
热应激对齐兴肉兔母兔繁殖性能、血清生化指标及生殖激素含量的影响
2021-08-05 13:51邝良德李丛艳郭志强任永军梅秀丽谢晓红
西南农业学报 2021年6期
邝良德,李丛艳,郭志强,任永军,郑 洁,梅秀丽,杨 锐,谢晓红,雷 岷*
(1.四川省畜牧科学研究院,四川 成都 610066;2.动物遗传育种四川省重点实验室,四川 成都 610066)
【研究意义】现代集约化生产中,应激反应易于发生并对畜禽生产有很大负面影响,尤其是夏季热应激会带来一系列问题。四川等南方地区的夏季潮湿多雨,极端气温可高达40 ℃以上,而肉兔全身被毛,汗腺功能不发达,温度和湿度的变化会扰乱它们的体温调节系统[1]。热应激对家兔会产生一系列广泛的影响。【前人研究进展】Asemota等[2]研究认为,温度-湿度指数(THI)会影响母兔的受胎率和仔兔断奶后死亡率。Ondruska等[3]研究表明,温度的升高对新西兰兔的总采食量、日采食量、饲料转化率、总增重和日增重均有显著影响。Marco-Jiménez等[4]研究表明,妊娠期间母兔热应激会对产仔数、初生窝重、死胎率有负面影响,哺乳期的热应激会影响仔兔的生长性能。Marder等[5]研究表明,热应激会使家兔体温升高,血液碳酸氢盐浓度显著下降,渗透压增加,血清尿素、葡萄糖、肌酐、球蛋白含量和谷草转氨酶活力升高,造成肌肉损伤、高血糖和酸中毒,甚至死亡。国内也有很多相关报道。张小丽等[6]研究认为热应激与肉兔的直肠温度、体表温度、呼吸频率、生长育肥等有直接关系。文凤云等[7]研究发现热应激使獭兔的血清葡萄糖、甘油三酯、氯化物和钾离子浓度升高,血清总胆固醇、钙离子和无机磷含量则下降。徐辰义等[8]认为随着温度的升高,家兔采食量明显下降,体液免疫和细胞免疫功能降低,激素水平和血液理化指标受到影响。热应激可引起动物生理调节异常,不同的品种、个体耐热性能差别较大,会受到遗传因素的影响[11-12]。成年兔的最适温度为15~25 ℃,温度过高就会产生热应激[13]。妊娠母兔更加难以消除体内多余的热量,极易出现热应激,从而影响肉兔养殖的生产效益。为了减少夏季热应激对养兔业造成的损失,通过选择耐热性好的个体培育耐热新品种,具有非常重要的现实意义。【本研究切入点】本试验拟研究热应激对齐兴肉兔母兔繁殖性能、血清生化指标及生殖激素含量的影响。【拟解决的关键问题】此研究为选育耐热性好的新品种提供理论依据。
1 材料与方法
1.1 试验设计
本试验于2019年6-8月在四川省畜牧科学研究院肉兔科研基地进行。选择200只健康齐兴肉兔初产母兔,随机分为适温组和热应激组2个处理,每个处理100只兔,分别饲养在2个环控兔舍中。其中适温组采用空调降温,人工加湿,环境温度为(21±1)℃;热应激组采用电暖风加热,人工加湿,环境温度为(32±1)℃。试验前对兔舍进行彻底消毒,试验期间自由采食、饮水,除温度外,其他各环境因素保持一致,相对湿度80%左右。适应期为2周,正式期为母兔配种至第一胎仔兔断奶(28日龄断奶),所有试验母兔均在同一天进行配种,饲养管理条件一致(日粮主要营养水平为粗蛋白18.12%,粗纤维14.53%,消化能10.70 MJ/kg)。
1.2 环境THI指数测定
在全环控兔舍中部距地面15 cm处挂置Apresys 179-TH温湿度记录仪,设10 min为间隔记录试验期间温度和湿度,利用Marai等[14]提出的ITH公式测定环境ITH指数,ITH指数公式如下:
ITH=db-[(0.31-0.31RH)(db-14.4)]
式中:db、RH分别表示温度计数值(℃)、相对湿度(%)。
依据Marai 等[14]的方法确定家兔发生热应激的ITH阈值为27.8,ITH<27.8 时为轻微热应激或无热应激反应;ITH在27.8~28.9之间处于中等热应激;ITH在28.9~30.0之间处于严重热应激;ITH>30 时为特别严重热应激。
1.3 测定指标及方法
1.3.1 繁殖性能测定 分别测定适温组和高温组母兔情期受胎率、胎产仔数、胎产活仔数、初生窝重、21 d窝重、仔兔断奶重、断奶成活率等繁殖性能指标。
1.3.2 血清生化指标测定 分别于母兔配种当天(G0)、妊娠7 d(G7)、妊娠11 d(G11)、妊娠18 d(G18)、妊娠21 d(G21)、妊娠28 d(G28)、产后当天(L0)、产后7 d(L7)、产后11 d(L11)、产后18 d(L18)、产后21 d(L21)、产后28 d(L28)从适温组和热应激组分别随机选取6只母兔采集血液5 mL,分离血清,-70 ℃保存待测。血清谷草转氨酶(GOT)、谷丙转氨酶(GPT)活力采用分光光度法测定,血清皮质醇、三碘甲状腺原氨酸(T3)、免疫球蛋白G(IgG)、免疫球蛋白A(IgA)、孕酮(P4)、雌二醇(E2)、催乳素(PRL)含量采用双抗体夹心法ELISA测定。
1.4 数据统计分析
试验结果用平均值±标准误表示。利用SPSS 11.5软件进行单因素方差分析和独立样本T检验分析,采用Duncans检验进行多重比较和显著分析,显著性水平为P<0.05,极显著水平为P<0.01。
2 结果与分析
2.1 兔舍温湿指数的测定
如图1 所示,适温组每日平均ITH指数均低于27.8,表明该组母兔整个试验期均处于无热应激状态;热应激组每日平均ITH指数均高于30,由此判断试验期间该组母兔处于特别严重热应激状态。

