
农田野花带群落构建及其稳定性特征
2021-07-29 08:30谢哲城高亦珂吴学峰李湛东张启翔
中国农业大学学报 2021年7期
谢哲城 高亦珂 吴学峰 徐 俊 刘 烨 李湛东 张启翔
(北京林业大学 园林学院,北京 100083)
农田野花带(Wildflower strip)是指以播种或栽植种苗的方法在农田、菜地和果园等农地边缘建植的人工植物群落。通过模拟自然草甸植物交错生长的状态,可为农田生态系统提供如净化水源、改良和修复土壤、为传粉者和害虫天敌提供蜜粉源和栖息地、改善农地生境质量与减少农药使用等多种生态系统服务[1],具有低维护和适生环境广泛等特点,在我国农业景观和绿色基础设施中有广阔的应用前景。农田野花带常使用一二年生植物与多年生植物混合播种的方式建植,但随着农田野花带应用范围的扩大,其群落建植过程中“1年好,2年衰,3年亡”的问题日益突出[2]。其中,混播群落构建过程中杂草侵入和植物消亡是影响农田野花带群落稳定性的重要因素。
杂草侵入对农田野花带的群落稳定性和功能的影响是研究热点。在清除土壤杂草种子库时,使用50 mm厚度的沙子作为播种基质能减少杂草侵入[3];极少数特定种类(如波斯菊Cosmosbipinnata和百日草Zinniaelegans等)构建的混播群落可使用专用封闭除草剂抑制土壤种子库的杂草[4],但不能满足农田野花带发展的多样化和生态需求。从种间竞争关系出发,研究者使用C-S-R植物竞争模型[5-6],或根据生态位重叠原理和功能多样性理论[7]选择和配置植物,希望增强混播群落稳定性以提高抵御杂草侵入的能力,但这些研究多停留在理论阶段。多使用莲座状植物(如报春花属Primula)能够减少杂草生长[8],但其群落结构和景观单一。目前,农田野花带群落杂草侵入问题尚未得到解决。
混播建植群落的物种多样性随时间推移逐渐降低[9-10],存在植物消亡过程。植物消亡使农田野花带稳定性降低,并释放群落空间为杂草侵入提供机会[11]。但针对混播群落植物的消亡过程开始、持续时间、程度与其对杂草侵入的影响尚未见报道。
本研究设计并持续监测8个不同物种种植比例的农田野花带群落,分析群落密度、物种多样性指数以及其中一二年生植物、短命宿根植物、长寿宿根植物和侵入植物的重要值在播种后的前4年随时间变化的趋势,并着重研究已建植6年的代表群落的变化趋势,以研究农田野花带建植初期群落构建过程规律。研究旨在从植物配置角度为建植稳定性高和功能持续的农田野花带提供参考,为解决农田野花带群落杂草侵入问题提供建议,并为农田野花带的推广应用提供理论基础。
1 材料与方法
1.1 群落设计
为充分发挥农田野花带功能,根据前人研究结果[2,12-13]以及生态位分化原理,各群落设计物种需同时满足以下2个条件:1)开花期错落。在4—10月份间,每月份至少2种植物开花;2)株高层次错落。且早花物种(7月15日以前开花)应处于群落下层(0~40 cm)或中层(40~60 cm),晚花物种(7月15日以后开花)应处于群落中层或上层(60 cm以上)。在此基础上,设计了1个一二年生植物群落(群落A),1个多年生植物群落(群落B),3个一二年生植物比例介于40%~50%的植物群落(群落C、D和H),3个一二年生植物比例介于15%~25%的植物群落(群落E、F和G)(表1)。
