
IGF1R反义非编码RNA在肝癌中的差异化表达
2021-08-03 08:03谢志海周志聪孙静锋
临床荟萃 2021年7期
谢志海,周志聪,桂 元,孙静锋
(1.广州市老人院 老年病治疗中心, 广州 510000;2.江苏省肿瘤医院 肝胆胰外科, 南京 210013;3. 南京医科大学第四附属医院 肿瘤外科, 南京 210013)
胰岛素样生长样因子1受体(insulin-like growth factor 1 receptor, IGF1R)作为胰岛素生长因子受体,其在人体发挥着重要的作用,然而,关于IGF1R基因失调对组织影响的分子机制知之甚少。在IGF1R基因内部有一条跨过启动子,同IGF1R基因编码方向完全相反的长链非编码RNA系列[1]。同我们通常所说的长链非编码核糖核酸(RNA)表达方式很不相同的是IGF1R的反义非编码RNA在人体的乳腺组织、心肌肌纤维中呈印记表达(即父源等位基因表达,母源等位基因沉默),国际上把该长链非编码RNA命名为IGF1R反义非编码RNA(IGF1 receptor antisense imprinted noncoding RNA, IGF1RAIN RNA)[2]。IGF1RNA反义非编码RNA基因在乳腺肿瘤、血液系统肿瘤以及肺部肿瘤中差异化表达同肿瘤的进展关系密切[3-5]。我们通过逆转录聚合酶链反应(reverse transcriptase polymerase chain reaction, RT-PCR)法比较IGF1RAIN RNA在肝癌癌瘤中、肝癌瘤旁组织及正常肝组织中的含量,对不同的肝癌分期及肝癌病理分期的肝癌组织中IGF1RAIN RNA含量进行比较,分析IGF1RAIN RNA含量同肝癌的关系,验证其作为肝癌标记物的可能性。
1 资料与方法
1.1病例选择 2017年1月-2020年6月手术切除的30例肝癌的癌组织及癌旁组织,癌旁组织定义为肝癌切缘外3 cm的肝组织,其中男19例,女11例, 年龄42~71岁,中位年龄63.75岁,肿瘤直径1.0~8.0 cm,中位直径5.8 cm。按照中国临床肿瘤学会(Chinese Society of Clinical Oncology, CSCO)肝癌中国指南2018版临床分期Ⅰ~Ⅱ期18例,Ⅲ~Ⅳ期12例;病理分级按WHO分级标准:高分化肝癌10例,中分化肝癌8例,低分化肝癌12例。另取同期10例肝囊肿以及6例肝血管瘤手术切除标本的切缘旁1 cm正常肝组织作为对照组。本研究经过手术单位伦理委员会批准[南京医科大学第四附属医院(20200016)],患者以及家属签署同意书准予用于实验。30例肝癌患者术前均未行其他如化疗、靶向、放疗、免疫、中药等治疗。
1.2试剂与方法
1.2.1试剂 Tri-Reagent购自Sigma-Aldrich西格玛奥德里奇(上海)贸易有限公司,三氯甲烷、异丙醇、75%乙醇、美国ABI公司Power SYBR Green PCR Mastm’mix实时PCR试剂盒及ABI 7900HT 实时PCR系统等。
1.2.2总RNA提取 (1)从液氮罐取出肝癌切除癌瘤标本,迅速转入保持低温的研钵中研磨直到完全成粉状后滴注1 ml的Trizol试剂裂解组织。(2)匀浆分相:将裂解的Trizol液1 ml转移到1.5 ml的离心管中,20 ℃静置5 min后向其中滴加三氯甲烷200 μl,混匀后继续静置3 min放置于离心机中,温度4 ℃,转速12 000 r/min条件下离心15 min。(3)RNA 沉淀:取400 μl离心后的上层清亮液体转移到另一只清洁干燥的EP管中,向其中滴注异丙醇400 μl,经过4 ℃条件下15 min静置后,以10 000 r/min快速离心15 min。(4)洗涤沉淀:离心结束后摒弃上层清亮液体后注入75%的醇1 ml,上下快速甩动EP管2次,4 ℃,以12 000 r/min离心5 min,再次摒弃上层清亮液体后将剩下的EP 管放置于清洁干燥的环境中待沉淀透亮后滴加无 RNA酶水40 μl,最后获取物保存于-70 ℃冰箱中备用。
1.2.3互补脱氧核糖核酸(cDNA)样品合成 吸取1 μg总RNA,使用Iinvitrogen公司的M-MLV逆转录酶试剂盒进行反转录。cDNA 样品置于-20 ℃保存,作为下一步RT-PCR的模板。