图1 试验期间不同兔舍温湿指数日变化曲线
2.2 热应激对齐兴肉兔初产母兔繁殖性能的影响
由表1 可知,与适温组相比,热应激组的情期受胎率、胎产仔数、初生窝重以及断奶成活率均降低,但差异不显著(P>0.05)。而热应激组的21 d窝重和28d断奶个体重显著低于适温组(P<0.05)。

表1 热应激对齐兴肉兔初产母兔繁殖性能的影响
2.3 热应激对齐兴肉兔初产母兔血清GOT、GPT活力的影响
由图2可知,适温组和热应激组兔在妊娠期G0~G28时,血清GOT活力均呈上升趋势;哺乳期L0~L28时,血清GOT活力均呈下降趋势;适温组和热应激组兔血清GOT活力均在G28达到顶峰。两组间相比较,热应激组兔血清GOT活力在G21、G28和L0时间点显著高于适温组(P<0.05),而在其他时间点两组之间差异不显著。

图2 热应激条件下齐兴肉兔初产母兔血清GOT活力变化
由图3可知,整个试验期间,适温组和热应激组兔血清GPT活力的变化模式基本一致,呈现出先下降后升高,再基本维持不变。两组之间相比,热应激组兔血清GPT活力在G0和G7时间点显著高于适温组(P<0.05),而其他时间点差异不显著。
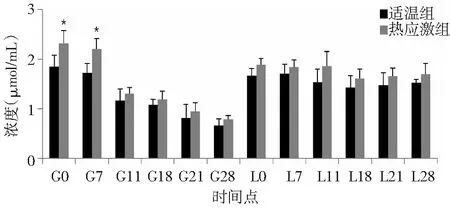
图3 热应激条件下齐兴肉兔初产母兔血清GPT活力变化
2.4 热应激对齐兴肉兔初产母兔血清免疫指标的影响
由图4可知,整个试验期间,适温组和热应激组兔血清皮质醇含量都是呈现出先升高后降低变化趋势,与适温组相比,在G11~L0时间点,热应激组兔血清皮质醇含量显著升高(P<0.05),在其他时间点热应激组高于适温组,差异不显著。

图4 热应激条件下齐兴肉兔初产母兔血清皮质醇含量变化
由图5可知,整个试验期间,适温组和热应激组兔血清T3含量变化规律较为一致,都是呈现出现先下降后升高再降低的趋势。与适温组相比,热应激组兔血清T3含量均有不同程度下降,除在时间点 G11和G18显著下降外(P<0.05),其他时间点均不显著低于适温组。
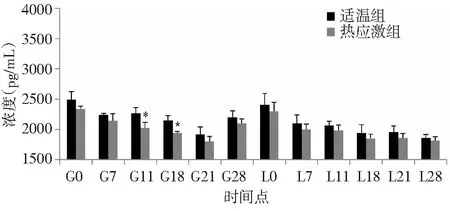
图5 热应激条件下齐兴肉兔初产母兔血清T3含量变化
由图6~7可知,整个试验期间,适温组和热应激组母兔血清IgA、IgG浓度均较稳定,没有显著变化。两组之间相比较,热应激组母兔血清IgA的含量均低于适温组,除时间点 G18显著下降外(P<0.05),其他时间点两组之间差异不显著。而对于血清IgG的含量,在各个时间点热应激组均低于适温组,但差异不显著。

