
元谋干热河谷侵蚀沟不同部位土壤含水量与植物多样性关系研究
2021-08-05 14:01苟嘉皓吴世祥胡翠华刘守江
西南农业学报 2021年6期
苟嘉皓,吴世祥,何 聪,胡翠华,杨 丹,2,刘守江,2,3
(1.西华师范大学国土资源学院,四川 南充 637009;2.四川省干旱河谷土壤侵蚀监测与控制工程实验室,四川 南充 637009;3.西华师范大学嘉陵江流域研究所,四川 南充 637009)
【研究意义】元谋干热河谷位于我国西南地区,该地区侵蚀沟充分发育,大面积发生沟蚀崩塌,水土流失严重。并且该地区气候干燥,降雨少且年降雨量分配不均,蒸发量大,土壤水分较低。植物生长与土壤情况密切相关,而土壤水分是影响植物生长发育的重要因素之一,尤其在干热河谷地区,土壤水分是制约当地植被恢复的主要限制因子[1],在侵蚀沟发育的元谋干热河谷地区,植被恢复[2]和生态保护对治理当地水土流失、恢复当地生态环境尤为重要,故对当地土壤含水量与植被关系的研究显得非常必要。【前人研究进展】从文献查询结果看,元谋干热河谷针对土壤部分主要集中在土壤质量评价[3]、土壤机械组成[4]、土壤水分研究[5-6]等方面,在植被方面主要有冲沟形态[7-8]、微地形[9]等因素的影响。而对于该区土壤水分与植物多样性的关系研究较少。【本研究切入点】本研究从元谋干热河谷典型侵蚀沟的土壤水分情况入手,分析土壤含水量对植物生长的反馈,研究该区不同部位土壤含水量与植物多样性关系,为元谋干热河谷的生态恢复与水土保持工作提供参考和借鉴。【拟解决的关键问题】通过元谋干热河谷不同部位土壤含水量与植物多样性指数之间的相关性,以分析当地水分对植物群落的影响程度,为侵蚀沟内植被恢复与冲沟有效治理提供依据。
1 材料与方法
1.1 研究区概况
元谋干热河谷位于滇中高原北部的金沙江一级支流一龙川江下游,介于东经101°35′~102°05′、北纬25°25′~26°07′,海拔位置980~1400 m。研究区选取的是云南省楚雄自治州元谋县沙地村、金雷村、小雷宰3个样区。样区多年平均气温21.9 ℃,最冷月(12月)均温15.4 ℃,最热月(5月)均温27.2 ℃,日照时数2550~2744 h,日照百分率为60%,光照充足,热量条件好且蒸发量大;年降雨量600.9 mm,表现为夏季降雨少而集中,气候炎热干燥。自然植被方面为稀树灌草丛,禾草为主,乔木稀少,灌木草本发育较好,乔木以桉树(EucalyptusrobustaSmith)为主,灌木以车桑子[Dodonaeaviscosa(L.)Jacq.]、银合欢[Leucaenaleucocephala(Lam.)de Wit.]和小桐子(JatrophacarcasL.)为主,草本主要为扭黄茅[Heteropogoncontortus(L.)Beauv.]、茅草[Imperatacylindrica(L.)Beauv.]和孔颖草[Bothriochloapertusa(L.)A.camus.]等[10]。土壤以燥红土和变性土为主[9]。