设计单个群落植物数量为14~25种,共54种常用混播植物(表1),其中一二年生植物22种,多年生植物32种。野生种白头翁(PulsatillaChinensis)、蓟罂粟(Argemonemexicana)、高山罂粟(Papaveralpinum)、瓣蕊唐松草(Thalictrumpetaloideum)和蓝刺头(Echinopslatifolius)的种子从野外采集获得,黄花菜(Hemerocalliscitrina)、射干(Belamcandachinensis)和西伯利亚鸢尾(Irissibirica)的种子来自于课题组栽培,其余植物种子来自北林科技股份有限公司。

表1 群落A~H设计种植植物的组成及占比Table 1 Designed species composition and proportion in community A-H
多年生植物中存在短命型植物和长寿型植物2种类型,而短命型植物可能在早期影响野花带群落植物构建过程规律及稳定性特征,但这在前期群落设计时并未单独考虑。为了进一步探究短命型宿根植物对野花带群落的影响,通过前期试验调查和查询TRY全球植物性状数据库(Plant trait database)[14]获取常用混播植物的寿命数据,并分为3个组别,即一二年生植物(Annuals and biennials,AB)、寿命不超过6年的短命宿根植物(Short-live perennials,SP)和寿命超过6年的长寿宿根植物(Long-live perennials,LP)。在上述8个群落中设计的32种多年生植物中,包含短命宿根植物11种,长寿宿根植物21种。
各群落在建植后均出现通过种子繁殖的非设计植物,其种子来源于邻近样地或单种栽培试验田。对于人工群落而言,侵入植物也是群落构建过程中不可忽略的组分。在本研究中,通过种子繁殖侵入群落的植物无法被消除,故调查侵入植物(Invasive plants,IP)并在后续分析中单独作为1个组别。
1.2 群落建植布局与数据采集
8个群落建植于北京市2个相距18 km且气候和环境条件相近的试验田内。所有样地均位于全日照环境下。2012年春季将群落H播种于试验地2(40°08′57.51″ N,116°26′24.63″ E)内1个5 m×10 m
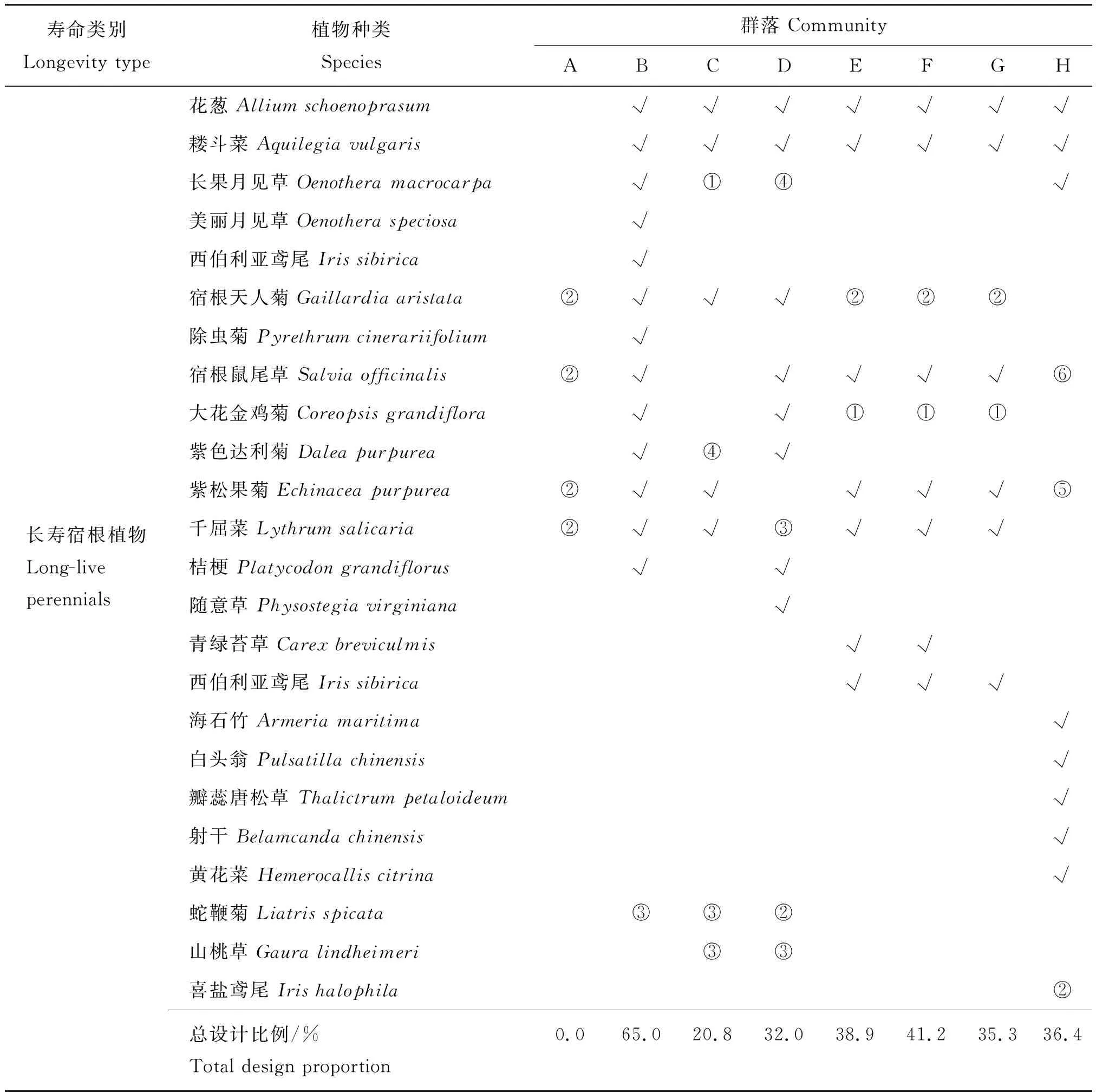
表1(续)
的样地,2015年春季将群落A~G分别播种于于试验地1(40°0′16.36″ N,116°19′54.05″ E)内7个5 m×5 m的样地(图1)。样地土壤种子库均为北京常见农田杂草,在播种前1个月对各样地充分灌溉,而后每隔1周清除1次萌发的本底杂草。样地内设置50 cm的保护行,并在周围使用80 cm园艺地布覆盖,以避免邻近样地中的植物通过无性繁殖侵入。各样地栽培管理方法相同。播种后每年春季3—5月每15日灌溉1次,每年土壤上冻前修剪地上部分并浇灌冻水,其余时间均不灌溉、不刈割;仅在播种后第1年冬季进行覆盖保护。
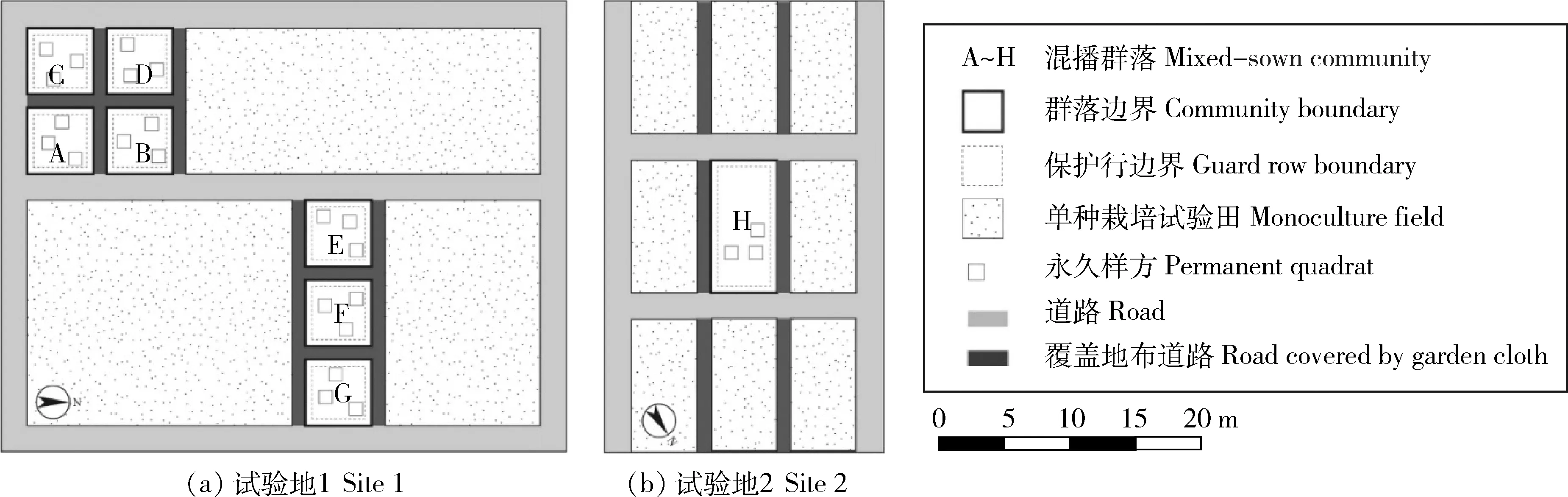
图1 试验样地布局Fig.1 Layout of sample plots
