
布鲁氏菌病患者尿液差异蛋白质组学分析
2021-08-06 08:40杨文涛张建中吴翠萍倪秀莹
中国人兽共患病学报 2021年7期
杨文涛,张建中,吴翠萍,王 磊,赵 利,倪秀莹,蓝 峰,姜 海,肖 迪
布鲁氏菌病(Brucellosis,简称布病)是由布鲁氏菌侵入机体所引发的传染—变态反应性传染病,是在全世界范围内广泛传播的一种人兽共患病,世界卫生组织将布病列为7种最容易被忽视的疾病之一[1]。经文献报道,布病可通过分娩、哺乳、性交、输血及骨髓移植等途径在人与人之间传播[2]。人患布病后,可发生多器官受损,在急性期因误诊或治疗不当可转为慢性布病,严重影响生存质量,在我国乃至全球范围内疾病负担巨大。人畜间布病疫情在全世界170多个国家和地区均有报道,每年新发病例约有50万例,布病流行国家的患病率超过每10/10万人[3]。近年多有未直接接触家畜的学生、儿童及离退休人员发病,表明布病已经由主要的职业接触感染向非职业的食源性感染转变,通过食用生鲜羊奶导致的食源性暴发公共卫生事件时有发生[4]。
目前,布病诊断依赖血清免疫学检查结合病原学细菌分离实验。常用的血清学检查方法有虎红平板凝集试验、试管凝集试验等。随着国家对布病的重视,快速检测技术已取得长足进步,间接酶联免疫吸附试验、胶体金免疫层析法等检测方法逐步开始使用,但现有技术仍存在一定缺陷,对临床诊断存在较大的漏诊和误诊率[5]。实时荧光PCR是目前病原体检测应用广泛和稳定的分子诊断技术,在新突发性传染病相关病原体感染的实验室确证中具有不可替代的地位和作用[6]。但在病原体分子检测领域存在突出问题:核心探针技术无自主知识产品,需要发展具有自主知识产权且性能更好的核心探针技术;布鲁氏菌荧光PCR检测产品尚无标准化试剂,且无标准化操作规程。针对国家高致病性传染病应急处置的重大需求,创建新突发传染病诊断方法、诊断试剂应急研发和产业化技术平台,形成快速响应体系迫在眉睫[7]。
继二维凝胶电泳-质谱可视化差异蛋白发现与确认技术之后,基于高效液相色谱-质谱串联系统(UPLC-MS/MS)的定量差异蛋白质组学技术已发展成熟,可实现快速、高通量地获得样本中差异表达的蛋白,在发现诊断标志物、确定疫苗相关抗原、发现药物作用靶点、阐述致病机理方面具有广泛的应用[8-9]。尿液是一种易获得、患者接受度高的检测分析物,已成为多种疾病或生理特征监测与检测的标本[10]。迄今国内外尚无采用定量蛋白质组学技术在人尿液标本中发现及确定布病诊断标示分子的报道。本研究将采用高效液相色谱-质谱串联系统进行布病相关尿液定量蛋白质组学分析,通过专业软件检索确认差异表达的蛋白,并通过生物信息学分析软件进行蛋白信号通路分析和生物标志物筛选分析,从而获得蛋白表达“全景图”,发现潜在的诊断布病的标志蛋白,挖掘关键蛋白的相关生物信息,在蛋白水平上为今后布病诊断、监测及治疗提供分子线索。
