
武夷山自然保护区不同海拔甜槠天然林土壤有机碳变化特征及影响因素
2021-08-07 02:11王世君吴承祯林勇明
生态学报 2021年13期
赵 青,刘 爽,陈 凯,王世君,吴承祯,李 键,*,林勇明
1 福建农林大学林学院,福州 350002 2 福建省高校森林生态系统过程与经营重点实验室,福州 350002 3 福建省生态产业绿色技术重点实验室,南平 354300 4 武夷学院生态与资源工程学院,南平 354300
在全球范围内大约有1550 Pg碳是以有机质的形式储存于土壤中,土壤有机碳(soil organic carbon,SOC)储量占到全球碳库总储量的2/3以上,是大气碳库(750 Pg)的2倍,生物碳库(500—600 Pg)的3倍[1-2]。森林土壤碳库作为陆地碳库的重要组成部分,占全球土壤有机碳库的70%以上[2],其微小变化即会显著引起大气圈中CO2浓度波动,从而影响全球气候变化。因此深入研究森林SOC,揭示其分布特征与影响因素对于准确评估全球碳储量及碳循环具有重要的意义。
目前,国内外学者已经对SOC的累积、组成及差异性开展了大量的研究工作,杨玉海等[3]对伊犁河谷9种不同植被类型土壤有机碳含量研究表明,土壤有机碳含量与植被类型显著相关;孟苗婧等[4]对不同海拔梯度下黄山松针阔混交林土壤有机碳含量研究得出,高海拔处土壤温度过低会对SOC的分解产生影响;丛高等[5]通过分析不同海拔SOC与黏粒矿物组成之间的关系,得出不同林型土壤有机碳含量及其组分之间差异显著;Post 等[6]表明SOC含量受植被类型的影响,其大小由植物残体量及微生物分解量决定;Zinn 等[7]通过研究不同海拔巴西森林SOC含量,指出海拔是影响热带森林SOC含量的主要因素;但这些研究集中于在不同区域、不同植被类型间对SOC含量进行研究,由于森林植被覆盖类型、水热条件、土壤理化性质等条件的综合影响,以及森林土壤时空异质性的复杂性,对不同森林植被类型在不同区域上SOC含量的估算存在极大的不确定性[8]。特别是海拔作为一个综合性的影响因子,可以通过对大尺度区域环境下水热条件的重新分配对SOC含量产生影响[9- 10],因此研究同一地区单一植被类型在不同海拔高度SOC含量分布特征及影响因素对于准确估算区域SOC储量具有重要的现实意义。
武夷山自然保护区作为世界双遗产保留地,拥有同纬度位置保存较为完好的中亚热带常绿阔叶林,是研究中亚热带森林SOC的理想区域[11]。甜槠林不仅是武夷山自然保护区常绿阔叶林群落的优势种之一[12],也是中亚热带的典型植被类型之一,在中亚热带森林生态系统碳平衡中占有重要的地位[13]。近年来,相关学者围绕武夷山自然保护区在不同海拔土壤磷素分布[14]、土壤微生物氮分布[15]、表层土壤轻组有机质分布[16],SOC矿化特征[17]等方面进行了大量研究,然而对于单一森林树种在不同海拔SOC分布特征及影响机制方面研究较少,因此,本研究以甜槠林集中分布的5个海拔高度土壤作为研究对象,分析甜槠林SOC沿不同海拔高度及不同深度垂直剖面土层分布规律,揭示SOC含量分布特征及影响机制,旨在为武夷山自然保护区土壤碳储量研究提供基础数据,同时也为中亚热带森林碳储量及碳循环研究提供一定的参考。
1 材料与方法
1.1 研究区概况