1.2 研究方法
样方设置与数据采集,研究样区位于元谋干热河谷侵蚀沟发育较为典型的沙地村、金雷村、小雷宰,根据研究样区侵蚀沟与植被的实际状况,于2019年7月中旬进行了为期10 d的野外调查与数据收集。分别在沙地村、金雷村和小雷宰地区选取4、3和5条典型侵蚀沟作为土壤和植被的采样区。其中,植被数据通过样方法进行采集,在每条侵蚀沟内设置10 m×10 m的乔木样方1个,并在每个乔木样方内设置5 m×5 m的灌木样方2个,然后在每个灌木样方内设置以1 m×1 m草本样方2个,3个样区共计设置了72个植物样方,统计其植物物种数、各物种的株数,并对植物盖度、高度等指标进行观测和记录,研究区植被基本情况如表1所示。为了提高土壤数据可靠性和精度,利用环刀在每条侵蚀沟的集水区、沟头、沟床分别各采集2个表层土壤,其中1个用于重复实验,每条侵蚀沟共计6个环刀土样,3个样区共计72个土样。将采集的土壤带至室内,在土壤实验室内采用烘干法进行土样处理,获得土壤的干重,计算出每个样地对应不同部位的土壤含水量。
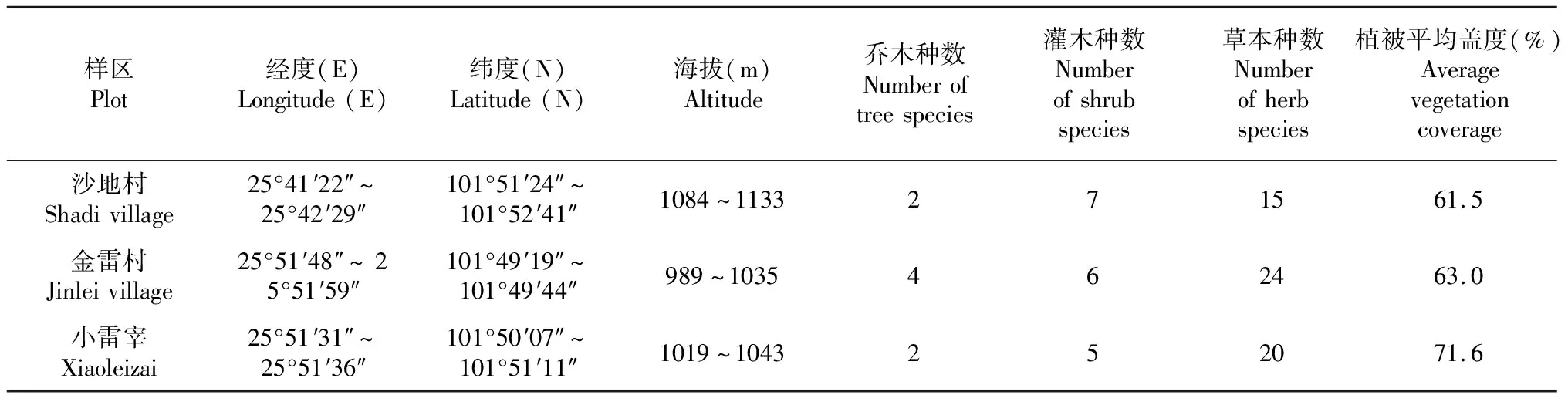
表1 研究区植被基本情况
1.3 计算公式
植物多样性指数可用来反映植物物种丰富程度和植物的生长状况,其中主要有Margalef丰富度指数Ma、香浓维纳指数H′、辛普森指数D、Pielou均匀度指数E等[11]。
Margalef丰富度指:
Ma=(S-1)/lnN
(1)
Shannon-Wiener指数:
H′=-∑(PilnPi)
(2)
Simpson指数:
(3)
Pielou 均匀度指数:
E=H/Hmax
(4)
式中,Pi为第i种个体数占群落中总个体数的比例,S为群落中物种总数,N为观察到的个体数,H为实际观察的物种多样性指数,Hmax为最大的物种多样性指数。
土壤含水量(%)=(原土重-烘干土重)/烘干土重×100%= 水重/烘干土重×100%
采用Excel软件对所获取的野外数据进行处理,并使用OriginPro 9拟合趋势线,运用SPSS22.0软件对数据进行相关性分析,为元谋干热河谷植物多样性与土壤含水量之间的关系研究提供参考。
2 结果与分析
2.1 侵蚀沟不同部位土壤含水量与植物多样性指数基本情况
由表2可知,研究样区侵蚀沟集水区的土壤含水量在4.49%~18.59%;沟头土壤含水量在3.69%~17.36%;沟床土壤含水量在2.51%~20.96%;土壤总体平均含水量在4.33%~17.29%。由此可见,研究样区集水区、沟头、沟床、总体平均含水量均在25%以下,表明研究样区土壤较为干旱,说明水分成为植物分布和生长的限制因子,对植物生长发育具有重要的影响。

