
不同种源多油辣木的形态变异分析
2021-08-04 07:31贺思腾魏静吴疆翀郑益兴张燕平
热带作物学报 2021年6期
关键词:聚类分析
贺思腾 魏静 吴疆翀 郑益兴 张燕平
摘 要:為揭示不同种源多油辣木(Moringa oleifera Lam.)的形态多样性、变异程度以及各种源的表型特点。对云南省引种栽培的世界范围内10个种源多油辣木的15个表型数量性状和质量性状进行差异性研究。结果表明,不同种源多油辣木的形态学特征存在广泛的变异,其中复叶长变异幅度最大,变异范围为6.052~14.325 cm,变异系数为25.192%,其次为种子千粒重、小叶长、种子长和小叶宽,而小叶对数的变异幅度最小,变异范围为4~6,变异系数为9.420%,侧脉数目和可育花药数量无表型变异;主成分分析显示多油辣木种源间的形态差异主要来源于小叶面积、小叶长、花瓣颜色、小叶叶型和种子长;UPGMA聚类分析表明,10个种源可以归为两大类群,其中中国台湾、缅甸实皆与印度南部班加罗尔的种源归为一个类群,缅甸木各具、肯尼亚、美国、古巴与印度北部新德里的种源归为另一个类群。不同种源地的多油辣木在其种源地因人为筛选或环境适应而形成了在叶片和种子的形态指标上较高的形态分化,因此,建议不同种源在良种选育时可根据利用部位的不同而选择培育方向。
关键词:多油辣木;形态变异;主成分析;聚类分析
中图分类号:S722 文献标识码:A
Morphological Variation of Different Provenances of Moringa
oleifera Lam.
HE Siteng1,2, WEI Jing1,3, WU Jiangchong1, ZHENG Yixing1, ZHANG Yanping1
1. Research Institute of Resource Insects, Chinese Academy of Forestry, Kunming, Yunnan 650233, China; 2. Puer Forest Ecosystem Research Station, National Forestry and Grassland Administration of China, Puer, Yunnan 665001, China; 3. Municipal Management and Maintenance Center of Urban Administration Bureau, Zhengzhou Airport Economic Comprehensive Experimental Zone, Zhengzhou, Henan 451162, China
Abstract: In order to reveal the morphological diversity, variation extent, and phenotypic traits among ten world-wide provenances of Moringa oleifera Lam. domesticated in Yunnan, China, a study was performed by examing the variation of 15 morphologic features. There were wide variation among the provenances and morphologic features. In all morphologic features, frond length was ranged from 6.025 to 14.325 cm, with the highest variance coefficient of 25.192%, followed by thousand-seed weight, leaflet length, seed length and leaflet width. Principal component analysis showed that the main variation was resulted from leaflet area, leaflet length, petal color, leaflet shape and seed length. Clustering analysis by the UPGMA method showed that the ten provenances could be divided into two groups. One was formed by the provenances of Taiwan of China, Sagaining, and Bangalore (South India), the other was formed by the provenances of Pakkoku, Kenya, USA, Cuba, and Bangalore Delhi (North India). M. oleifera Lam. from different provenances is highly differentiated in the leaf and seed shape index by artificial selection or environmental adaptation. Different provenances could be chosen in the breeding direction according to the different using parts in the breeding of improved varieties.