每个群落内随机设置3个1 m×1 m的永久样方,共计24个永久样方。各群落在播种后经历的生长季,按顺序命名为第1生长季、第2生长季……第6生长季,在本文图表中使用罗马数字“i,ii,…,vi”表示各生长季序次。每个生长季内的04-15、05-05、05-25、06-15、07-05、07-25、08-15、09-05、09-25和10-15,全面调查样方内出现的物种及其密度,各个物种随机抽样9株调查株高和冠幅,1个生长季内共测量10次。其中同种植物真叶>5枚与≤5枚的密度、株高和冠幅分别调查;未脱离母体的无性系不单独调查。在本文图表中使用阿拉伯数字“1,2,…,10”表示同一生长季中的各调查序次。群落A在2个生长季后明显退化,剩余植物均为侵入植物,故仅监测至第2生长季。
1.3 重要值和物种多样性指数计算
将不同组别内的所有物种重要值加和代表该组别在群落中的重要值。各物种重要值的计算公式为:

(1)
式中:Hi为物种i在群落3个永久样方中的平均高度,cm;Di为物种i在群落3个永久样方中的平均盖度,cm2;Ci为物种i在群落3个永久样方中的平均密度,株/m2;S为群落中出现的物种数量,个。
使用Simpson多样性指数(D)表征群落物种多样性,计算公式为:
(2)
式中:S为样方中出现的物种数量,个;Pi为物种i的相对丰度。每个样方的D值单独计算,取1个生长季内10个测量时间点D值的平均值代表该样方在某一年份的物种多样性指数,并取3个永久样方的平均值代表该群落在该年份的物种多样性指数。
使用Pearson相关性指数检验两组数据之间是否具有相关性,并对结果进行双侧显著性检验。同一群落在不同生长季的群落特征值的平均值(n=3),将使用Duncan法进行多重比较,置信区间设置为95%,以此检验不同生长季的平均值间是否具有显著性差异。
数据的统计分析在IBM SPSS 21.0中进行,图表采用Microsoft Excel 2016和Adobe Illustrator 2020绘制。
2 结果与分析
2.1 农田野花带群落密度年变化
第1生长季,各群落平均密度峰值均出现在7月5日(第5次调查)以前,而后开始下降(图2(a)~(h))。这主要是因为幼苗密度超过群落可容纳的密度,出现竞争淘汰现象。一二年生植物比例为100%的群落A在生长季末(10月15日)的平均密度仅为其峰值(5月5日)的8.3%(图2(a)),而一二年生植物比例仅为5%的群落B在生长季末的平均密度为其峰值(7月5日)的83.9%(图2(b)),其余一二年生植物比例介于15%~50%的群落C~H,在生长季末的平均密度为其峰值的38.4%~56.2%(图2(c)~(h))。群落初始设计的AB比例越高,在第1生长季末期的密度降幅越大,表明各群落在第1生长季平均密度的下降,还与AB植物在开花后枯萎死亡有关。

