腹鱼自然保护区等措施,以保护和恢复北盘江光唇裂腹鱼野生资源和种群遗传多样性。
3.2 北盘江光唇裂腹鱼种群系统树分析
韩虎峰和代应贵[3]构建了珠江水系光唇裂腹鱼可渡河种群mtDNA单倍型NJ分子系统树,其所属的40尾光唇裂腹鱼被分为两支,推测该群体可能源于2个母系或两个繁殖群体。王绪祯等[26]研究表明,青藏高原特有的多倍体化裂腹鱼类包含了两个独立起源的类群。本研究中,光唇裂腹鱼为六倍体[26],基于个体间遗传距离构建的北盘江光唇裂腹鱼种群UPGMA和NJ系统树具有相似的拓扑结构,其所属的30尾光唇裂腹鱼聚为两支,显示北盘江光唇裂腹鱼种群可能源于具有不同亲缘关系的两个亚群体。
3.3 本研究中AFLP标记结果的的可靠性
AFLP标记多态性条带比例高,实验结果较稳定,不受基因组来源和复杂程度的影响,适用于检测亲缘关系较近的生物材料之间的遗传差异[27]。杨彦平等[28]采用AFLP技术对长江靖江段鳗鲡(Anguilla japonica)种群遗传多样性进行了分析,结果表明其多态位点比例高达95.06%。叶宁等[29]对凡纳滨对虾(Litopenaeus vannamei)7个群体遗传多样性的AFLP分析中,7个引物组合扩增出的多态位点比例为85.71%。基于AFLP方法,刘良国等[10]对洞庭湖水系五强溪水库光泽黄颡鱼(Pelteobagrus nitidus)遗传多样性的分析显示,其多态位点比例为87.7%。本研究中,用8个扩增引物组合共计检测出北盘江光唇裂腹鱼种群多态位点数为897,多态位点比例为92.00%(表3)。可见,AFLP技术多态性位点检出率高,可获得较丰富的遗传变异信息,是一种高效的分子标记方法,可以用于北盘江光唇裂腹鱼种群遗传多样性及遗传结构的研究。
据TAIJIMA[30]的DNA序列抽样分析理论,在DNA水平上估计种群变异时,当样本量≥10,抽样样本的方差便不会太大。大多数学者认为,为了避免因人为取样对遗传多样性研究结果造成影响,实验的样本量应在30以上,由此获得的遗传多样性结果才具有较高的可信度[31]。秦艳杰等[32]通过逐步增加AFLP引物组合数量进行了中间球海胆(Strongylocentrotus intermedius)群体遗传多样性的检测,认为至少要用3个引物组合才能对其群体进行可信的遗传学评价。沈德周等[6]研究了AFLP引物组合数量对马缨杜鹃(Rhododendron delavayi)遗传多样性的影响,表明基于5个以上引物组合即可获得较为准确的遗传多样性研究结果。本研究中,供检测的北盘江光唇裂腹鱼种群样本量为30,引物组合数为8,可实现对该种群体遗传多样性较准确的评价。因此,本研究中获得的北盘江光唇裂腹鱼种群遗传多样性研究结果可信度较高。
猜你喜欢
医学理论与实践(2021年13期)2021-07-12
疯狂英语·爱英语(2020年12期)2020-12-29
农村百事通(2019年2期)2019-03-26
电脑知识与技术(2017年10期)2017-06-05
电子技术与软件工程(2017年8期)2017-05-10
科技视界(2016年27期)2017-03-14
科学种养(2017年1期)2017-02-16
农家科技(2014年11期)2015-03-11
中国美容医学(2004年4期)2004-10-25
中国美容医学(2004年3期)2004-09-17