Keywords: Moringa oleifera Lam.; morphologic variation; principal component analysis; cluster analysis
DOI: 10.3969/j.issn.1000-2561.2021.06.008
物种的遗传变异是长期进化的产物,是物种生存适应和发展的前提[1-2]。遗传变异体现在不同水平上,从形态学或表型性状上检测遗传变异是最古老的也是最简便的方法[3-4]。这种方法向来是物种分类、资源鉴定以及探讨物种进化问题的基础[5],尤其是当需要在短期内对变异程度有所了解或在其他生化方法无法開展时;因此,形态学方法不失为一种有价值的选择[6-7]。形态标记具有直观、有效和测量简单等特点,是长期以来种质资源评价、育种后代选择和遗传多样性研究最基本的标记。通过观察性状并结合一定的分析方法,就可以在短期内对所研究物种的遗传变异水平有一个基本的认识[8-11]。
多油辣木(Moringa oleifera Lam.)为辣木科(Moringaceae)辣木属(Moringa)的一种多用途树种,原产印度北部西喜马拉雅地区,全株均可利用,尤以富含营养、药用和油料成分的叶、果实、种子经济价值较高[12-13]。多油辣木的花、叶和果都可以作为美味的蔬菜,干种子和幼苗的干燥根可以碾成粉末作为调味料,多油辣木的籽油可以用作食用油,多油辣木营养极为丰富,富含维生素A、B、C、E,蛋白质及钙、铁、钾等矿物质;多油辣木的种仁含油约35%~40%,油质清澈、不粘、细腻、润滑且芳香,具有良好的芳香固着性和极佳的氧化稳定性,耐贮放不易腐败[14];古印度传统医学认为,多油辣木有退热、排石、消炎、降压、利尿、止痛、催欲和强心等功用[15-16];多油辣木的另一种重要的用途是用于水的净化处理、植物油的澄清以及饮料与啤酒的过滤处理等,这由于多油辣木种仁含有一系列大量的低分子水溶性蛋白质,在溶液里呈正电性,与溶液中混浊的负电性微粒如泥沙、细菌等结合凝成絮状沉淀,再过滤除去,多油辣木净水剂可避免常用化学净水剂中的铝离子残留对人体健康的损害,从而预防阿耳茨海默氏病(Alzheimers disease)或称早老性痴呆的发生,因此在偏远地区与汛期的生活用水净化及城市日常废水处理方面有潜在的市场前景[17-18]。