罗马数字ⅰ~ⅵ代表生长季序次。下同。误差线为使用3个样本数据计算的标准误差。仅第1生长季的第1~10次测量的密度数据中含真叶不超过5枚的幼苗。Roman numerals mean the order of growing seasons (GS). The same below. Error bars represent standard deviation among 3 quadrats. Only 1st to 10th observed density data in 1st growing season of each community include individuals with no more than 5 true leaves.图2 群落A~H在各调查序次的平均密度Fig.2 Mean densities of community A-H at each order of survey
第2生长季,仅由一二年生植物组成的群落A在5月25日达到352株/m2的峰值,在9月25日达到13株/的谷值,但相比第1生长季的峰值(482株/m2)和谷值(40株/m2)均有所下降(图2(a))。表明AB通过自播繁衍,其种子萌发、幼苗竞争、快速建植和植株开花后死亡等过程在第2生长季中重复发生,同时群落密度减小。各群落第2生长季的年平均密度与第1生长季10月15日平均密度显著正相关(Pearson相关系数为0.830,P<0.05)。即群落在第1生长季未死亡而剩余的多年生植物越多,在第2生长季具有更高的密度。且除群落A以外的其余群落,不同调查序次间的平均密度和同一调查序次中不同样方间的密度的变异程度相比第1生长季均下降(图2(b)~(h))。表明多年生植物设计比例超过50%的群落,开始由多年生植物主导,群落密度变化在时间上与样方间均趋向稳定。
第3和4生长季,各群落相比第2生长季的密度值变化不大或小幅下降(图2(b)~(h))。但群落G在第4生长季出现7株/的密度最低值(图2(g)),或因冬春季节无降水和管理不善所致,故群落G第4生长季的数据作为异常值处理,后续不做分析。
第5生长季,群落H的平均密度(35株/m2)与第3(44株/m2)和第4生长季(30株/m2)基本持平;但在第6生长季,群落H的平均密度小幅升高(58株/m2)。这是因为在第5生长季末期群落出现了大量黑心菊和松果菊的幼苗(表1),其中一部分幼苗在第6生长季真叶数量超过5枚而被计入密度数据中,从而增加了群落密度。
以上结果表明在模拟农田野花带的管理措施时,农田野花带混播群落的密度在第1生长季因为幼苗竞争淘汰和一二年生植物的死亡而快速下降;多年生植物设计比例超过50%的群落在第2~4生长季内由多年生植物主导而群落密度相对稳定或小幅下降;侵入植物能补充群落密度。
2.2 农田野花带群落不同寿命组植物和侵入植物的重要值
混播群落第1生长季,一二年生植物在开花后枯萎死亡,使除群落B以外的7个群落的AB重要值随时间推移下降(图3(a)、(c)~(h))。柳叶马鞭草在温暖地区为多年生植物,但在北京地区越冬困难通常作一二年生植物栽培,其花期偏晚(9月15日以后),且具有茎节伸长开花的特点,使其在9月中下旬开花时株高增长,从而增加了群落B在第1生长季末期的AB重要值(15.7%)(图3(b))。5个群落的IP重要值开始大于1%(图3(a)、(c)、(e)、(f)和(g)),本研究以重要值超过5%作为IP对群落结构产生影响的标志,其中群落A和C的IP重要值超过5%的时刻分别在第1生长季7月25日和8月15日。
