
不同森林经营模式对土壤氮含量及酶活性的影响
2021-07-28 04:07关庆伟王亚茹林鑫宇胡敬东
南京林业大学学报(自然科学版) 2021年4期
惠 昊,关庆伟王亚茹,林鑫宇,陈 斌,王 刚,胡 月,胡敬东
(南京林业大学,南方现代林业协同创新中心,南京林业大学生物与环境学院,江苏 南京 210037)
土壤是森林生态系统物质循环、能量流动的重要场所[1-2],为林木的生长提供养分,是森林生态系统营养元素转化的重要枢纽。存在于森林土壤中的氮元素是植物生长发育不可缺少的元素之一,也是限制森林生态系统生产力的重要因素[3-4],其含量特别是无机氮含量直接影响植物的吸收,同时也是造成环境污染的主要形式。
不同森林经营模式作为一种干扰,通过改变树种组成、调控林分密度等优化林分空间结构,提高生态系统功能。同时,也会改变植被条件和土壤温度、水分等因子,进而引起土壤氮和养分循环发生变化[5-7]。不同森林经营模式对土壤氮含量增加与否尚无一致结论,Wang等[8]研究发现,杉木桤木混交林和杉木刺楸混交林土壤全氮和微生物生物量氮含量均高于杉木纯林;Chen等[9]和Bolat[10]研究间伐后土壤微生物生物量氮含量显著增加,认为土壤温度和有机碳含量是控制土壤微生物生物量氮的主要因素;而Bai等[11]在澳大利亚的研究发现,间伐对土壤碳、氮含量无显著影响,可能是由供试土壤性质的差异所致。
目前,关于不同森林经营模式下土壤氮的动态变化研究主要以针叶林、阔叶林或针阔混交林等乔木为主,关于乔灌复层混交林对土壤氮含量的影响研究还鲜有报道。为此,本研究以杨树(Populuseuramericanacv. 35)纯林(PPP)、杨树女贞(Populuseuramericanacv. 35-Ligustrumlucidum)混交林(LMP)以及杨树石楠(Populuseuramericanacv. 35-Photiniaserrulata)混交林(PMP)3种林分为对象,探讨不同森林经营模式对林下土壤氮含量及相关酶活性的影响,为太湖沿岸防护林模式的构建提供科学参考。
1 材料与方法
1.1 试验地概况及样地设置
研究区域位于江苏省宜兴市周铁镇沙塘港村(120°01′E,31°25′N),距离太湖约1 km,地处亚热带季风气候区,夏季高温多雨,冬季低温少雨,平均气温15.7 ℃。降雨主要集中在夏季,且主要出现在梅雨和台风季节,6月和8月多暴雨,年平均降雨量为1 277.1 mm。土壤由太湖湖积物和长江冲积母质发育而成,为微酸性重壤质黄泥土,土质黏性重,腐殖质厚度2~3 cm,周边区域以农业为主,主要种植水稻、小麦、油菜等[12]。