图6 热应激条件下齐兴肉兔初产母兔血清IgA含量变化

图7 热应激条件下齐兴肉兔初产母兔血清IgG含量变化
2.5 热应激对齐兴肉兔初产母兔生殖激素含量的影响
由图8可知,整个试验期间,适温组和热应激组兔血清E2含量变化规律基本一致,在配种当天,血清E2含量较低,然后快速升高在G11时达到最高值,再慢慢下降在产后当天(L0)时达到最低值。从L0到L28时,也都是先升高后降低。与适温组相比,热应激组兔血清E2含量均有不同程度下降,差异不显著。
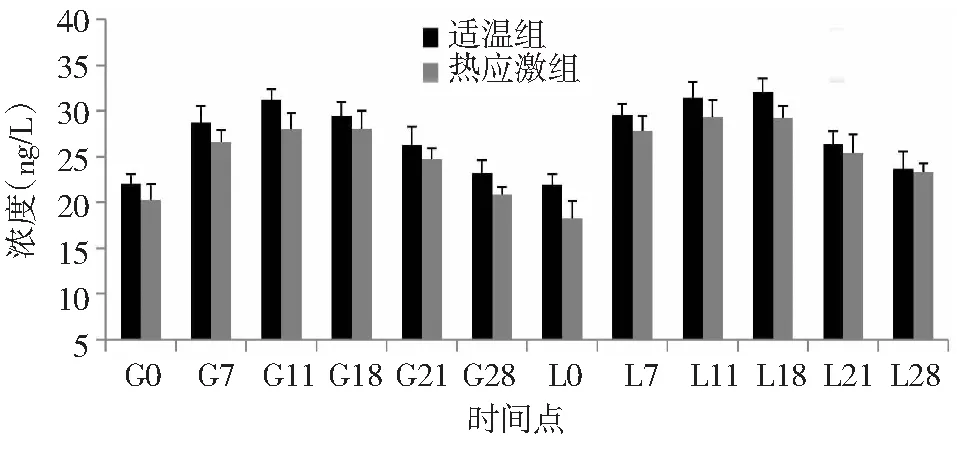
图8 热应激条件下齐兴肉兔初产母兔血清E2含量变化
由图9可知,在妊娠期G0到L0,适温组和热应激组兔血清P4含量先升高后下降,在G11时达到最高值。在母兔产后,血清P4含量较为稳定,维持基础值水平。在整个试验期间,与适温组相比,除G11、L0和L28 3个时间点热应激组兔血清P4含量显著低于适温组外(P<0.05),其余时间点均差异不显著。

图9 热应激条件下齐兴肉兔初产母兔血清P4含量变化
由图10可知,在妊娠期G0到G28,适温组和热应激组兔血清PRL含量较为稳定,均维持在较低水平,在产后当天(L0)时含量急剧升高,而从L0到L28,均先升高后降低,在L11时达到峰值。与适温组相比,整个妊娠期间,热应激组兔血清PRL含量与适温组差异不显著。而产后L0到L28,除L11和L18 2个时间点热应激组兔血清PRL含量显著低于适温组外(P<0.05),其余时间点均差异不显著。