1.2.4IGF1RAIN RNA分析 实时定量RT-PCR测量肝癌切除癌瘤组织、瘤旁组织中IGF1RAIN RNA含量,根据模板或引物配置RT-PCR反应的混合液[IGF1RNA反义非编码RNA上游引物正向序列:5’-CGACACATGGTCCAATCACTGTT-3’;IGF1RAIN RNA下游引物正向序列:5’-AGACTCCCCTAGGACTGCCATCT-3’; GAPDH正向序列:5'-GAAGAGAGAGACCCTCACGCTG-3’;甘油醛-3-磷酸脱氢酶(GAPDH) 序列:5’-ACTGTGAGGAGGGGAGATTCAGT-3’],严格按照RT-PCR实验步骤进行操作,采用探针法检测IGF1RAIN RNA的表达值,以甘油醛-3-磷酸脱氢酶作为对照,相对表达量采用比较CT法(2-△△CT)进行数据分析。
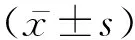
2 结 果
IGF1RAIN RNA在肝癌癌旁组织中含量较正常肝组织低(P<0.05);肝癌瘤体中IGF1RAIN RNA含量较正常肝和癌旁组织均显著下降(均P<0.05),见表1。

表1 IGF1RAIN RNA在肝癌、癌旁及正常肝组织中表达
IGF1RAIN RNA含量与肝癌者的性别、年龄以及是否有乙型肝炎史无关(均P>0.05),但其和癌瘤的大小、癌瘤分化程度及国际抗癌联盟(Union for International Cancer Control, UICC)肝癌分期有关(均P<0.05),见表2。

表2 IGF1RAIN RNA在肝癌中含量分层比较
3 讨 论
正常人组织中也存在癌瘤细胞,只是这些癌瘤细胞都处于静默的G0期,只有这些静默的癌瘤细胞启动进入G1期才能使发生癌瘤的细胞呈几何级增殖[6]。研究证实IGF1RAIN RNA在CD90+细胞肝癌干细胞中发挥促进肝癌增殖的作用[7-10]。我们前期通过RT-PCR法、Western-blot法检测了5例肝癌组织中CD45-CD90+细胞内IGF1RAIN RNA,结果显示其表达明显低于正常肝组织,这一结果提示IGF1RAIN RNA可能参与抑制CD90+肝癌的增殖,这和IGF1RAIN RNA在其他组织来源的恶性肿瘤中表达降低的特点相一致的[11]。研究发现,在乳腺癌患者样本中,IGF1RAIN RNA的含量和乳腺癌的增生及分期相关,IGF1RAIN RNA在内癌中含量较高,在进展期或者转移癌中含量则较低[12]。研究发现,IGF1RAIN RNA在白血病细胞系和高风险AML患者的血液中表达下调[13]。IGF1RAIN RNA可能在癌瘤中扮演着负向阻碍机制,其高表达时可以阻碍癌瘤的发展及转移[14]。我们通过实验发现肝癌患者的癌瘤中含量更低,分期及分化这两项条件相同的情况下,IGF1RAIN RNA随着癌瘤体积增大数值更低,而与患者年龄是否大于60岁,是否为男性,是否患有乙型肝炎没有关系,这说明IGF1RAIN RNA含量可能只和肝癌的本身恶性力有关。
IGF1RAIN RNA含量降低使得肝癌更容易发生,IGF1RAIN RNA含量正常可抑制肝癌的发展,另外,IGF1RAIN RNA含量降低失去了对肝癌侵袭力、浸润能力的阻碍作用,IGF1RAIN RNA可抑制肝癌细胞增殖、诱导肝癌细胞分化,也可能抑制肝癌的发生发展,其可能成为肝癌的早期诊断指标之一[15]。
IGF1RAIN RNA促肝癌的机制探索还处于起步阶段,更多的IGF1RAIN RNA在肝癌中的作用机制还需要深入研究,已有的研究证实IGFIR在癌瘤自分泌的信号传递中是一种重要的物质,肝癌的恶性程度及分期与IGF1RAIN RNA含量呈负相关,这对肝癌诊断及预后判断可能存在临床价值,IGFIR信号传递受阻后肝癌的发展及增殖同样出现了迟滞,这可能为肝癌治疗带来新的发现[16]。IGF1RAIN RNA在肝癌组织中含量较非肝癌组织低,其正常含量下可以抑制肝癌的发展及增殖,在分化程度及肿瘤分期的统计中,IGF1RAIN RNA与分化程度呈负相关,与分期呈正相关,具体机制尚不清楚。IGF1RAIN RNA是否可以成为抑癌基因家族中的一员还需要继续研究,其也可能是靶向抑制肝癌进展的一种新分子。
猜你喜欢
清华金融评论(2022年4期)2022-04-13
小学生学习指导·低年级(2021年6期)2021-09-10
考试与评价·七年级版(2021年4期)2021-08-14
昆明医科大学学报(2021年4期)2021-07-23
国际放射医学核医学杂志(2021年10期)2021-02-28
昆明医科大学学报(2021年1期)2021-02-07
医学概论(2021年18期)2021-01-21
房地产导刊(2020年7期)2020-08-24
中国外汇(2019年14期)2019-10-14
小学阅读指南·低年级版(2018年5期)2018-11-02