供试试验地为2009年营造的杨树人工林,杨树品种为‘35杨’(Populuseuramericanacv.35),初始造林密度为720株/hm2,为进一步优化林分结构,提高生态系统功能,于2016年春季分别在杨树纯林下栽植了女贞和石楠两种灌木,形成了林龄相同的杨树纯林(PPP)、杨树女贞混交林(LMP)以及杨树石楠混交林(PMP)3种森林经营模式,林下植被以一枝黄花(Solidagodecurrens)、狼尾草(Pennisetumalopecuroides)、菟丝子(Cuscutachinensis)等为主,试验地林分基本概况见表1,土壤理化性质见表2。

表1 试验地林分基本概况

表2 试验地土壤理化性质
1.2 样品收集与指标测定
2019年3月,在每种模式下设置2块6 m×50 m的样地,随机选取4个小区作为4个重复。每个小区由随机选取的相邻4株杨树组成,分别在两株杨树之间1 m处以及距离杨树2.5 m处即4株杨树的中心,挖取1 m深的土壤剖面用于采集土壤样品,分0~20、≥20~40、≥40~60、≥60~80 cm 共4层采集,同一土层样品为两株杨树之间及距离杨树2.5 m处的混合土样。将新鲜土样置于保温箱中带回实验室,去除石块、植物根系等杂质,先在铝盒中称取少量鲜土置于烘箱烘干,测定土壤含水量。之后将土壤分成3份,其中1份保留较大土块自然风干,用于土壤团聚体的测定;其余土样过孔径为2 mm的钢筛,一部分置于4 ℃冰箱保存,用于测定土壤微生物生物量及不同形态氮,另一部分自然风干后粉碎磨细用于测定土壤理化性质及相关酶活性。另外,在每个样地内随机选取3个1 m×1 m的小样方,收集各样方内地表枯落物、草本地上部分,带回实验室测定其生物量。
各指标测定方法参照文献[13-14]:土壤全氮采用元素分析仪(Vario Element Ⅲ,Germany)测定;土壤硝态氮采用KCl浸提-双波长比色法测定;土壤铵态氮采用KCl浸提-靛酚蓝比色法测定;土壤微生物生物量氮采用氯仿熏蒸-K2SO4提取法测定;土壤脲酶活性采用靛酚蓝比色法测定;土壤硝酸还原酶活性和亚硝酸还原酶活性参照武志杰等[13]的方法进行测定。其余理化指标根据常规方法[14]进行测定,枯落物生物量和草本生物量采用烘干称质量法测定。
1.3 数据统计与分析
运用Excel 2016对试验数据进行整理,数据表示为平均值±标准误差(n=4)。利用SPSS 21.0进行统计分析,采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较不同森林经营模式之间和土层之间氮含量的差异,并用Origin 8.0 软件作图。