M. oleifera有‘PKM1和‘PKM22个栽培品种,均由印度泰米尔纳都农业大学园艺学院研究所选育。‘PKM1是通过传统方式选育的变异体,可用种子繁殖,主要特点为主干比较矮,分枝多,生长速度快,种子结实早、产量高,适宜截杆栽培,病虫害少,适应热带各种土壤类型;‘PKM2是MP31×MP28杂交产生的,该品种种子少,果荚肉香味浓,适合在印度热带平原生长,可以在结实前期与可可等热带作物间种,也可以庭院种植。多油辣木有较长的引种时间,目前多油辣木已经广泛栽培于亚洲的印度、缅甸、中国台湾,非洲的肯尼亚,以及美洲的古巴、美国等30多个热带、亚热带国家和地区[19]。
因为该资源植物具有重要的经济价值,开发潜力巨大,能够增加我国多油辣木产业的竞争力,本研究组分批次引入10个不同种源的多油辣木到云南适生地区进行引种栽培、优质良种筛选及推广试验。在此基础上本研究以这10个种源多油辣木的叶片、花和种子为材料,采取统计学的方法,对其表型性状进行观测和分析,以检测不同种源间多油辣木的形态变异程度,探索各种源的基础遗传背景,为下一步开展本土适生良种的选育和种质改良提供参考。
1 材料与方法
1.1 材料
分批次引进10个多油辣木种源的种子并培育实生苗,栽培于云南省红河流域干热河谷地区元阳县的种质保存圃内(23°17 N,102°51 E,海拔260 m)。引种地年平均气温24 ℃,最冷月均温18 ℃,最热月均温29 ℃,极端最低气温–0.1 ℃,极端最高气温43.5 ℃,≥10 ℃年积温4000~ 8700 ℃,无霜期200~364 d,年降水量770~ 860 mm。
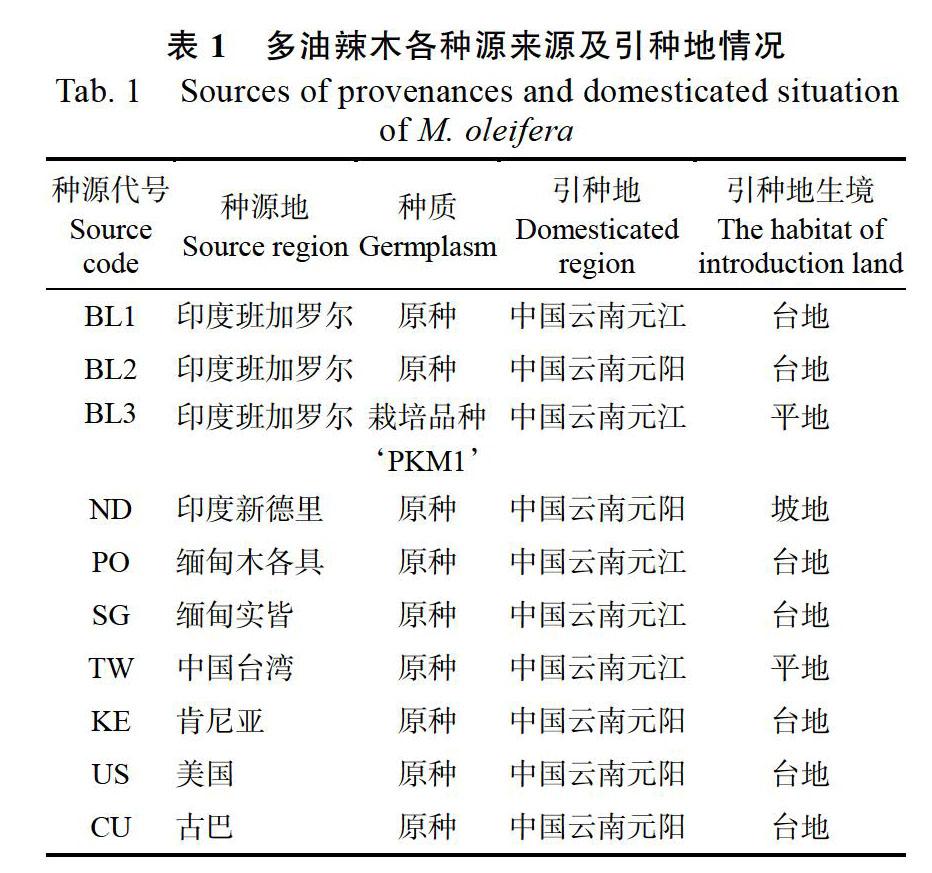
采集1~3年生的各种源植株的完全发育叶、花和种子进行形态学分析,每个种源随机抽取30株个体。种源来源和引种地情况见表1。
1.2 方法
1.2.1 表型数量性状测量 选取多油辣木的复叶长度、小叶对数、小叶长、小叶宽、小叶面积、侧脉数目、种子长、种子宽、千粒重、可育花药数量共10个性状作为连续的形态指标进行测量,其中叶面积用叶面积测量仪测量。以上各项指标在每个单株上重复测量10次。