研究区位于武夷山自然保护区内(27°32′36″—27°55′15″N;117°24′12″—118°02′50″E),平均海拔1200 m,本区拥有世界同纬度现存面积最大、保存最为完整的中亚热带常绿阔叶林生态系统,气候类型属典型的亚热带季风气候,年平均气温为12.7—18.1℃,年平均降水量在2000 mm左右,年平均相对湿度为82%—85%,年平均雾日在120 d以上,无霜期为252—272 d。区内地势高低悬殊,起伏较剧烈,土壤垂直分异显著,沿海拔从下至上分别为红壤(海拔700 m以下)、黄红壤(海拔700—1100 m)、黄壤(1100—1900 m)、山地草甸(1900 m以上);植被垂直带谱完整,沿海拔由低到高分别为常绿阔叶林(200—1000 m)、针阔混交林(1000—1350 m)、针叶林(1350—1750 m)、亚高山矮林(1750—1900 m)与高山草甸(1900 m以上)[18- 19]。
1.2 样地设置及样方调查
将武夷山自然保护区甜槠林集中分布的5个不同海拔作为实验样地,分别为生态定位站(540 m)、一里坪(700 m)、三港(850 m)、七里村(1022 m)、玄峰岭(1200 m)。每个海拔梯度内设置3块规格为20 m×20 m的固定样地,共选取15块样地,利用手持GPS确定每块样地位置,同时记录每块样地坡度、坡向等地形因子、平均胸径、树高等林分因子、腐殖质层、淋溶层厚度等土壤因子(表1),采样时间为2017年6月。

表1 样地情况调查
林下植被状况:甜槠天然林海拔540 m处植被群落乔木层主要树种为木荷(Schimasuperba)、赤楠(Syzygiumbuifolium)、少叶黄芪(Engelhardtiafengelii)等,灌木层树种主要有宜兴苦槠(Pleioblastusyixingensis)、马银花(Rhododendronovatum)、冬青(Ilexchinensis)、芒萁(Dicranoprerisdichotoma);海拔700 m处甜槠天然林乔木层树种主要有赤楠(Syzygiumbuifolium)、深山含笑(Micheliamaudiae)、灌木层树种有香槟杜鹃(R.champinicane)、马银花(Rhododendronovatum)、赤楠(Syzygiumbuifolium)、草本层树种主要有鸡血藤(SpatholobussuberectusDunn)、里白(Diplopterygiumglaucum)。海拔850 m处植被群落乔木层主要树种为木荷(Schimasuperba)、青冈栎(Cyclobalanopsisglauca)、马尾松(Massonpine)、刺毛杜鹃(RhododendronsimsiiPlanch)等,灌木层树种主要有马银花(Rhododendronovatum)、山杜英(Elaeocarpussylvestris)、香槟杜鹃(R.champinicane),草本层树种主要有淡竹叶(Lophatherumgracile)、里白(Diplopterygiumglaucum);海拔1022 