1 材料与方法
1.1 样本信息 布病患者样本来自于已通过血清学检查确诊,试管凝集试验(SAT)滴度为1∶100及以上或血培养中分离到布鲁氏菌加之临床表现或有流行病学史明确确诊的人群。非布病患者样本来源为非布鲁氏菌感染、无临床表现、无流行病学史的人群。两组各10例样本,按统一要求采集晨起中段尿液50 mL,采集后冻存-80 ℃。所有尿液样本采集均经过伦理委员会审核。
1.2 样本前处理
1.2.1 尿液蛋白提取和定量 将样本从-80 ℃冰箱中取出后,12 000 g 4 ℃离心10 min,取上清3.5 mL,加入8倍体积-20 ℃预冷含20%三氯乙酸(TCA)的丙酮溶液,混匀后将样本置于-20 ℃过夜;12 000 g 4 ℃离心30 min后,弃上清,向沉淀中加入8倍体积-20 ℃预冷丙酮,振荡混匀后置于-20 ℃ 4 h;12 000 g 4 ℃离心30 min后, 弃上清,室温下静止2~3 min使沉淀干燥。用8 mol/L尿素溶解蛋白沉淀,震荡混匀后BCA试剂盒(Thermo Fisher Scientific, USA)测定蛋白浓度。
1.2.2 溶液内酶解 向提取的蛋白溶液中,加入200 mmol/L 三(2-羧乙基)膦(TCEP),混匀后室温放置1 h进行还原,再加入375 mmol/L碘乙酰胺(IAA),混匀后避光放置30 min进行烷基化。向处理好的样本中加入6倍体积-20 ℃预冷丙酮,混匀后-20 ℃过夜放置,8 000 g 10 min 后弃上清,室温下静置2~3 min 使沉淀干燥,沉淀中加入100 μL 100 mmol/L NH4HCO3混匀,按胰酶和蛋白1∶50质量比加入胰酶,混匀后37 ℃水浴过夜。
1.2.3 肽段除盐和定量 向样本中加入10%三氟乙酸(TFA)后,用C18 除盐柱(Thermo Fisher Scientific, USA)对肽段进行纯化,氮气吹干样本。对所有样本用肽段定量试剂盒(Thermo Fisher Scientific, USA)进行定量。