第2生长季,AB在设计比例低于25%的4个群落之中不再出现(图3(b)、(e)、(f)和(g));尽管AB在设计比例大于40%的群落中出现, 但在生长季末期重要值均下降为0(图3(a)、(c)、(d)和(h))。这与各群落在第1—2生长季内总密度大幅下降的现象吻合(图2),并表明一二年生植物无法通过自播繁衍维持种群大小。AB消失后,各个群落开始由多年生植物主导,SP和LP重要值维持着此消彼长的关系。5个群落的IP重要值超过5%(图3(d)、(e)、(f)、(g)和(h)),开始对群落结构产生影响。
第3生长季,AB在所有群落中均不再出现,宿根亚麻和西洋滨菊等SP开始死亡,导致6个群落的SP年均重要值相比第2生长季下降13.0%~25.3%(图3(c)~(h)),但各群落SP和LP重要值仍维持此消彼长的关系。IP在群落B中首次出现,且重要值超过了5%(图3(b))。而在已经出现IP的群落中,IP在AB设计比例低于25%的3个群落中重要值持续低于8%(图3(e)~(g));在AB设计比例高于40%的4个群落中重要值分别上升至100%(图3(a))、33.7%(图3(c))、34.0%(图3(d))和15.0%(图3(h))。
第4生长季,SP在5个群落年均重要值相比第3生长季下降23.3%~62.2%(图3(b)~(e)和(h)),原因是大滨菊、菊花、穗花婆婆纳和蓝刺头等SP开始死亡。而群落F的SP平均重要值相比第3生长季不降反增了27.8%(图3(f)),是因为SP中寿命稍长的加拿大美女樱平均重要值大幅增加(增幅为136.8%),抵消了其他SP死亡带来的影响。
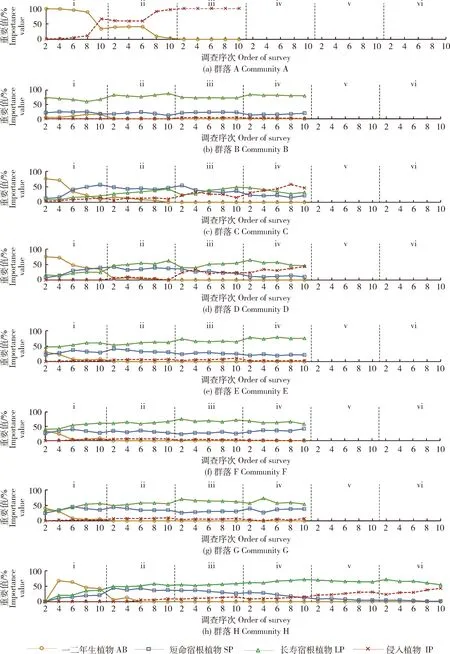
图3 群落A~H在各调查序次不同寿命组和侵入组植物的重要值Fig.3 Importance value (IV) of species composition in each longevity type (AB, SP & LP) and invasive plant (IP) within community A-H at each order of survey
随着SP死亡,LP开始占据优势。IP重要值在AB设计比例低于25%的群落中均持续低于5%(图3(b)、(e)和(f));在AB设计比例高于40%的群落中分别上升至58.0%(图3(c))、39.2%(图3(d))和15.0%(图3(h))。
在第5—6生长季,群落H的SP重要值持续下降,并在第6生长季末期重要值降为0(图3(h))。这表明SP也不能通过自播繁衍维持其种群大小。在第6生长季,IP重要值在9月25日达到了38.1% 的峰值,与LP维持着重要值此消彼长的关系。
以上结果表明,在混播群落构建过程中,AB的消亡过程从第1生长季中后期开始,至第2生长季末期结束。在AB设计比例为5%(图3(b))、15%~25%(图3(e)、(f)和(g))、40%~50%(图3(c)、(d)和(h))与100%(图3(a))的群落中,第2生长季与第1生长季年均AB重要值的差值分别为9.6%、18.6%±1.8%、46.4%±9.5%与64.9%;IP重要值超过5%的时刻分别为第3生长季、第2生长季、第2或第1生长季与第1生长季;最高IP重要值平均值分别为5.2%、6.7%±1.5%、37.2%±21.5%与100%。这些结果表明群落AB设计比例越高,其AB的消亡过程程度越剧烈,IP对群落产生影响的年份越靠前且IP重要值越容易上升。SP的消亡过程从第3生长季开始,持续至第6生长季。因SP的寿命存在个体差异,且其死亡后释放的群落空间资源由尚未死亡的SP、LP和IP共同竞争,使其重要值在第3生长季可能基本不变(图3(b))或在第3生长季下降后又在第4生长季小幅回升(图3(f))。