m甜槠天然林群落乔木层主要有木荷(Schimasuperba)、山杜英(Elaeocarpussylvestris)、拉氏栲(CastanopsislamonteiiHance),灌木树种主要有大叶赤楠(Photiniasp)、少叶黄芪(Engelhardtiafengelii)等,草本树种里白(Diplopterygiumglaucum)、淡竹叶(Lophatherumgracile);海拔1200 m甜槠林植被群落乔木层树种有木荷(Schimasuperba)、石栎(Lithocarpusglabra)、细枝柃(Euryaloquaiana)、鹿角杜鹃(Rhododendronlatoucheae)、以及一些针叶树种如马尾松(Massonpine)、杉木(Cunninghamialanceolata)等混生其中,灌木层主要有肿节少穗竹(Oligostachyumoedogonatum)、冬青(Ilexchinensis),草本层较疏松,主要以蕨类为主要如里白(Diplopterygiumglaucum),狗脊(Woodwardiajaponica)等。
1.3 土壤样品采集及测定
每块样地内采用S型方法布设3个土壤采样点,共45个采样点;土壤采样采用分层采样法,取样时将土壤表层枯枝落叶清除,用直径为5 cm的土钻采集0—10、10—20、20—30 cm 3个土壤土层,每个样地重复取样3次。将相同海拔同一样地同层土层土壤样品混合均匀装入密封袋带回实验室,待土样自然风干后,捡出石砾、根系、枯枝落叶等杂物,过2 mm筛用于指标测定。每个样地挖掘1个0—30 cm土壤剖面,用环刀采集土样带回实验室用于测定土壤容重。根系生物量测定采用土钻法及全根挖掘法进行取样,每块样地随机选取3棵林龄相近的成年甜槠(20 a左右),在距离树干1—1.5 m处用直径为5 cm的根钻钻取土芯,共钻取3个,用于采集0—10、10—20、20—30 cm土层垂直土芯,将同块样地采集土芯分层混合装袋,每个海拔3次重复,将其带回实验室用20目筛网反复清洗后,捡出细根(根径≤2 mm),风干后称鲜重,然后烘干至恒重。
土壤理化性质测定均采用常规方法测定[20]。SOC采用硫酸—重铬酸钾外加热法进行测定,在温度170—180℃条件下的油浴锅加热保持5 min,然后用5 ml的浓硫酸氧化土壤中的有机碳,再用硫酸亚铁来对添加了邻啡罗琳指示剂的溶液进行滴定,根据所消耗的硫酸亚铁来计算SOC的含量,将本研究已测定的SOC含量乘以换算系数1.724即求得土壤有机质含量。土壤全氮采用硫酸—半微量凯氏定氮法(ATN—300全自动定氮仪);土壤全磷采用碱熔—钼锑抗比色法;土壤pH值采用水土比为2.5∶1纯水浸提pH计法(Starter 3100);土壤温度、含水量采用TDT(Time Domain Trans-missometry; Acclima, Inc. Meridian, ID, USA)探头进行测定,探头水平埋设在距离地表5、15、25 cm处,分别代表土壤0—10、10—20、20—30 cm土层温度及土壤含水量值,同时用烘干法对测出的含水量结果进行校准。
1.4 数据分析
采用SPSS17.0进行单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)分析比较不同土层SOC在不同海拔间的差异(α=0.05),采用Pearson相关系数法确定各因子间相关关系,采用主成分分析法对相关环境因子进行分析;采用线性回归分析(一元及多元)分析SOC与土壤理化性质、气候因子、植物因子间的相关性。