1.3 高效液相色谱-质谱串联分析 肽段定量后,每个样本取1 μg肽段使用EASY-nLCTM1000色谱仪(Thermo Fisher Scientific, USA)进行分离。流动相A液为0.1%甲酸水溶液,流动相B液为0.1%甲酸乙腈水溶液(乙腈80%)。色谱分析柱: PepMap 100, nanoViper C18,100 Å, 3 μm, 75 μm×25 cm,预柱: PepMap 100, nanoViper C18,100 Å, 5 μm, 100 μm×2 cm。流速为300 nL/min。液相梯度设置:0~45 min, B液4%~15%;45~73 min,B液15%~25%;73~100 min, B液25%~40%; 100~105 min, B液40%~95%;105~120 min, B液95%。使用Q-ExactiveTMPlus质谱仪(Thermo Fisher Scientific, USA)Full MS/ddMS2(TopN)串联模式采集数据,具体参数为:全扫描一级谱图范围300~1 800 m/z,分辨率70 000,最大积分时间100 ms,离子目标值3×106个。而后选择10个最强母离子进行二级数据采集,高能碰撞解离,分辨率17 500,最大积分时间60 ms,离子目标值2×105个,设置串联质谱扫描的动态排除时间为30 s。应用 Xcalibur 软件(version 4.1, Thermo Fisher Scientific, USA)记录谱图。
1.4 数据分析
1.4.1 数据质量控制 为避免人为造成蛋白表达呈现差异,本研究采取以下方法确保数据的可靠性:为保证样本处理一致性,同组尿液样本同批次进行样本前处理;为保证定量准确性,对所有酶切后的尿液蛋白肽段样本进行再定量,每个样本均取1 μg肽段进行高效液相色谱-质谱串联分析;为保证色谱质谱检测的稳定性,检测样本前高效液相色谱-质谱串联系统进行校正,并在实验过程中用保留时间校正液对样本出峰时间和杂质峰进行监测。
1.4.2 蛋白质组数据分析 将10例布病患者与10例非布病患者的尿液样本质谱数据对比,采用Proteome Discoverer(version 2.4, Thermo Fisher Scientific, USA)软件检索Proteomes-Homo sapiens(Human)数据库(20 600条蛋白序列,下载日期2020年8月26日)。选择两个酶解漏切位点;半胱氨酸脲甲基化(carbamidomethyl)为固定修饰,蛋氨酸氧化(oxidation)为可变修饰;肽质量偏差为10 ppm,离子片段偏差为0.02 Da,用反转数据库去除假阳性。