表2 各侵蚀沟不同部位土壤含水量基本情况
由图1可知,3个样区中,小雷宰的总体土壤含水量大大低于沙地村与金雷村,可能是因为小雷宰地区植被盖度较大,生长发育较好,植被对土壤水分有更多的需求,土壤含水量也就更低;沙地村的土壤含水量大于小雷宰,可能是因为沙地村位于城郊且靠近龙川江,由此获得的降雨量和水分均大于小雷宰;但沙地村的土壤含水量较金雷村相比要低,可能是因为沙地村植被物种较金雷村丰富,沙地村主要以燥红土为主,而金雷村为变性土,燥红土比变性土更有利于植被的生长[9],因此植被生长发育较好,尤其是有桉树等需水量较大的植被生长,土壤中的水分可能被植被吸收较多,导致该地区土壤含水量不及金雷村,所以沙地村土壤含水量<金雷村土壤含水量。在总体上,侵蚀沟各部位的平均含水量情况:沟床(12%)>沟头(11%)>集水区(10%),这与吴汉[5]等前人的研究不完全一致,原因可能是调查期间刚好是雨季,沟床积水偏多,土壤含水量总体提高所致。

图1 侵蚀沟不同部位平均含水量区域分异
研究区共调查到36种植物,每个样方调查到的植物物种数在10~14,平均每个样方约有12个物种;研究区调查到乔木、灌木、草本植物共计1132株,每个样方株树数在56~125,平均每个样方约有94株,最高最低差69株。研究区侵蚀沟植物多样性指数基本情况见表3所示。研究区的丰富度指数Ma数值范围为1.890~3.040,平均值为2.470,极差值为1.150;香浓维纳指数H′范围为1.841~2.289,平均值为2.094,极差值为0.448;辛普森指数D范围为0.787~0.870,平均值为0.836,极差值为0.083;Pielou均匀度指数E范围为0.794~0.904,平均值为0.840,极差值为0.110。由此可见,研究区植物物种总体较少,而且分布较为不均衡,侵蚀沟生境较为脆弱,受到外界因素影响较大,侵蚀沟生态环境亟待恢复与治理。在3个样区中,沙地村的香浓维纳指数H′、辛普森指数D、Pielou均匀度指数E的平均值均优于其他两地,原因在于沙地村相对金雷村和小雷宰而言,获取龙川江的水分更为容易,植物群落乔灌草层较为完整,植物群落更为稳定。小雷宰丰富度指数Ma、香浓维纳指数H′的极差值均为最大,说明该区植物物种数目分布不均衡、群落结构复杂程度差异较大。

表3 植物多样性指数基本情况
2.2 不同部位土壤含水量与植物多样性指数相关分析
利用SPSS22.0分析软件,在数据基本符合正态分布的前提下,对侵蚀沟不同部位含水量与植物的多样性指数进行相关分析,分析其间的关系。从表4可知,在所选样区,土壤含水量与植物多样性的相关性总体趋势上以正相关为主,表明土壤含水量越高有助于当地植物物种的分布和生长发育,但显著性较低,可能是由于调查时间在雨季,侵蚀沟水土流失较为严重,地形和降水对植被的影响更为显著。