2 结果与分析
2.1 同一土层土壤有机碳含量沿海拔梯度分布特征
武夷山天然甜槠林不同海拔高度SOC在同一土层深度上的垂直分布规律如图1所示,5个海拔高度中,SOC含量均随土层的增加而降低,且降幅随海拔的升高逐渐增加。其中,SOC含量在海拔1200 m中0—10 cm土壤土层达到最大,为72.59 g/kg;在海拔540 m中20—30 cm土层土壤降至最低,为15.58 g/kg。除海拔1022 m外,所有海拔3个土层SOC含量差异均显著(P<0.05)。0—10 cm土层除海拔1022 m SOC含量与850 m、1200 m SOC含量差异不显著外,其余海拔SOC含量差异均显著;10—20 cm,除海拔540 m SOC含量差异显著外,其余海拔SOC含量差异均不显著;20—30 cm,除海拔1200 m SOC含量与850 m、1022 m差异不显著外,其余海拔SOC含量均有显著差异(P<0.05)。0—10 cm土壤表层SOC含量占整个剖面(0—30 cm)SOC含量的48.2%—52.7%,且各海拔0—10 cm土层SOC含量显著高于10—20、20—30 cm土层(P<0.05)。

图1 同一土层深度土壤有机碳含量沿海拔梯度分布特征Fig.1 Distribution characteristics of soil organic carbon content in the same soil depth along the uplift gradient不同大写字母表示同一海拔不同土层间差异显著(P<0.05),不同小写字母表示同一土层不同海拔间差异显著(P<0.05)
2.2 环境因子相关指标及与土壤有机碳含量相关关系
从表2可知,同一海拔不同土层土壤全氮差异均显著,0—10 cm土层土壤全磷在海拔850 m处最高,海拔1200 m处最低,除海拔850 m外,其余海拔土壤全磷差异均不显著;10—20、20—30 cm土层在海拔850 m处最高,海拔540 m处最低,且20—30 cm土层海拔850m土壤全磷与其余海拔差异均显著。0—10土层土壤pH值在海拔540 m处最高且与其余海拔显著不同;0—10、10—20、20—30 cm土层土壤含水率随海拔升高逐渐增大,且海拔1200 m土壤含水率与其余海拔差异显著;除海拔850 m、1022 m外不同土层土壤温度差异均显著;同一海拔不同土层土壤容重、细根生物量、土壤有机质、土壤粉粒、粘粒含量差异均显著。
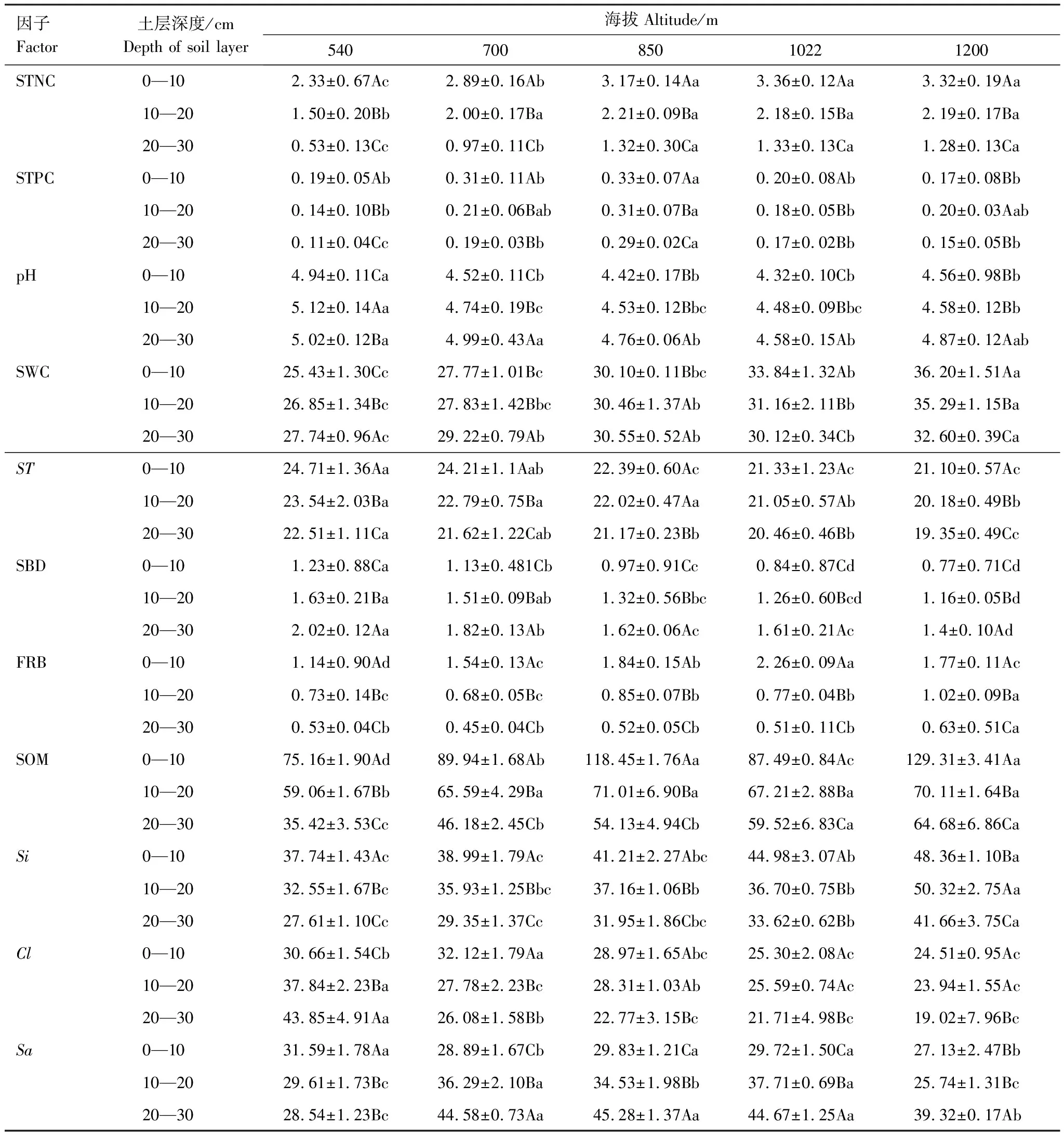
表2 不同海拔不同土层环境因子相关指标
通过Pearson相关性分析结果显示(表3):不同土层SOC含量与海拔、土壤全氮、土壤含水量、土壤温度、土壤粉粒均呈均呈显著(P<0.05)或极显著相关关系(P<0.01);土壤细根生物量、土壤有机质含量仅与0—10、10—20 cm土层SOC含量呈显著(P<0.05)或极显著(P<0.01)正相关关系;不同土层SOC含量与土壤全磷不相关,土壤pH值、土壤砂粒含量仅与20—30 cm土壤底层SOC含量显著(P<0.05)或极显著相关(P<0.01)。这表明武夷山天然甜槠林0—30 cm范围内不同深度土层SOC含量的改变依赖于海拔高度、土壤容重及土壤机械组成变化,以及由海拔升高引起的土壤温度,土壤含水量变化。