1.4.3 生物信息学分析 将归一化处理后蛋白定量数据中蛋白水平FDR阈值设置为0.01,剔除任意缺失的数据,对归一化后的数据进行log2转换,筛选组内变异系数小于0.3的数据进行单因素方差分析,并以非布病患者为对照得到蛋白表达的差异倍数(Fold Change)。将| Fold Change(log2)|>1.5,且BH法校正P<0.05定义为差异表达蛋白,同时对仅在一组中检测到并在组内无任意缺失的蛋白定义为特异表达蛋白或缺失表达蛋白。使用“悟空”平台(https://www.omicsolution.org/wkomics/main/)进行假设检验和GO富集分析。应用商业化生物信息分析软件IPA(Ingenuity pathway analysis)对获得的差异表达蛋白进行典型通路分析(Canonical pathways)和生物标志物筛查分析(Biomarker filter)。
2 结 果
2.1 高分辨液质联用数据采集 使用UPLC-MS/MS采集尿液样本的质谱数据,总离子流图(Total Ion Chromatography,TIC)见图1。本研究中质谱数据采集灵敏度高,TIC图显示所有样本峰强度均大于1E10,同组的TIC图总离子流强度变化趋势基本一致,同组样本的出峰时间偏差较小。

图1 布病患者(A)和非布病患者(B)尿液样本的总离子流图
2.2 蛋白鉴定与数据质量评估 应用Proteome Discoverer软件进行原始数据检索,采用label-free定量分析方法将布病患者与非布病患者尿液样本原始数据进行比对,得到定性和定量蛋白的数据。蛋白数据散点图显示了在每个采集时间点,谱图与肽段的匹配(Peptide spectrum matching, PSM)的强度分布的集中范围在105~1010(图2A),共采集到164 616条PSMs。样本丰度图显示组内样本蛋白相应肽段的丰度有较好的一致性,保证了定量分析时组间的可比性和定量的准确度(图2 B)。UPLC-MS/MS分析定性鉴定了1 817个蛋白(unique peptide≥1),其中定量鉴定到1 732个蛋白,蛋白定量数据经质量控制后将其中873个蛋白用于后续蛋白质组学分析。经差异倍数及显著性水平筛选后,最终得到185个与布病相关的尿液差异表达蛋白,其中13个蛋白上调、172个蛋白下调,使用火山图来确定两组之间差异表达蛋白变化的意义。
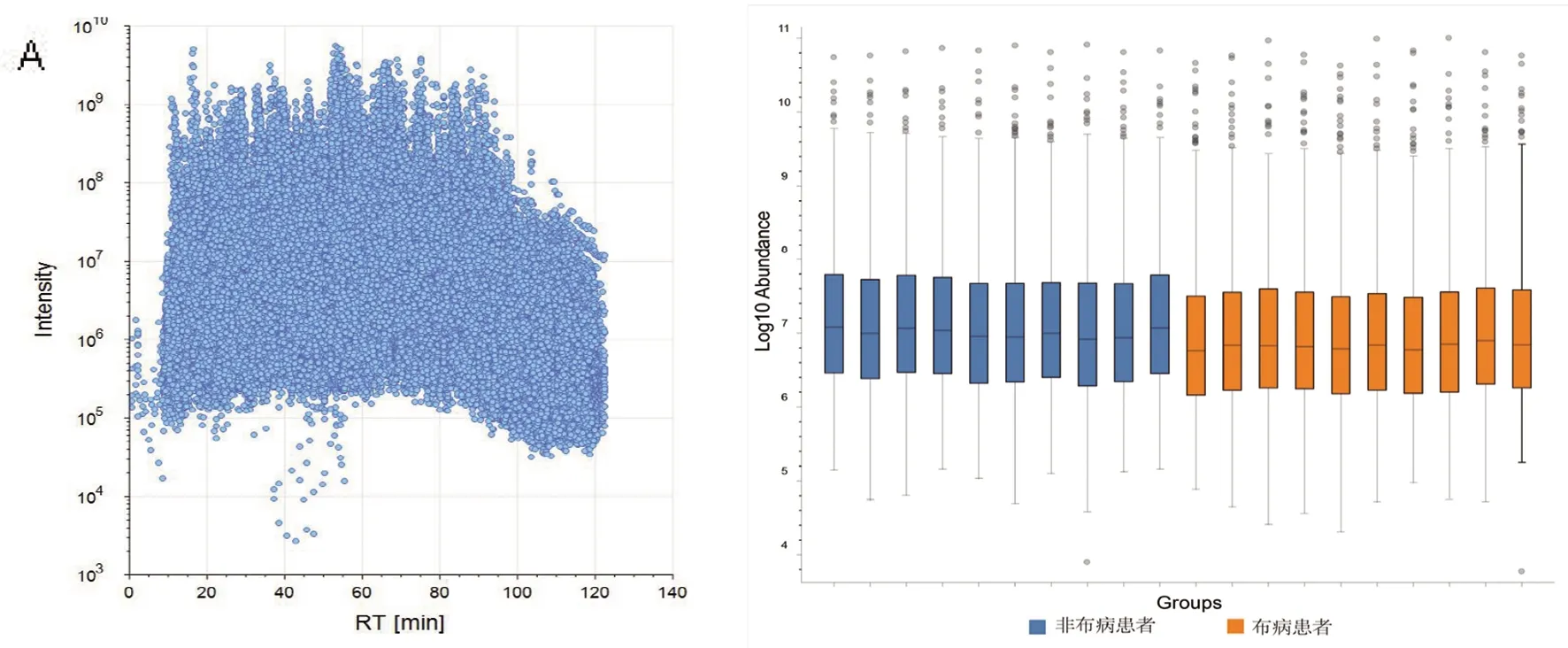
图2 散点图(A)和样本丰度图(B)
和程度(图3)。

图3 差异表达蛋白火山图