2 结果与分析
2.1 不同森林经营模式对土壤氮含量的影响
不同森林经营模式改变了土壤氮含量(图1),土壤全氮含量为0.17~1.35 g/kg,各森林经营模式之间差异显著(df=2,F=102.820,P<0.05)。与杨树纯林相比,杨树石楠混交林和杨树女贞混交林土壤全氮含量分别增加了21.8%和69.7%。3种经营模式下土壤全氮含量均随土层深度增加而降低,且各土层之间存在显著性差异(df=3,F=108.289,P<0.05)。
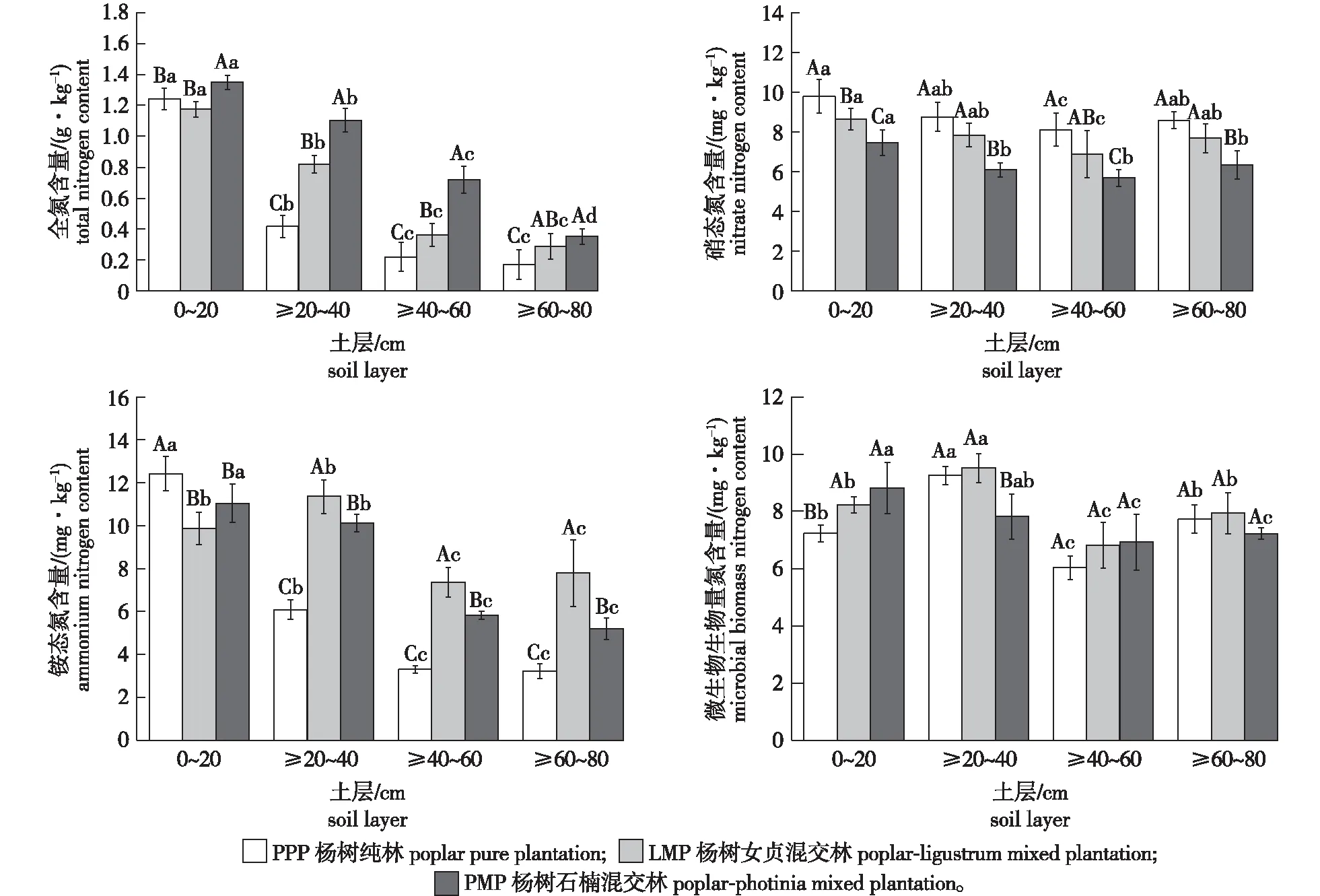
不同大写字母表示模式之间差异显著,不同小写字母表示土层之间差异显著。下同。Different capital letters indicate significant difference among the different forest management modes, and different lowercase letters indicate significant difference among the different soil layers. The same below.
从图1看出,土壤硝态氮含量为5.67~9.79 mg/kg,杨树纯林和杨树石楠混交林之间差异显著(df=2,F=18.764,P<0.05)。与杨树纯林相比,杨树女贞混交林和杨树石楠混交林土壤硝态氮含量分别降低了11.8%、27.3%;各森林经营模式土壤硝态氮含量均随土层深度增加而降低;土壤铵态氮含量为3.22~12.43 mg/kg,各森林经营模式之间存在显著性差异(df=2,F=9.655,P<0.05)。与杨树林相比,杨树女贞混交林和杨树石楠混交林土壤铵态氮含量在≥20~40、≥40~60和≥60~80 cm土层显著增加(df=3,F=106.230,P<0.05; df=3,F=119.794,P<0.05);此外,杨树纯林和杨树石楠混交林土壤铵态氮含量随土层深度增高而降低,杨树女贞混交林则先增高后降低。
土壤微生物生物量氮含量为6.04~9.52 mg/kg,各森林经营模式之间无显著性差异。与杨树纯林相比,杨树女贞混交林土壤微生物生物量氮含量增加了7.5%,杨树石楠混交林在0~20 cm和≥40~60 cm土层微生物生物量氮含量则分别增加了21.9%和14.8%;此外,杨树纯林和杨树女贞混交林土壤微生物生物量氮随土层深度增加先增高后降低,杨树石楠混交林则逐渐降低,且≥40 cm土层与<40 cm土层之间差异显著(df=3,F=4.554,P<0.05)。