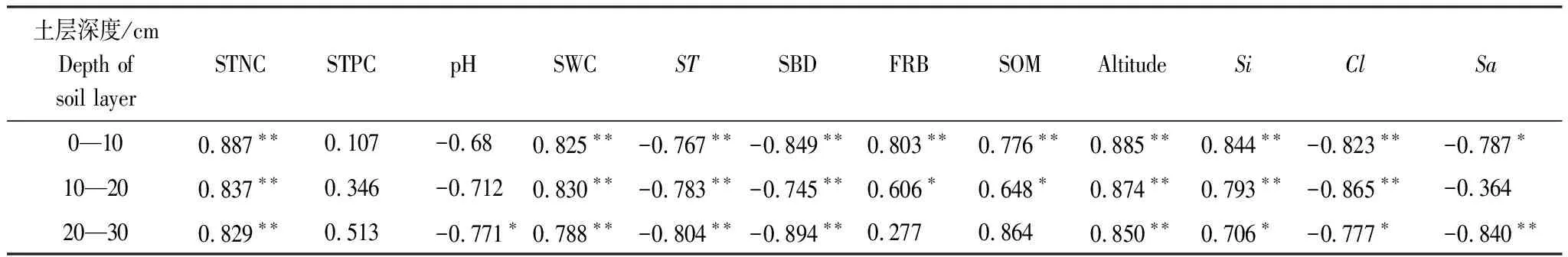
表3 不同土层土壤有机碳与环境因子相关性
2.3 影响土壤有机碳含量的因子分析
由于细根生物量是植物自身指数,因此归为植物因子,土壤pH值、土壤全氮含量、土壤容重、土壤有机质、土壤粉粒、粘粒、砂粒是土壤理化指标,归为土壤因子,土壤含水量、土壤温度归为气候因子。本研究通过主成分分析提取影响武夷山天然甜槠林土壤各层SOC的主要因子(表4)。结果显示,0—10、20—30 cm土层主成分中大于1的特征值成分有2个,累计解释变量分别为83.277%、82.291%,10—20 cm土层主成分中大于1的特征值成分有3个,累计解释变量为87.752%,能反映不同海拔垂直分异土层SOC的绝大部分信息。0—10 cm土层第一主成分SOC受土壤全氮、土壤含水量、土壤温度、土壤粉粒、粘粒、容重、生物量、海拔等地形、土壤、气候、植被因子的影响,累计解释变量达68.809%;10—20 cm土层第1主成分SOC与海拔、土壤含水量、土壤温度、粘粒、容重相关性大,累计解释变量达45.635%,第二主成分中土壤粉粒与SOC相关性大,累计解释变量达67.086%;20—30 cm土层第1主成分SOC受海拔,土壤温度、土壤含水量、土壤有机质、土壤粉粒等土壤因子的影响,累计解释变量达48.719%,第二主成分中土壤砂粒与SOC相关性最大,累积解释20—30 cm土壤SOC变异达82.291%。
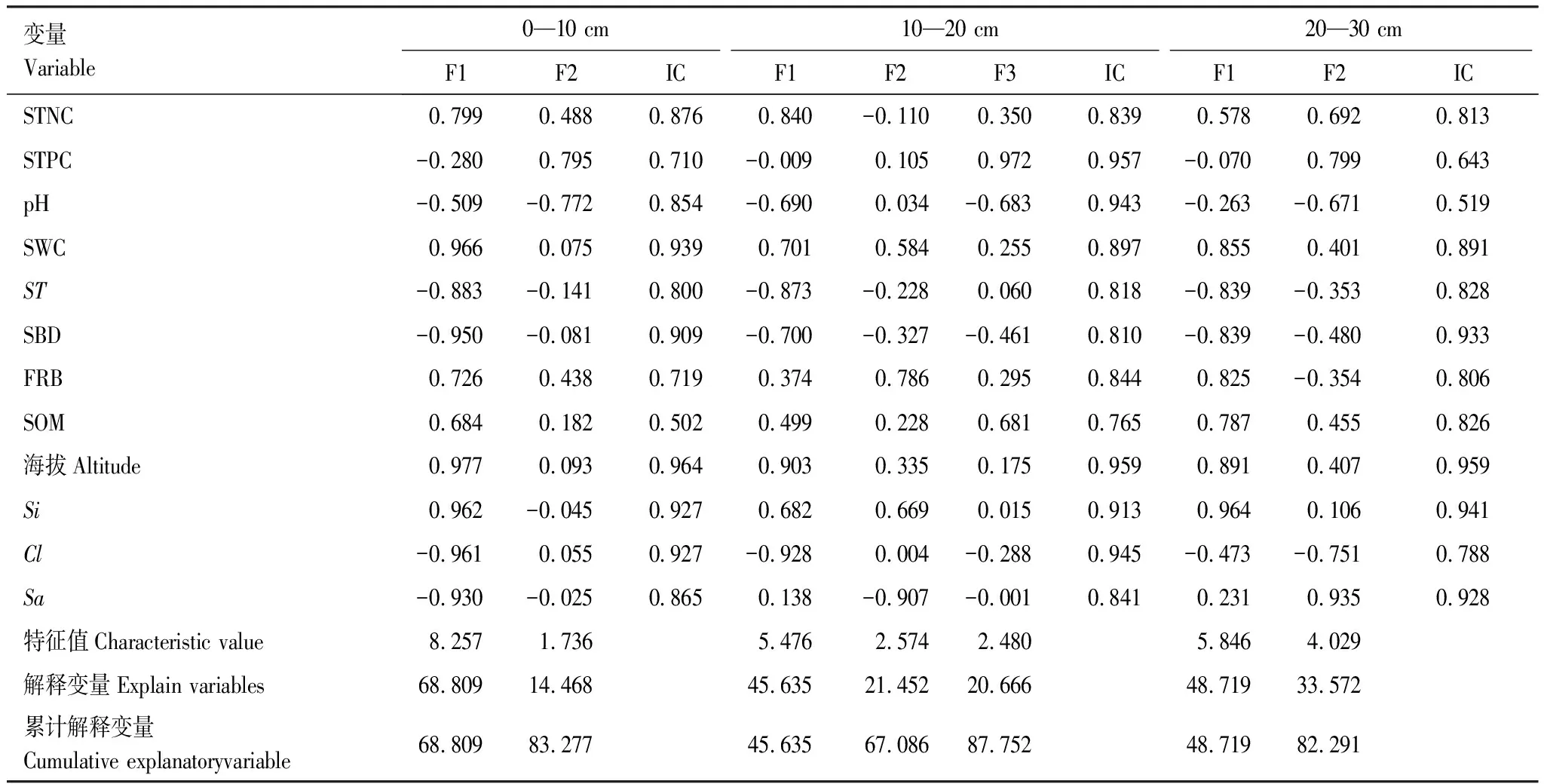
表4 不同土层土壤有机碳与环境因子分析
2.4 土壤有机碳含量回归模型创建
主成分分析法的应用虽然可以得到各影响因子间总体分类关系,同时众多因子间多重共线性能进一步被消除,但不能得到各影响因子间的定量关系,只分析单个影响因子不能完全弄清各因子对SOC含量的影响机制,因此对主成分分析得出的各土壤层次主要影响因子进一步做回归分析,建立相应模型量化各影响因子对SOC的影响程度。
2.4.1不同土层土壤有机碳含量一元线性回归模型
基于单变量一元回归分析,可得出不同土层主要影响因子与SOC含量的一元线性回归方程(表5)。三个垂直土层SOC含量均与海拔、土壤含水量、全氮、容重呈极显著相关关系(P<0.01),除此之外,0—10 cm土层与细根生物量、土壤粉粒、粘粒亦呈极显著相关关系(P<0.01),能解释土壤有机碳含量的64.5%—78.6%的变异,其中全氮与土壤有机碳含量拟合R2最大(78.6%);10—20 cm土层除细根生物量与SOC呈显著相关外(P<0.05),亦与土壤粉粒、粘粒达到极显著相关(P<0.01),能解释土壤有机碳含量的55.6%—74.8%的变异,其中土壤粘粒与土壤有机碳含量拟合R2最大(74.8%);20—30 cm土层还与土壤砂粒、粘粒达极显著相关关系(P<0.01),能解释SOC含量的60.5%—78.9%的变异。

表5 不同土层土壤有机碳含量一元线性回归方程