1.2.2 表型质量性状观察与记录 选取羽状复叶类型(奇数/偶数)、小叶叶型(椭圆/近圆)、小叶尖明显程度、小叶对称性、花瓣颜色共5个性状作为离散的形态指标进行观察与记录。
1.3 数据处理
利用Excel 2007和SPSS 18.0软件对供试材料的各表型性状数据进行描述性分析和方差分析,了解变异范围、平均数、变异系数和标准差等;通过主成分分析确定各性状之间的相互关系。利用NTSYS V2.11软件计算各引种种源间的Nei遗传距离,并进行UPGMA聚类分析,了解各种源间的亲缘关系。
2 结果与分析
2.1 表型变异程度
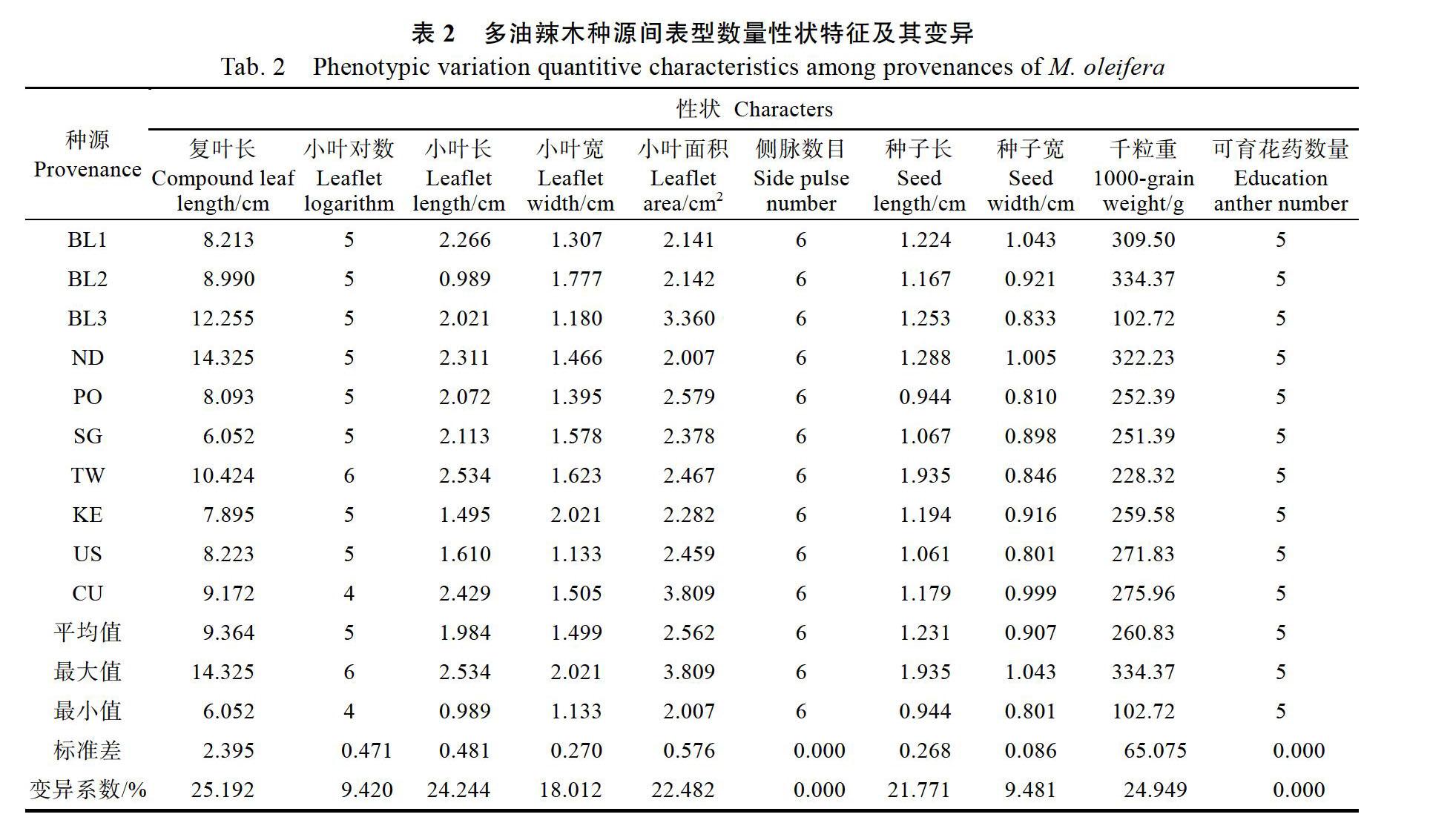
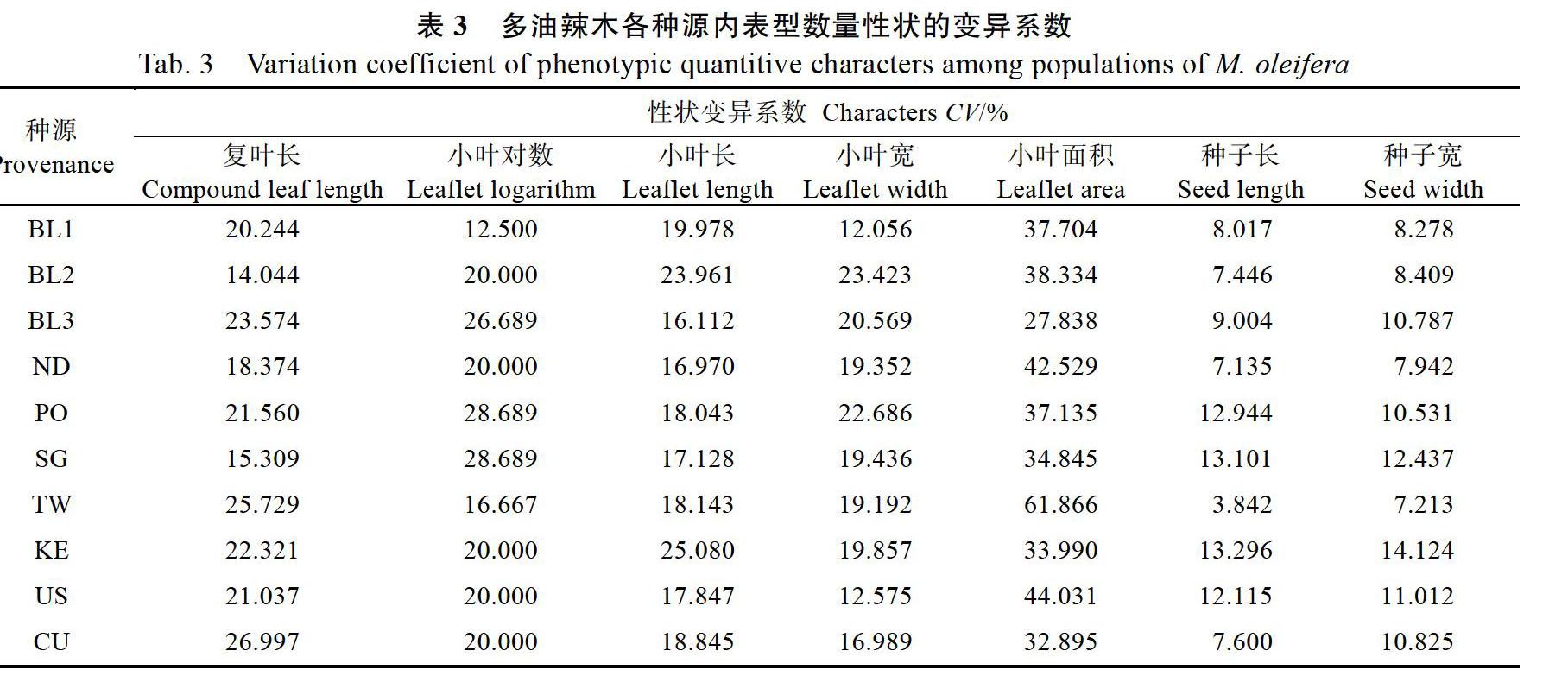
对10个多油辣木种源的15个形态学表型性状进行统计分析,在数量性状方面,不同种源间在10个数量性状上存在较为广泛的变异,变异系数分布为0%~25.192%,其中复叶长变异幅度最大,变异范围为6.052~14.325 cm,变异系数25.192%;其次为千粒重、小叶长、小叶面积、种子长和小叶宽,其变异系数分别为24.949%、24.244%、22.482%、21.771%和18.012%;变异系数较小的包括种子宽和小叶对数,二者的变异系数分别为9.481%和9.420%,而侧脉数目和可育花药数量在10个种源间没有变异(表2)。在各种源数量性状的变异中,变异系数分布为3.842%~61.866%,小叶面积的变异幅度最大,种子长的变异幅度最小(表3)。在质量性状方面,除羽状复叶类型没有区别外,小叶叶型、小叶尖明显程度、小叶对称性、花瓣颜色在种源间均存在变异(表4)。
2.2 表型性状的主成分分析
主成分分析是将多个实测变量转化为几个不相关的综合指标的多元统计方法,可以用较少的指标最大限度反映出原来较多指标反映的信息。对10个种源多油辣木的12个存在变异的表型性状(侧脉数目、可育花药数量和羽状复叶类型在种源间无变异)进行主成分分析。由表5可知,前5个主成分的累计贡献率为87.148%,根据累计贡献率≥85%的标准,不需要再增加主成分,其已經代表12个性状所提供的大部分信息,可以用于对表型变异进行综合评价。