2.3 GO富集分析 使用GO二级术语的基因本体论分析,根据蛋白质在3个主要类别(细胞成分、分子功能和生物过程)中的参与情况对蛋白质进行分类。185个差异表达蛋白主要为细胞外外泌体、质膜和细胞外空间的成分;分子功能分类表明,大多数差异表达蛋白的细胞功能与钙离子结合、肝素结合及蛋白质均与二聚活性有关;差异表达蛋白主要参与的生物过程包括细胞黏附、血管生成及细胞外基质组织等(图4)。布病患者中缺失表达的胰岛素样生长因子2(IGF2)功能分类分析表明,该蛋白与骨骼系统发育、骨化和血小板脱颗粒等生物进程有关。
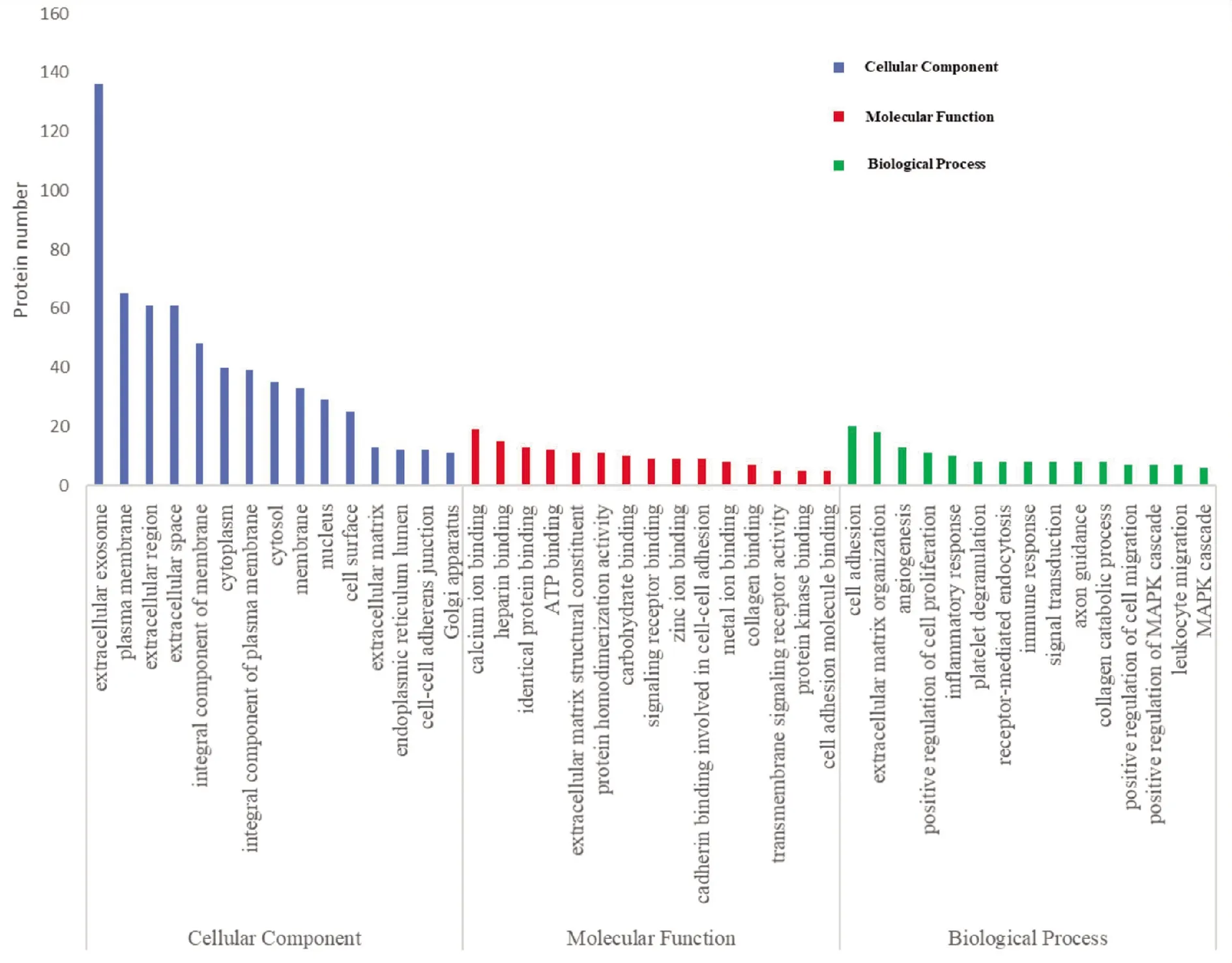
图4 差异表达蛋白GO富集分析(前15条)
2.4 生物信息学数据挖掘 商业化软件IPA是分析和挖掘多组学数据的生物信息分析平台。IPA整合了已有公共数据库、已知的生物学知识,经几百名科学家人工阅读生物医学文献、总结科研成果,构建了包含700多万条生物信息的IPA支持数据库,并不断更新。IPA广泛应用在多组学研究、靶标的发现及验证、机制研究、生物网络分析、生物标记物研究等研究领域,具有分析全面、数据可靠的特点。本研究应用IPA软件对差异表达蛋白进行典型通路分析和生物标志物筛查分析,以确保数据结果的严谨性和准确性。本研究在尿液样本中共发现185个与布病相关的差异表达蛋白,其中有1个蛋白在IPA数据库中未匹配成功,因此对其余的184个差异表达蛋白进行了生物信息学分析。这些差异表达蛋白参与疾病和功能排名前五的是:细胞妥协、炎症反应、细胞运动、癌症、组织损伤和异常(图5A)。这些差异表达蛋白参与了肝纤维化/肝星状细胞活化、糖酵解、GP6 信号通路、LXR/RXR 激活和动脉硬化信号等45个经典通路(图5B)。其中预测对糖酵解、GP6信号通路、LXR/RXR激活、葡萄糖生成及IL-15生产等共12个通路有抑制作用,预测对LPS/IL-1介导的RXR功能抑制通路有激活作用。本研究中发现IGF2蛋白在布病患者中表达缺失,其参与的调控网络主要调节胚胎发育、器官形态和生物发育3种疾病和功能,重叠上下游调节因子后显示有16个下游调节因子和6个上游调节因子(图6)。对184个差异表达蛋白进行生物标志物筛选分析,发现有30个蛋白已作为多种疾病的效应、诊断、预测指标,其余的154个差异表达蛋白目前尚未作为其他疾病的生物标志物;此外有17个差异表达蛋白是本研究在人的尿液中首次发现并呈现布病表达差异的蛋白(表1)。

表1 人尿液中首次检测到的17个差异表达蛋白
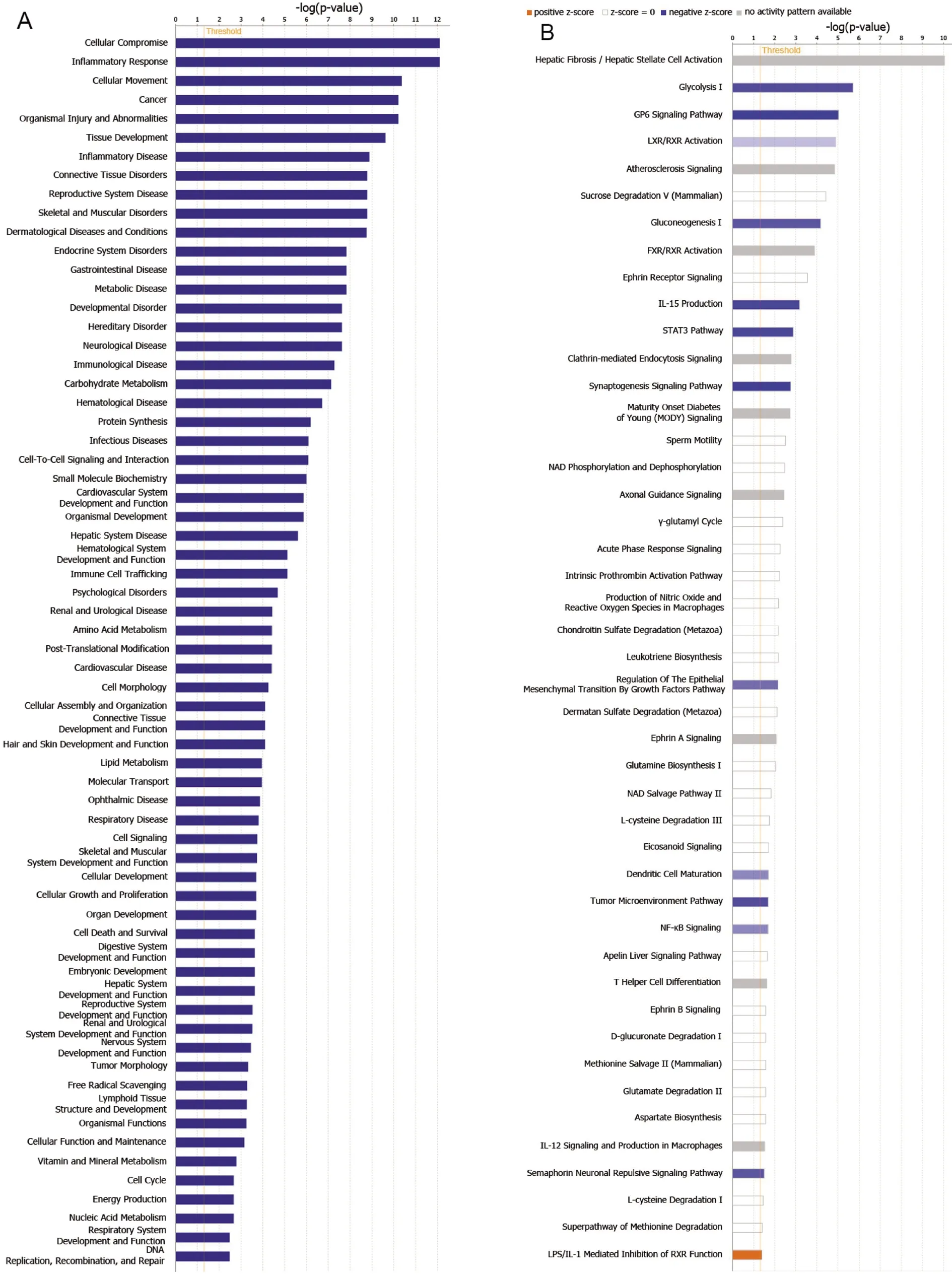
A:疾病和功能分布;B:信号通路分布
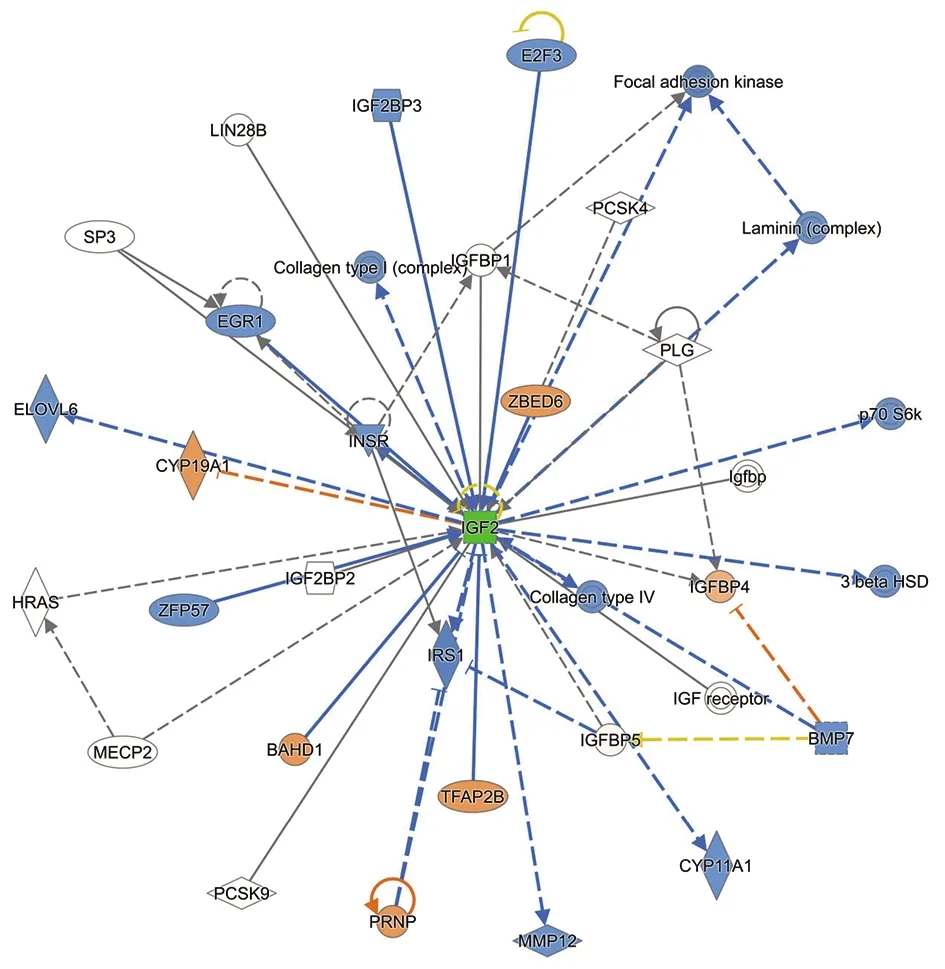