2.4.2不同土层土壤有机碳含量多元线性回归模型
基于多元回归分析,可得出不同土层主要影响因子与SOC含量的多元线性回归方程(表6)。0—10 cm土层中地形因子(海拔)、土壤性质(全氮、容重)与SOC含量拟合R2为0.832,地形因子(海拔)、土壤性质(全氮)、植被因子(细根生物量)与SOC含量拟合R2为0.843,高于单因素回归拟合R2;地形因子(海拔)、土壤性质(全氮、容重、土壤粉粒、粘粒)与SOC含量拟合R2为0.866,高于单因素回归拟合及相关多元线性回归组合R2,可见海拔、全氮、容重与土壤机械组成结合能更好的解释0—10 cm SOC含量的变异;10—20 cm土层中,海拔、土壤全氮、水分、温度、土壤粉粒、粘粒与SOC含量拟合度最高(0.940),能更好的解释不同海拔10—20 cm SOC含量的变异;20—30 cm土层中海拔、土壤砂粒、容重、pH与SOC含量拟合度最高(0.981)。可见,多因子对SOC的解释明显高于单因子,SOC含量变化是多种因子综合作用的结果。

表6 不同土层土壤有机碳含量多元线性回归方程
3 结论与讨论
3.1 甜槠天然林土壤有机碳剖面分布特征及空间分布规律
武夷山天然甜槠林同一海拔SOC含量垂直分异表现出随土层增加而降低,且土壤表层(0—10 cm)SOC含量明显高于底层(20—30 cm),呈现明显的“表聚性”特征,这与王会利等[21]对青冈林土壤剖面SOC垂直分布及蔡文良[22]对热带山地雨林SOC分布研究结果一致。一方面这可能是因为土壤表层积累了大量枯枝落叶及动植物残体,同时甜槠天然林根系主要分布于土壤表层,而表层SOC主要来源于根系分泌物,枯枝落叶及根系的腐殖质等,加之表层适宜的土壤温度、水分等环境因子,良好的通气和养分状况使得好气微生物活动旺盛[23- 24],更有利于地表凋落物分解、腐殖化过程,使得土壤容重降低,有机质含量增加,土壤碳含量不断累积;另一方面随着土层深度增加,植物凋落物减少,植被根系密度降低,土壤质地变的愈发紧实(表2),表层土壤有机质向下迁移受到限制[25],使得SOC含量随土层深度增加而降低,但在不同海拔间降幅存在较大差异,这种垂直分布差异可能与不同海拔甜槠天然林土壤有机质及细根生物量分布差异有关,甜槠林细根主要分布于0—20 cm土壤表层,而细根高度富集于土壤表层有利于高效吸收来自凋落物分解释放的养分,同时可以截留林冠淋溶的养分,影响进入土壤中有机物的含量及在剖面中的分布[26],进而对不同土壤剖面SOC含量造成影响。
3.2 甜槠天然林土壤有机碳剖面分布差异及影响因素
SOC累积量与土壤表层有机质输入及周转速度相关,尤其是对于山地森林生态系统,海拔因素是影响SOC累积的主导因子之一[27]。本研究表明,武夷山自然保护区甜槠天然林SOC分布特征与海拔因素呈极显著正相关(P<0.01),表现为0—30 cm SOC含量随海拔高度增加而增大,这与张亚茹等[28]表明鼎湖山常绿阔叶林SOC具有强烈的空间异质性,且随着海拔升高而增加研究结果一致,但与Sheikh[29]对亚热带松果林、温带阔叶林SOC随海拔升高而降低;曹瑞等[30]对高山峡谷区SOC组分研究表明,SOC、水溶性碳随海拔增加呈先增加后减少结果不一致,这可能与植被类型、地形地貌条件不同导致SOC在不同海拔地区呈现不同的变化规律有关。