由表5和表6可知,第1主成分的贡献率为29.902%,主要由种子长、花瓣颜色、小叶对数和小叶尖明显程度决定;第2主成分的贡献率为22.047%,主要由小叶面积、小叶长和小叶对称性构成;第3主成分的贡献率为14.636%,其中小叶叶型的系数最大,且小叶叶型与小叶尖明显程度有反向趋势;第4主成分贡献率为11.898%,其中以种子宽系数最大;第5主成分的构成中主要是小叶宽,其贡献率为8.665%。在前5个主成分中,小叶面积、小叶长、花瓣颜色、小叶叶型和种子长等性状同时也是种源间差异较大的几个性状,可见这5个性状是构成表型差异的主要因素。各形态性状之间也存在一定相关性,种子长、宽和千粒重以及小叶叶型和小叶尖明显程度都存在正相关关系,小叶面积随着小叶长、宽的变化表现出一致性,叶片越长,相应的宽度也越宽,叶面积也越大。
2.3 表型性状的聚类分析
根据15项表型特征,计算种源间的Nei遗传距离后以非加权组平均(UPGMA)法进行的聚类分析表明:10个种源可以归为两大类群,其中中国台湾(TW)、印度班加罗尔(BL)、缅甸实皆(SG)的种源归为第一类群;缅甸木各具(PO)、印度新德里(ND)、肯尼亚(KE)、美国(US)、古巴(CU)的种源归为第二类群(图1)。在第一类群中,印度班加罗尔的2个原种种源(BL1、BL2)和1个栽培品种‘PKM1种源(BL3)在形态特征上高度相似,聚为一个小分支,并与缅甸实皆种源较为接近;而中国台湾种源的形态有别于上述几个种源,单独形成一个分支。在第二类群中,缅甸木各具、印度新德里、肯尼亚种源在形态特征上高度相似,聚为一个小分支;并与美国种源较为接近;而古巴种源的形态有别于上述几个种源,单独形成一个分支。
3 讨论
形态性状受物种本身的遗传组成和所处环境两方面的影响,具有一定的稳定性和变异性。种源所处的生境是动态的,植物很难处于生长发育最适宜的环境条件,无论是气候变化还是生长发育过程所处的条件,总是要产生或大或小的差异[20]。