2.2 不同森林经营模式对土壤氮循环相关酶活性的影响
土壤氮循环相关酶活性对森林经营模式的响应不一致(图2)。土壤脲酶活性为3.43~9.16 mg/(g·d),与杨树纯林相比,杨树女贞混交林土壤脲酶活性增强了18.2%,且在各土层差异显著(df=3,F=19.600,P<0.05);而杨树石楠混交林土壤脲酶活性在各土层均显著降低(df=3,F=10.915,P<0.05);此外,随着土层深度增加,杨树纯林土壤脲酶活性呈先增大后降低的趋势,杨树女贞混交林的除深层土壤外,总体呈增强趋势,杨树石楠混交林的则先降低后升高。
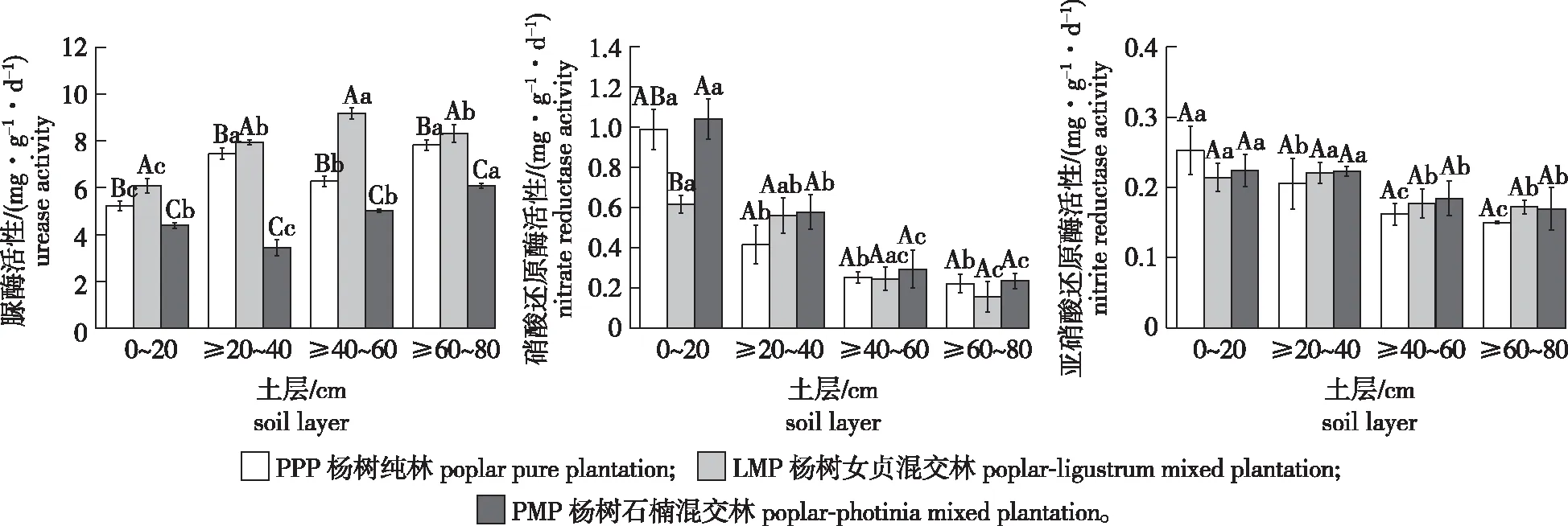
图2 不同森林经营模式对土壤氮循环相关酶活性的影响
土壤硝酸还原酶活性为0.16~1.04 mg/(g·d),与杨树纯林相比,杨树女贞混交林0~20 cm和≥60~80 cm土层土壤硝酸还原酶活性分别降低了37.7%和30.2%,≥20~40 cm土层硝酸还原酶活性则提高了34.5%;而杨树石楠混交林在各土层土壤硝酸还原酶活性均呈增强趋势。此外,随着土层深度的增加,3种森林经营模式土壤硝酸还原酶活性总体呈降低趋势,其表层土壤分别与≥40~60、≥60~80 cm土层之间存在显著性差异(df=3,F=17.825,P<0.05)。土壤亚硝酸还原酶活性为0.15~0.25 mg/(g·d),各森林经营模式之间土壤亚硝酸还原酶活性无显著性差异;随土层深度的增加,3种森林经营模式土壤亚硝酸还原酶活性总体呈降低趋势。
2.3 土壤理化性质与氮含量及相关酶活性之间的相关性分析
不同森林经营模式通过改变水热、pH等因子,影响土壤养分转化、相关酶活性等,是引起土壤氮含量发生变化的重要因素。相关性分析表明,土壤理化性质与氮含量及相关酶活性之间关系密切(表3)。土壤含水量与铵态氮、微生物生物量氮呈极显著正相关(df=3,F=164.759,P<0.01;df=3,F=37.241,P<0.01);土壤pH与全氮呈显著负相关(df=2,F=8.267,P<0.05),与铵态氮呈极显著负相关(df=3,F=287.421,P<0.01);脲酶与全氮呈极显著负相关(df=3,F=121.391,P<0.01),与铵态氮呈显著负相关(df=3,F=117.302,P<0.05);硝酸还原酶与全氮、硝态氮、铵态氮呈极显著正相关(df=3,F=198.349,P<0.01;df=2,F=23.731,P<0.01;df=3,F=101.092,P<0.01),与微生物生物量氮呈显著正相关(df=3,F=8.932,P<0.05);亚硝酸酶与铵态氮、微生物生物量氮呈极显著正相关(df=3,F=167.832,P<0.01;df=2,F=22.637,P<0.01)。表明土壤pH和含水量是引起土壤氮含量和相关酶活性发生变化的关键因素。