海拔不仅作为一个地形因子,还作为一个综合性的环境因子,并非直接对土壤中的SOC含量产生影响,而是通过与温度、湿度、光照等多种环境因子对其进行响应[31],一方面通过制约植被生产力从而影响进入土壤中的有机质含量,另一方面土壤温度、水分等环境因子随海拔条件的改变影响了土壤微生物活动,进而对SOC的分解及转化产生影响[32]。本研究表明,不同海拔垂直土层SOC含量与土壤温度呈极显著负相关(P<0.01),与土壤含水量呈极显著正相关(P<0.01),细根生物量及植被凋落物随土壤含水量的增加而增大,成为土壤表层SOC的主要输入来源[33]。随着海拔升高土壤含水量及凋落物含量显著增加(P<0.05),增强了微生物活性[34],而土壤温度随着海拔梯度上升开始降低,使得有机质分解转化速率减慢,增加了SOC的累积。本研究中SOC含量与土壤含水量之间的相关性高于土壤温度与SOC含量间的相关性,这可能是因为在同一气候区内采样时间气温相差不大,处于雨季降水量较多、垂直差异较大有关。
3.3 不同海拔梯度下甜槠天然林土壤有机碳影响因素
相关研究表明,植被群落结构特征与土壤理化性质共同决定了SOC的储量,而土壤理化性质对SOC的影响要高于植被群落结构特征[35],这与本研究结果相似。Pearson相关分析表明,虽然SOC含量与土壤理化性质随垂直土层增加存在一定的差异性,但不同海拔垂直剖面SOC含量与土壤全氮,有机质均呈极显著(P<0.01)或显著正相关(P<0.05),这与相关学者研究一致[27,30,36]。土壤容重作为土壤物理性质的一个重要指标,反映了土壤的通气、透水以及松紧程度,能显著影响土壤的持水能力和入渗性能[37],本研究指出,不同海拔垂直剖面SOC含量与土壤容重呈极显著负相关,即土壤土层深度加大,甜槠天然林根系减少,土壤通气性变差,土壤容重也随之增大,这与相关学者研究一致[5,8,17- 18]。这可能是因为甜槠天然林土壤表层植被凋落物丰富,凋落物的分解与积累增加了土壤有机质,使得土壤回还养分增多土壤容重变小;随着土层深度增加,孔隙度变小,土壤水分和有机物质向下输入受到制约[38],同时土壤透气性变差导致土壤微生物因氧气供给受限而活性减弱,土壤质地紧实容重变大。不同土层SOC仅在20—30 cm土层与土壤pH值呈显著负相关,这与潘忠松[39]研究结果一致,即马尾松人工林SOC含量随pH值降低呈增加趋势。主要是因为土壤pH值能直接影响矿质元素溶解度,影响土壤中各元素的有效性及迁移转化,随着土壤pH值的降低,土壤中微生物活性减弱,同时参与SOC循环中的酶活性也受到一定程度的限制[40],SOC周转及矿化作用下降,使得SOC不断累积。而本研究中土壤pH值仅与底层土壤(20—30 cm)显著相关很可能是因为武夷山自然保护区甜槠林表层环境适宜,微生物活性强,土壤全氮及有机质的极显著相关关系可能掩盖了土壤pH值对SOC累积的影响。本研究表明,土壤细根生物量与0—20 cm土层SOC呈极显著或显著相关,甜槠天然林在海拔1022 m处细根生物量最大,海拔540 m处细根生物量最小,这种变化趋势与相关学者研究结果不同,刘建军[41]在对天然油松锐齿栎林研究发现,随着海拔升高油松锐齿栎林细根生物量显著减少;Davis[42]对阿巴拉契亚山5种典型植被类型细根生物量研究也发现了同样规律,这种差异性可能与植被所处气候带、林分类型以及取样方式不同有关。本研究中海拔1022 m处细根生物量最大可能与高海拔甜槠天然林枯枝落叶层较厚,受人为干扰少,而海拔540 m处甜槠天然林枯枝落叶层较薄且受人为因素影响较大有关。