由于地理分布差异以及生境的多样化,种内不同种源为适应生态环境形成了不同的生态型,相应表现出丰富的形态多样性[21-22]。一般来说,种源间分化越厉害,其适应能力的差别越大[23]。本研究的材料栽培于气候条件均一的引种地苗圃中,但种源间从表型性状上仍存在较大的形态分化,表明不同种源地的多油辣木已在其种源地因人为筛选或环境适应而分化出了一定程度的形态特征,这种分化主要表现在叶和种子的形态特征上。叶是植物的营养器官,关系到营养物质的累积。多油辣木在叶和种子上存在较高的形态分化,因此对不同种源在良种选育时可根据利用部位的不同而选择培育方向。本研究建议复叶长度较长和小叶面积较高的印度班加罗尔栽培品种‘PKM1、中国台湾、古巴和新德里原种种源可作为以叶为主要产出的重点选育方向,而印度班加罗尔的2个原种及新德里原种可作为以种子为主要产出的重点选育方向。多油辣木起源于印度,曾经以野生状态分布于印度北部西喜马拉雅地区的森林中[24],之后作为蔬菜和药用植物经人工种植利用,遂以栽培和半野生状态遍布印度全境[25]。随着高品质油料用途的发现,多油辣木开始在世界范围内广泛引种栽培。从印度向东,多油辣木于19世纪传到中国,台湾是我国最早引种辣木的地区[26],并继续向东到达东南亚的菲律宾。从印度向西,同样于19世纪初经埃及进入非洲和环地中海地区,并最终传入美洲[27]。
尽管形态变异具有一定的遗传基础,人工选择和环境压力在导致形态变异中也起着重要的作用[28-30]。一方面表现为起源于印度北部的多油辣木在传播至印度全境的栽培利用过程中,形成了与印度北部种源存在一定形态特征差异的南部种源,以致多油辣木在世界范围内的各引种栽培种源分别与印度北部和南部种源聚类,形成2个形态上有差异的类群,这与本研究中10个种源聚成2个分支的结果吻合,表明我们在引种过程中已覆盖了多油辣木的2个主要原始栽培种源,引种具有代表性。另一方面,由于岛屿的海洋性气候和环境湿度等方面的特殊性,使适生的引种栽培植株与陆地上的植株产生一定的形态变化,在本研究中表现为中国台湾和古巴种源在各自类群内没有与陆地上的种源聚在一起,而是单独形成一个小分支,引入这2个已分化的种源证明了我们的引种工作在多样性上是比较全面的,可以基于这些种源开展我国多油辣木的良种选育和推广工作。
参考文献
[218] 葛 颂, 洪德元. 泡沙参复合体(桔梗科)的物种生物学研究[J]. 植物分类学报, 1995, 33(5): 433-443.