为进一步探究LP对IP的影响,对群落B~H的LP与IP在第2、第3和第4生长季的年均重要值做相关性分析,发现二者在3年间的年均重要值均极显著负相关(Pearson相关系数分别为-0.899(P<0.01)、-0.990(P<0.001)与-0.951(P<0.01)),表明持续存在的LP能够长时间抑制IP。而第2生长季的LP的平均重要值与LP设计比例极显著正相关(Pearson相关系数为0.970,P<0.001)。这表明增加LP的设计比例,有利于提高群落在第2生长季的LP重要值,提高群落在建植早期抑制侵入植物的能力。
2.3 农田野花带物种多样性指数的变化
相比第1生长季,混播群落第2生长季物种多样性指数有3个群落显著下降(图4(a)、(c)和(h)),4个群落下降但差异不显著(图4(b)、(d)、(f)和(g))以及1个群落小幅上升但差异不显著(图4(e))。多个群落物种多样性指数下降是因为大部分AB在第2年消失。群落E物种多样性小幅增加可能是因为群落物种均匀度增加的缘故。
相比第2生长季,第3生长季各群落物种多样性指数有1个群落显著下降(图4(g)),5个群落下降但差异不显著(图4(b)、(c)、(d)、(f)和(h))以及1个群落上升但差异不显著(图4(e))。所有AB种类在第3生长季春季消失,少部分SP种类在第3生长季夏季后死亡,导致群落物种多样性降低;但个体之间寿命存在差异,导致同一群落不同样方的物种多样性指数变异程度大,即使下降但仍未达显著水平。
相比第3生长季,第4生长季各群落物种多样性指数有4个群落下降但差异不显著(图4(b)、(d)、(e)和(f))以及2个群落小幅上升但差异不显著(图4(c)和(h))。多个群落物种多样性下降主要是因为SP种类持续死亡造成的,同样因个体寿命存在差异导致样方变异程度大,所以这些群落物种多样性指数下降仍未达显著水平。
相比第4生长季,第5生长季群落H物种多样性指数上升但差异不显著(图4(h)),这与群落H出现2种侵入植物(表1)有关,但其数量在不同样方中的异质性大。受SP种类持续死亡影响,第6生长季群落H物种多样性指数相比第5生长季下降但差异不显著。
6个群落(图4(a)、(c)、(d)、(f)、(g)和(h))有效最末观测生长季的物种多样性指数相比其第1生长季均显著下降,2个群落(图4(b)和(e))下降但差异不显著,表明群落构建过程中存在物种消亡,混播群落的物种多样性指数随时间推移呈下降趋势。
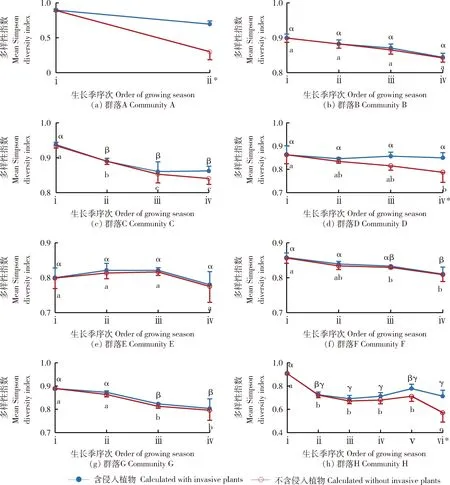