通过各多样性指数与不同部位含水量相关系数可看出,沟床含水量与丰富度指数Ma的相关性呈负相关(-0.146),这可能是因为丰富度指数Ma反映的是植被的组成,与物种数成正比,与个体分布情况无关[12],沟床含水量较其他部位高,植物优势种对水的利用更为有效,生长更为迅速,从而占据了其他物种的生境条件;沟头含水量与均匀度指数E的相关性也呈现负相关(-0.164),可能是因为均匀度指数反映物种个体的相对丰富度,沟头区域灌草生长较好,土壤含水量的增大加剧了种间竞争,从而使物种间个体所占比例下降。总体上,研究样区土壤含水量对植被多样性具有一定影响,但影响较小,从中找出相对影响较大的部位,加强对该部位的后续研究。
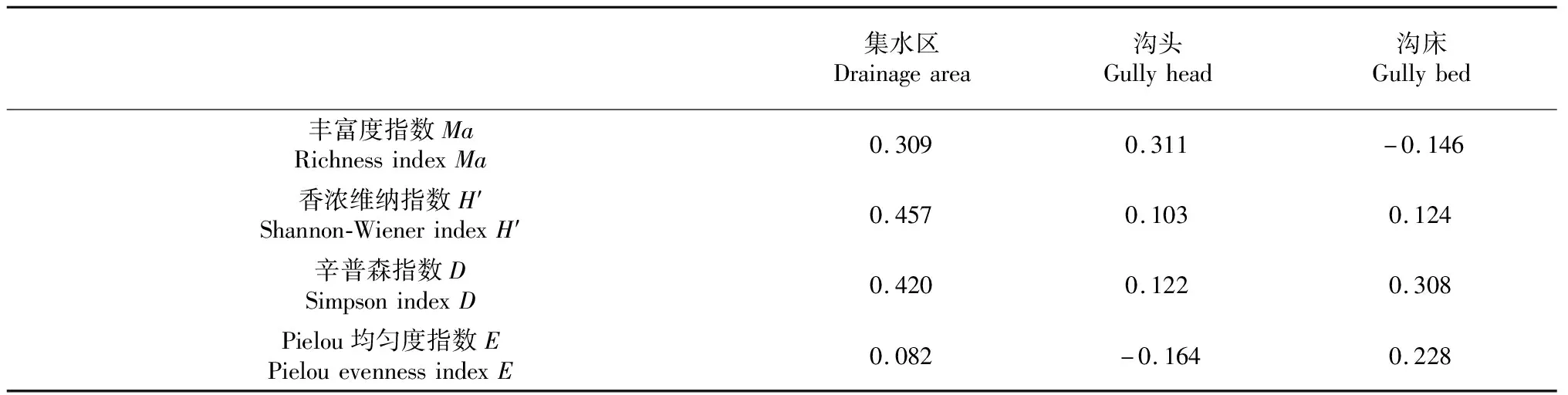
表4 侵蚀沟不同部位含水量与多样性指数Pearson相关性
2.3 趋势拟合分析
对样区不同部位土壤含水量与植物多样性进行趋势拟合分析,从图2可知,在集水区与沟床区域,丰富度指数Ma、香浓维纳指数H′、均匀度指数E以及辛普森指数D均随土壤含水量的增加呈现上升的趋势,原因可能是集水区与沟床相对沟头地势平缓,植物对水分的有效利用有助于植物的生长;在沟头区域,丰富度指数Ma、香浓维纳指数H′、辛普森指数D随含水量的增加呈现上升的趋势,而均匀度指数E呈现下降的趋势。总体上看,多样性指数的变化趋势并不明显,这说明在湿季,样地区域内土壤水分对植被的生长有一定的促进作用,但是土壤水分并非影响植被生长的关键因素,地形因素占据一部分。在湿季,降雨会造成对土壤的冲刷,发生侵蚀沟的侧蚀以及溯源侵蚀,都一定程度上影响着植被的生境。通过比较侵蚀沟3个不同部位与植物多样性的相关关系,以正相关为主,集水区部位的土壤含水量与植物多样性指数的相关性最为强烈,表明集水区部位的土壤含水量变化可能对植物物种的生长发育影响较大,需加强对集水区部位土壤含水量变化的关注。

图2 不同部位含水量与植物多样性指数的关系
2.4 各样地不同部位含水量与植物多样性指数相关分析
对样区总体上进行了不同部位含水量与植被多样性的相关分析,得出土壤含水量在时间空间的限制下对植被多样性影响较小。现从各样地之间进行土壤含水量与植被多样性的相关分析来探究不同部位在各地的影响,各部位含水量于各地植被多样性指数相关性如表5所示,对集水区部位来说,植物多样性总体上与各样地间呈正相关的关系,甚至均匀度指数E与小雷宰集水区土壤含水量呈显著正相关(P<0.05),相关系数为0.941,这说明集水区的含水量对植物生长有一定促进作用,在金雷村地区,采集的数据表明其土壤含水量为最高,其表现作用也更强。在沟头与沟床部位,金雷村的多样性指数与土壤含水量均呈负相关关系,金雷村含水量最高,沟头、沟床植被盖度虽大,但优势种对水分的利用也最强,种间斗争明显,最终表现为整体植物多样性并不明显。