[219] Soltis P S, Soltis D E. Genetic variation in endemic and widespread plant species: examples from Saxifragaceae and Polystichum (Dryopteridaceae)[J]. Aliso, l991, 13: 215-223.
[220] 中国科学院生物多样性委员会. 生物多样性研究的原理与方法[M]. 北京: 科学出版社, 1994.
[221] Schaal B A, Kane S L, Rogstad S H. DNA variation in plant populations[J]. Trends in Ecology and Evolution, 1991, 6: 329-333.
[222] Stebbins G L. Chromosomal Evolution in Higher Plants[M]. London: Edward Arnold, 1971: 43-46.
[223] 葛 颂, 洪德元. 遗传多样性及其检测方法[M]//钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1994: 122-140.
[224] 虞 泓, 徐正尧, 杨彩云. 云南松居群花粉形态多态性[J]. 云南大学学报(自然科学版), 1999, 21(2): 86-89.
[225] 王洪新, 胡志昂, 钟 敏, 等. 毛乌素沙地锦鸡儿种群形态变异[J]. 生态学报, 1994, 14(4): 366-370.
[226] 葛 颂,洪德元. 泡沙参复合体(桔梗科)的物种生物学研究:I.表型的可塑性[J]. 植物分类学报, 1994, 32(6): 489-503.
[227] 孟 林, 张国芳, 高洪文. 氮磷钾施肥量对饲用菊苣生产性能的影响[J]. 草地学报, 2003, 11(4): 325-328.
[228] 葛 颂,洪德元. 濒危物种裂叶沙参及其近缘广布种泡沙参的遗传多样性研究[J]. 遗传学报, 1999, 26(4): 410-4l7.
[229] 刘昌芬. 辣木的研究现状及其开发前景[J]. 云南热作科技, 2002(3): 20-24.