图6 IGF2的调控网络以及上下游调节因子(橙色代表上游调节因子,蓝色代表下游调节因子)
3 讨 论
基于高效液相色谱-质谱联用的蛋白质组学已经成为研究生物标志物的热门工具,疾病生物标志物的发现有助于疾病的诊断、预后效果的评估以及药物靶点的发现,对临床应用和疾病监测有着重要的意义。本研究采用UPLC-MS/MS对布病患者尿液进行定量蛋白质组学的检测,采集的质谱数据灵敏度高、样本定量一致性较好,为发现布病感染相关的生物标志分子提供了准确和可比的蛋白数据。本研究是生物标志分子的发现阶段,获得了大量的差异表达蛋白数据,通过数据深度挖掘在蛋白表达差异倍数、生物标志物应用和参与的经典信号通路分析方面发现更具研究意义的蛋白,后续将进一步扩大样本量对所发现的差异蛋白进行验证,获得切实可用的生物标志分子,为开发基于尿液的布鲁氏菌感染诊断试剂提供靶标抗原。
本研究共定性鉴定到1 817个蛋白,定量鉴定到1 732个蛋白。相较于非布病患者,布病患者尿液中有13个蛋白表达上调,172个蛋白表达下调。研究表明,机体感染病原体后,病原体与宿主免疫系统之间的持续相互作用会启动复杂的免疫反应,以通过多种抗寄生物效应子功能阻止病原体感染和生长,包括抑制侵袭和细胞粘附,抗体依赖性细胞毒性和细胞抑制作用[11]。与布病感染相关差异蛋白的GO富集分析中, 其中一些功能条目支持了目前研究认为的可能与布病感染相关的分子生物学过程, 包括细胞黏附、炎症反应和免疫反应等。此外,基于IPA数据库,本研究发现17个蛋白在已有的人尿液相关研究中未见报道,这些差异表达蛋白可以对现有的公共数据库进行补充。
布病患者尿液差异表达蛋白中IGF2、CD300分子样家族成员f(CD300LF)、血清类粘蛋白1和2(ORM1,ORM2)、富含亮氨酸α-2-糖蛋白1(LRG1)、SH3结构域谷氨酸富集样蛋白3(SH3BGRL3)、半胱氨酸丰富跨膜BMP调节剂1(CRIM1)和谷氨酸氨连接酶(GLUL)是更具潜力的诊断布病的候选标示蛋白。
IGF2有调节细胞生长、分化和新陈代谢的作用[12]。细菌感染后,细菌细胞壁脂多糖刺激单核巨噬细胞释放TNF-α、白细胞介素等,均可促进生长激素分泌,间接促成IGF2及其受体合成及分泌[13]。本研究发现IGF2在健康人尿液中正常表达[14],并已经用于威尔姆斯肿瘤、大肠癌、卵巢癌的诊断标志物,但在布病患者尿液样本中缺失表达,提示IGF2在尿液中缺失表达可能是布鲁氏菌感染后的特异性表现,可作为潜在的布病诊断生物标志物。