而在小雷宰的沟头区域,香浓维纳指数H′、辛普森指数D、Pielou均匀度指数E与含水量呈负相关关系,小雷宰含水量低,沟头部位大多长有乔木、灌木等植被,草本对水的利用情况不及乔灌,因此显现出植物多样性不强。在沙地村的沟床区域,丰富度指数Ma与含水量呈显著负相关(-0.982),沟床对水的截留较多,土壤含水量较高,当地的优势种生长茂盛,导致样地沟床植物种类丰富度程度不够。从整体上看,不同部位的土壤含水量与样地植被多样性大多呈不显著相关。通过分样地对总体情况的验证,由此可见,仅从土壤含水量的角度来看,对植物多样性的影响还是较小且有限的。
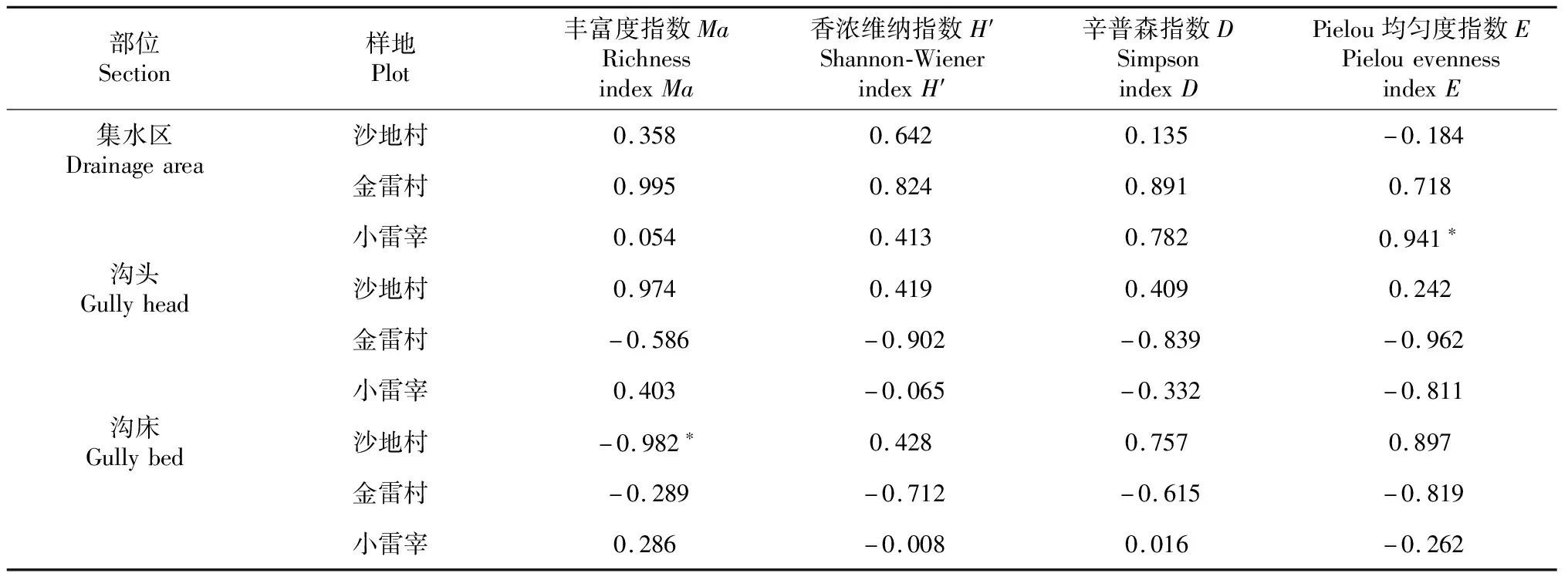
表5 各部位含水量与各地植被多样性指数Pearson相关性
3 结 论
通过对3个样地的36个样方的调查结果,以及从整体和区域性的对侵蚀沟不同部位土壤含水量与植物多样性指数之间的相关性分析,得出以下结论。
不同部位土壤含水量分别为集水区(9.59%)、沟头(10.11%)、沟床(11.31%),研究样区土壤较为干旱。
研究样区的丰富度指数Ma数值平均值为2.470;香浓维纳指数H′平均值为2.094;辛普森指数D平均值为0.836;Pielou均匀度指数E平均值为0.840。研究样区植物物种较少,分布不均,生态较为脆弱。
在雨季,从总体趋势上看,集水区、沟头、沟床含水量与丰富度指数Ma、香浓维纳指数H′、辛普森指数D、Pielou均匀度指数E均为不显著相关,大多呈正相关,从影响程度上看,可以加强对集水区部位土壤含水量的关注。从各样地情况来看,仅沙地村沟床与丰富度指数Ma、小雷宰集水区与Pielou均匀度指数E呈现显著相关,其余皆为不显著相关。表明各样地之间,不同部位含水量对植物多样性影响较小的。
此次研究不足之处限于时间空间及人力,调查起来较为困难,没有对元谋干热河谷整体上做详尽的调查,且土壤样方显得颇少。针对元谋干热河谷地区,侵蚀沟内部地形破碎,地势条件也尤为复杂[7],植被的研究对于当地生态恢复有重大意义。植被的恢复情况有助于减缓侵蚀沟的水土流失,从不同部位的土壤水分着手来研究植物多样性情况,利于人工恢复植被对种植植被的把控,为其研究提供参考。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
科学技术创新(2022年30期)2022-10-21
恋爱婚姻家庭·养生版(2021年10期)2021-10-28
课外生活·趣知识(2021年2期)2021-05-24
学校教育研究(2021年24期)2021-03-28
少儿科学周刊·少年版(2021年20期)2021-01-17
农家科技(2020年3期)2020-05-11
湖北畜牧兽医(2017年4期)2017-05-18
哈尔滨理工大学学报(2016年2期)2016-09-12
养禽与禽病防治(2012年9期)2012-08-01