[230] Ayerza R. Seed yield components, oil content, and fatty acid composition of two cultivars of moringa (Moringa oleifera Lam.) growing in the Arid Chaco of Argentina[J]. Industrial Crops & Products, 2011, 33(2): 389-394.
[231] Gupta S, Jain R, Kachhwaha S, et al. Nutritional and me-
dicinal applications of Moringa oleifera Lam.—Review of current status and future possibilities[J]. Journal of Herbal Medicine, 2017, 11: 1-11.
[232] Bhutada P R, Jadhav A J, Pinjari D V, et al. Solvent assisted extraction of oil from Moringa oleifera Lam. seeds[J]. Industrial Crops and Products, 2016, 82: 74-80.
[233] 张燕平, 段琼芬, 苏建荣. 辣木的开发与利用[J]. 热带农业科学, 2004, 24(4): 42-48.
[234] 李 东, 赵一鹤. 辣木研究现状及发展趋势[J]. 西部林业科学, 2018, 47(5): 32-38.
[235] Mahaman, Yacoubou, Abdoul, et al. Moringa oleifera (mo) alleviates homocysteine-induced alzheimers disease-like pathology[J]. Alzheimers & Dementia, 2017, 13: 272-279.
[236] Keogh M B, Elmusharaf K, Borde P, et al. Evaluation of the natural coagulant Moringa oleifera as a pretreatment for SODIS in contaminated turbid water[J]. Solar Energy, 2017, 158: 448-454.
[237] 邹金拓, 王 鑫, 周 玮, 等. 构树不同种源种子性状地理变异及其趋势分析[J]. 种子, 2020, 39(3): 62-67.
[238] 康智明, 徐晓俞, 郑开斌, 等. 木豆种质资源形态与农艺性状的多样性分析[J]. 热带亚热带植物学报, 2017, 25(1): 51-56.
[239] 郝 蕾, 张国盛, 穆喜云, 等. 北沙柳种质资源居群表型多样性[J]. 西北植物学报, 2017, 37(5): 1012-1021.
[240] 陈家宽, 杨 继. 植物进化生物学[M]. 武汉: 武漢大学出版社, 1994.
[241] Hooker J D. The flora of British India. Vol.II[M]. London: L. Reeve & Co., 1879.
[242] Selvam A B D. Distribution, phenology and utilization of Moringa oleifera Lamk.: An indigenous medicinal plant of India[J]. Journal of Economic and Taxonomic Botany, 2005, 29: 102-108.
[243] 罗云霞, 陆 斌, 石卓功. 辣木的特性与价值及其在云南引种发展的景况[J]. 西部林业科学, 2006, 35(4): 137-140.
[244] Juliana Silver. Moringa oleifera: A brief overview[R/OL]. (2011-05-15)[2020-06-02] http://www.villagevolunteers. org/wp-content/uploads/2011/05/Overview-of-the-Moringa-Tree.pdf.
[245] 谭文婧, 李 娟, 林建勇, 等. 闽楠种源苗期生长性状地理变异研究[J]. 林业与环境科学, 2019, 35(2): 48-54.
[246] 杨 继. 植物种内变态的机制及其研究方法[J]. 武汉植物研究, 1991, 20(4): 429- 434.
[247] 任红剑, 丰 震, 乔 谦, 等. 元宝枫叶片形态特征的地理变异[J]. 西北林学院学报, 2018, 33(1): 113-119.
责任编辑:沈德发
猜你喜欢
软件导刊(2016年11期)2016-12-22
科技创新导报(2016年21期)2016-12-17
对外经贸(2016年8期)2016-12-13
数学学习与研究(2016年19期)2016-11-22
商场现代化(2016年26期)2016-11-21
大经贸(2016年9期)2016-11-16
中国市场(2016年33期)2016-10-18
科技视界(2016年20期)2016-09-29
企业导报(2016年9期)2016-05-26