为避免重叠,剔除侵入植物以前的结果标准误差只显示正值;剔除侵入植物以后的结果标准误差只显示负值。剔除侵入植物之前和之后的数据Duncan多重比较结果(置信区间为95%)分别使用小写希腊字母和英文字母标注,字母相同表示二者之间不存在显著差异。*代表同一生长季中剔除侵入植物前后的Simpson多样性指数存在显著性差异(P<0.05)。For avoiding overlap, if invasive plants were eliminated in calculation, the result will only show positive standard error, otherwise the opposite. In each plot, use small Greek letter and small English letter to show Duncan’s Multiple Range Test of the two series of results independently. * means there is significant difference (P<0.05) between that two results in the same growing season.图4 群落A~H在各生长季序次剔除侵入植物前后的Simpson多样性指数Fig.4 Simpson diversity index calculated with and without invasive plants at each order of growing season in community A-H
为进一步探究IP对群落物种多样性指数的影响,剔除群落中的IP种类后重新计算了物种多样性指数(图4空心点)。发现剔除IP以后各群落的物种多样性指数总是不大于剔除IP以前的(图4实心点)。表明允许IP的存在不仅能补充群落密度,还能减缓群落物种多样性的丧失。IP在AB设计比例高于40%的群落中对于物种多样性指数的贡献量有逐年增加的趋势(图4(a)、(c)、(d)和(h)),且群落A在第2生长季,群落D在第4生长季以及群落H在第6生长季的物种多样性指数在剔除IP前后之间的差异达到了显著性水平(P<0.05)。但在前4个生长季中,AB设计比例低于25%的群落的物种多样性指数在剔除IP以后的降幅均不超过0.9%(图4(b)、(e)、(f)和(g)),表明IP对其影响甚微。
3 讨 论
3.1 混播建植的农田野花带群落结构变化规律
混播群落存在2次植物消亡过程。第1次消亡过程自一二年生植物在第1生长季开花后枯萎死亡开始,持续至第2生长季末;第2次消亡过程自短命宿根植物在第3生长季开花后枯萎死亡开始,持续至第6生长季末或更远。混播群落所使用的一二年生植物和短命宿根植物寿命到达年龄上限,且不能通过自播繁衍维持种群大小,是导致2次植物消亡过程的原因。以往的研究表明这是因花卉种子由于群落冠层的阻挡不能落入土壤种子库所致,而栽培品种结实能力低下或产出种子活力低也能造成这个结果[12,15]。