图10 热应激条件下齐兴肉兔初产母兔血清PRL含量变化
3 讨 论
3.1 热应激对齐兴肉兔初产母兔繁殖性能的影响
热应激被认为是动物福利、生产繁殖的限制因素,它会导致山羊、绵羊、兔等哺乳动物繁殖等生产性能降低[15-16]。研究表明,在外界环境达到30 ℃以上时,新西兰兔死亡率较28 ℃以下时显著增高。同时,热应激会使母畜发情期缩短或者发情表现不明显影响适时配种,从而降低受胎率[17-18]。也有研究表明:妊娠期热应激对母兔产仔数、仔兔初生重和死胎率均有负面影响[4]。本试验中,热应激组母兔情期受胎率、胎产仔数、初生窝重以及断奶成活率比适温组有所降低,但差异不显著,而热应激组母兔的21 d窝重和28 d断奶个体重显著低于适温组。这与其他人的研究结果基本一致。
3.2 热应激对齐兴肉兔初产母兔血清酶活力的影响
急性热应激主要导致动物机体血浆代谢物的紊乱,而慢性热应激则通过血浆乳酸脱氢酶(LDH)、谷草转氨酶(GOT)、血清激酶(CK)的升高引起组织损伤,同时GPT浓度也会明显升高[19]。Xie等[20]研究发现,经过6~8周的热应激,肉鸡血浆GOT活性增加,可能是由于肝脏和肌肉损伤的原因。Marder等[21]认为兔发生热应激两小时后,血清GOT水平的升高可能是热应激对肌肉组织细胞造成了损害。本试验中,适温组和热应激组兔血清GOT活力呈先上升后下降趋势,其中热应激组兔血清GOT活力在G21~L0时显著高于适温组。而适温组和热应激组兔血清GPT活力的变化呈现出先下降后升高趋势,且热应激组兔血清GPT活力在G0和G7时间点显著高于适温组。这说明热应激会引起血清中这两种酶活力上升,推测这是由于热应激对肝脏和肌肉损伤引起的。
3.3 热应激对齐兴肉兔初产母兔血清免疫指标的影响
环境温度升高会刺激下丘脑-垂体-肾上腺轴,最终产生皮质醇应对热应激所产生的不利影响;而动物体温升高同时会刺激下丘脑-垂体-甲状腺轴应激系统,降低T3的分泌量,从而降低能量代谢[22-23]。Bharati等[24]研究发现特伯卡牛血清皮质醇浓度在短期热应激中升高,在长期热应激中下降。本研究中,整个试验期间,适温组和热应激组兔血清皮质醇含量都是呈现出先升高后降低变化趋势,在妊娠12 d血清皮质醇含量达到最高值。随着热应激时间的延长,血清皮质醇的水平持续下降,与适温组相比,在G11~L0时间点,热应激组兔血清皮质醇含量显著升高。同时,与适温组相比,热应激组兔血清T3含量均有不同程度下降,除在时间点 G11和G18显著下降外,其他时间点均差异不显著,推测这可能与齐兴肉兔对热应激环境的适应能力有关[23]。
热应激会抑制机体免疫器官和淋巴组织的蛋白质合成,降低体液免疫和细胞免疫功能[25]。免疫球蛋白IgG和IgA的变化可以代表机体体液免疫的状况。整个试验期间,适温组和热应激组母兔血清IgA、IgG浓度均较稳定。两组之间相比较,热应激组母兔血清IgA和IgG含量均低于适温组。这可能与齐兴肉兔耐热性能好,体液免疫功能强有关。
3.4 热应激对齐兴肉兔初产母兔生殖激素含量的影响
动物生殖器官的发育及卵泡发育主要受下丘脑-垂体-性腺轴调控,其中在很大程度上依赖于下丘脑GnRH的脉冲方式释放。在鸡上研究表明,热应激会导致机体的内分泌生理失调,首先减少下丘脑GnRH分泌,使垂体GTH生成减少,并导致LH、FSH、E2和P4含量显著降低[26]。本研究结果表明,与适温组相比,热应激组兔血清E2含量均有不同程度下降,差异不显著。对于血清P4含量,除G11、L0和L28 3个时间点热应激组兔血清P4含量显著低于适温组外,其余时间点均差异不显著。整个妊娠期间,热应激组兔血清PRL含量与适温组差异不显著,而产后L0到L28,除L11和L18两个时间点热应激组兔血清PRL含量显著低于适温组外(P<0.05),其余时间点差异不显著。这与其他物种上的研究基本一致,然而与适温组相比,热应激组母兔生殖激素含量下降不明显,这可能与齐兴肉兔耐热性好有关。
4 结 论
在热应激条件下齐兴肉兔母兔表现出较好的繁殖特性,这可能与其血清生化指标和生殖激素的变化相关,该研究结果为选育耐热性好的新品种提供了依据。
猜你喜欢
今日农业(2022年2期)2022-11-16
中国急救医学(2022年2期)2022-11-15
四川畜牧兽医(2022年10期)2022-10-28
健康之家(2020年7期)2020-11-02
考试与评价·高二版(2020年1期)2020-09-10
疯狂英语·新读写(2020年3期)2020-06-06
家庭科学·新健康(2018年12期)2018-03-01
中国体育教练员(2017年2期)2017-07-31
农家科技(2016年4期)2016-06-21
农村农业农民·B版(2011年5期)2011-06-16