人患布病的主要临床表现是发热、疲劳、关节痛和肌肉疼痛[15],常见并发症有骨关节炎[16-17]和血液系统并发症[18]等。IGF2在体内参与了骨骼系统的发展和血小板脱粒等生物进程,提示尿液中IGF2的缺失表达可能对布病临床并发症有一定的预测作用。CD300家族的成员在白细胞膜上表达,该家族由7个基因组成,命名为CD300a到CD300g,可通过成对的触发和抑制功能调节免疫反应[19]。树突状细胞(DC)是一类异质性白细胞,被认为是关键的抗原提呈细胞和免疫应答的重要启动者[20]。CD300分子现已被证明其在体外和体内都能调节DC的功能[21]。本研究中CD300LF表达上调8.57倍,但在已有的人尿液相关研究中CD300LF蛋白未见报道,提示CD300LF上调可能是诊断布病感染的重要尿液标志蛋白。
潜在生物标记物表达水平的改变经常是非特异性的,并且可能在多种传染性疾病中都会发生改变,这可能是炎症介导的急性期反应信号转导的作用[22]。ORM1和ORM2是与急性炎症相关的蛋白,ORM蛋白主要的肝细胞中表达[23]。目前经过验证ORM1的上调与乳腺癌有关[24],脑脊液中ORM1的上调与慢性疲劳综合症有关[25],血清中ORM1的上调与COVID-19相关[26]。本研究中ORM1和ORM2表达上调,上调倍数分别为6.14和4.22,并参与了LXR/RXR激活、动脉粥样硬化信号、FXR/RXR激活和急性时相反应信号等信号通路。有研究表明RXR与FXR的异二聚体和RXR与LXR的异二聚体均可参与免疫反应[27-28],进一步的提示ORM1和ORM2可能是布病诊断及感染后调节体内的免疫反应的重要蛋白。LRG1是参与蛋白相互作用、细胞粘附、信号转导和细菌感染的急性炎症的蛋白[29]。LRG1在人血清中含量丰富,主要由肝细胞产生,并与肿瘤发生和血管生成有关[30]。已有研究显示LRG1的表达与亨廷顿病、帕金森病痴呆、肝癌、进行性核上性瘫痪疾病有关,但目前并未作为任何疾病的标志物。本研究中LRG1表达上调5.46倍,提示LRG1有潜力作为布病诊断候选标示蛋白。
有研究在溃疡性结肠炎患者尿液中检测到SH3BGRL3上调,SH3BGRL3在尿路上皮癌中的高表达与复发风险增加和生存率降低有关[31]。本研究中发现SH3BGRL3表达上调8.66倍。CRIM1是骨形态发生蛋白(BMPs)的拮抗因子,可抑制BMPs前蛋白转变为成熟的BMPs,并且可以抑制成熟的BMPs呈递到细胞表面的过程[32]。本研究中CRIM1表达下调10倍(差异倍数为0.10)。谷氨酰胺连接酶在氨解毒、酸碱平衡和细胞信号转导中起重要作用[33],并参与了血管生成过程中的病理性细胞迁移[34]。本研究中GLUL表达下调12.5倍(差异倍数为0.08)。因此,人尿液中蛋白SH3BGRL3表达上调、CRIM1表达下调和GLUL表达下调,可能对布病感染以及临床并发症的预测有指示作用。