2次植物消亡过程为侵入植物的进入和扩张提供机会。在第1次植物消亡过程中,一二年生植物的迅速死亡会损失大量植株,这将使群落冠层打开“窗”,为侵入植物的种子进入群落和幼苗建植提供机会,释放的空间资源将由侵入植物和多年生植物共同竞争,这与天然林窗带来的群落自然更新过程类似[16]。一二年生植物的初始比例越大,第1次植物消亡过程越剧烈,群落冠层打开的“窗”越多,侵入植物的侵入机会和生长空间越大;反之,群落因一二年生植物死亡损失的植株少(如群落B),能够形成更郁闭冠层,侵入植物的侵入机会和生长空间小。短命宿根植物的消亡也为群落冠层打开“窗”,为侵入植物的进入提供机会,同时为已侵入的植物腾出生长空间。长寿宿根植物寿命长,其种群大小相对稳定,可通过无性繁殖占领2次植物消亡过程留下的群落空间,维持较高的重要值,并与侵入植物持续竞争。
3.2 构建稳定性高和功能持续的农田野花带混播群落的建议
2次植物消亡过程使混播群落的稳定性随时间推移降低。具体表现为群落密度、一二年生植物重要值、短命宿根植物重要值以及群落物种多样性指数随时间推移下降,同时侵入植物重要值随时间推移上升。尽管适度刈割能够维持或增加自然草甸的物种多样性[17-18],但在模拟人工建植的农田野花带“每年冬季刈割1次、低灌溉”的管理措施下,植物消亡过程发生在所有群落之中。保留侵入植物,能够减缓混播群落密度和物种多样性的丧失。兼顾农田野花带短期和长期的生态系统服务供给,根据混播群落结构变化规律和稳定性特征,从植物配置角度提出3个构建稳定性高和功能持续的农田野花带群落的建议。
1)提高长寿宿根植物比例。一二年生植物的设计比例越低,越能抑制侵入植物重要值的上升;且第1次植物消亡过程开始后,长寿宿根植物的重要值越高,群落抑制侵入植物能力越强。故可用快速开花的长寿宿根植物,如宿根鼠尾草、瓣蕊唐松草、宿根天人菊和紫色达利菊等[2,13]代替一二年生植物和短命宿根植物。快速开花的长寿宿根植物在第1~2年的花量较少,不能完全代替一二年生植物和短命宿根植物的功能,但可适当提高其设计比例,以提高长寿宿根植物的重要值。
2)适时人为扰动或补播。在植物结实后,人为干扰群落帮助种子落入土壤种子库,有助于一二年生植物和短命宿根植物自播繁衍。在一二年生植物和短命宿根植物花期后枯萎的同时对群落补播花卉种子,也能弥补群落密度和物种多样性的损失,并抢占杂草生存空间,抑制杂草侵入。
3)将自生植物融入农田野花带群落设计过程。在实际应用中难以清除土壤种子库,且非设计植物的侵入亦能补足群落因植物消亡产生的生态位空缺,增加群落密度和物种多样性。所以可采取“不对抗”的设计策略,事先调查本地土壤种子库,筛选其中生态和观赏价值较高的植物,在设计群落时为其预留生态位;或直接向农田周围自生植被中添加新植物,改善自生植被生态系统功能和景观表现[19]。这类设计策略有利于保护当地植物物种多样性[20]。
以上措施能帮助农田野花带长效且持续地发挥功能,为建设“美丽乡村”提供低投入、低维护和高效益的植物群落建植方法,从而给农村居民创造福祉,保护当地的生物多样性。
猜你喜欢
散文诗(2021年22期)2022-01-12
现代畜牧科技(2021年11期)2021-12-21
现代畜牧科技(2021年7期)2021-07-28
流行色(2020年3期)2021-01-04
花卉(2020年4期)2020-01-07
西藏农业科技(2019年1期)2019-07-25
新教育(2018年27期)2018-02-22
西南农业学报(2016年5期)2016-05-17
杂草学报(2015年2期)2016-01-04
现代农业(2015年5期)2015-02-28