土壤机械组成比例能通过其化学保护机制降低土壤SOC的分解速率,同时促进土壤SOC的固持[43]。本研究中不同海拔土层土壤粒径组成中粉粒含量随海拔升高而升高,与土壤SOC分布特征一致,土壤粘粒含量随海拔升高而降低,与土壤SOC分布变化相反,土壤砂粒含量在0—10 cm土壤表层随海拔升高逐渐降低,在10—20、20—30 cm土层中随海拔增加呈先增加后减小趋势。其中0—10 cm土层土壤粒径组成中,粉粒含量与SOC的影响最强,可作为衡量土壤表层SOC含量的重要因素,这与胡宗达等[44]对川滇高山栎林土壤SOC与不同土层土壤机械组成关系的研究结果一致。表3中0—10 cm土层土壤SOC含量与土壤粘粒及砂粒含量呈显著或极显著负相关,与土壤粉粒呈极显著或显著正相关,20—30 cm土层土壤砂粒含量与SOC呈极显著负相关,印证了以上分析,也与前人研究一致[45- 46]。这可能是因为土壤SOC,特别是其活性组分会与土壤机械组成中的矿质土粒机械混合,且其中一部分土壤SOC会溶解于低分子化合物,部分氨基酸等土壤溶液中,其余一部分则通过生命体方式存在于土壤中[47]。然而大部分土壤SOC是与土壤中的无机成分结合形成有机—无机复合体,一般认为,土壤SOC易与土壤颗粒中的粉粒结合,对土壤水分的有效性及植被生长产生影响,从而影响土壤SOC分布[48]。一方面,由于粉粒颗粒较细,具有较大的表面积,能暴露更多的正电荷位与土壤中带负电荷的腐殖质结合[49];另一方面,土壤颗粒越细,其通透性越差,一旦与土壤SOC结合,即很难被微生物所分解[46],而砂粒等土壤颗粒较大的土壤组分与粉粒、粘粒刚好相反,其颗粒大,带正电荷位少,不易与土壤SOC结合,且通透性强,微生物易分解,因此,土壤SOC与砂粒含量呈显著负相关,与粉粒含量呈显著正相关。
SOC具有明显的空间变异性,即使在土壤质地相同区域内,空间位置的不同也会造成土壤有机碳含量存在显著差异[50]。本研究揭示了武夷山自然保护区甜槠天然林SOC随海拔梯度变化特征及影响SOC含量变化的环境因子,不同环境因子组合虽然能解释不同土层SOC含量的大部分变异,但仍有部分变异未被解释,因此未来研究应进一步考虑其他因素如坡向、土壤微生物、林龄等因子及其交互作用对SOC累积的影响。
猜你喜欢
橡塑技术与装备(2022年7期)2022-12-13
西北林学院学报(2022年5期)2022-10-04
农业工程学报(2022年8期)2022-08-08
北京大学学报(自然科学版)(2022年3期)2022-06-17
专用汽车(2021年11期)2021-11-18
棉花学报(2020年3期)2020-08-08
中国水土保持科学(2017年3期)2017-07-05
浙江工业大学学报(2016年3期)2016-06-29
浙江大学学报(工学版)(2016年2期)2016-06-05
中国农业文摘-农业工程(2016年5期)2016-04-12