布鲁氏菌最重要的特征是能够在吞噬细胞和非吞噬细胞内生存和繁殖。布鲁氏菌主要毒力因子包括脂多糖(LPS)、T4SS分泌系统和BvrR/BvrS系统,它们可以与宿主细胞表面相互作用,形成早、晚BCV(含空泡的布鲁氏菌),并在细菌繁殖时与内质网(ER)相互作用[35]。目前对布鲁氏菌致病机制研究表明布鲁氏菌含有不同的毒力因子,可降低抗原提呈细胞的吞噬能力与抗原提呈能力,延缓细胞成熟,抑制细胞凋亡,减少相关细胞因子的释放[36]。本研究中CD58等5个差异表达蛋白参与抑制树突状细胞成熟信号通路,表明这些差异表达蛋白可能延缓了细胞成熟,降低了机体对布病感染的免疫反应的调节。NF-κB转录因子是免疫反应、应激反应、细胞凋亡和分化的重要调节因子,细菌和病毒感染、炎性细胞因子和抗原受体的结合都能诱导NF-κB的激活[37]。有文献报道称布鲁氏菌强弱毒株侵染小鼠巨噬细胞后可以不同程度地激活NF-κB通路,而布鲁氏菌LPS对NF-κB信号通路的激活的影响不大[38]。但本研究中显示布病感染后EGF等5个差异表达蛋白参与抑制NF-κB信号通路,提示布病感染后导致机体内蛋白表达差异,从而影响对不同信号通路的调节,因此人感染布病后对NF-κB信号通路的影响有待进一步研究。
目前传染病的常规诊断主要依赖于对致病病原生物的检测以及临床症状的检查,但也可以通过宿主标志物的分析实现对病原体感染的敏感的和早期的诊断[39]。另外,临床中应用多个蛋白标志物往往比单个标志物在疾病机理,诊断和治疗等方面可以提供更全面的信息[40]。本研究基于非标记定量蛋白质组学发现了多个潜在的布病相关标志蛋白,为布病临床诊断、监测及治疗提供了技术支撑。
利益冲突:无
猜你喜欢
上海人大月刊(2022年4期)2022-04-14
中老年保健(2021年4期)2021-08-22
现代畜牧科技(2021年4期)2021-07-21
中国畜禽种业(2020年4期)2020-12-16
农民致富之友(2019年35期)2019-01-13
中国畜禽种业(2019年6期)2019-01-05
中国卫生(2016年1期)2016-11-12
中国民族医药杂志(2016年9期)2016-05-09
中国民族医药杂志(2016年4期)2016-05-09
中国卫生标准管理(2015年17期)2016-01-20