表3 土壤理化性质与氮含量及相关酶活性之间的相关性
3 讨 论
植物对土壤、化学和生物学过程有着重要的影响[15],其中地上凋落物是土壤有机养分供应者[16],与土壤全氮含量的关系密切。本研究中杨树石楠混交林和杨树女贞混交林的土壤全氮含量显著高于杨树纯林,这与王娟娟等[17]的结论一致。这是由于混交林林下凋落物量大于杨树纯林,凋落物组成结构和归还路径发生改变,促进了营养物质的供给和释放,提高了土壤养分和供氮能力,增加了氮源[18]。3种森林经营模式土壤全氮含量均随土层深度增加而降低。这是由于土壤养分具有表聚性,凋落物、细根等主要分布在表层土壤[19],参与有机质分解活动的微生物也集中在土壤表层;而随土层深度增加,土壤有机质来源减少,土壤水热、通气条件等亦不利于微生物活动,微生物分解和酶促反应受到抑制,土壤氮循环速率减弱,全氮含量逐渐降低。
土壤中硝态氮和铵态氮是可以被植物直接吸收利用的氮素[20],其中硝态氮为氧化态的氮源,易被植物吸收利用但易向土壤剖面深层移动;而铵态氮为还原态的氮源、移动性差,易被土壤吸附固定[21]。本研究中,除了土壤表层外,2种混交林的土壤铵态氮含量均高于杨树纯林,与土壤全氮含量变化规律总体一致。这是由于混交林下凋落物分解速率和氮素释放速率更快,促进了土壤氨化过程[22],土壤铵态氮含量高;同时,混交林偏酸性土壤对硝化作用的抑制以及黏土矿物和有机胶体对铵态氮的吸附作用也是导致混交林铵态氮含量偏高的原因[23]。然而,与杨树纯林相比,2种混交林下土壤硝态氮含量均显著降低,特别是深层土壤的硝酸盐含量,这与Zhu等[20]的结论一致。这一方面可能是由于女贞、石楠本身的生物学特性,且根系主要分布在土壤表层和亚表层,对硝态氮的吸收能力强[24-25];同时,硝态氮易向土壤剖面深层移动,杨树为深根系木本植物,可以吸收土壤剖面深层的硝酸盐[12,26];另一方面,混交林下凋落物分解和释放对土壤硝化过程影响较弱[22],大气来源的氮素[27]、土壤质地、水分和通气条件等也会影响硝化过程[28],抑制铵态氮向硝态氮的转化,导致土壤中累积的硝态氮含量偏低。
土壤微生物生物量氮作为土壤活性氮的来源, 对土壤氮素的供给起着重要的调节作用[29]。本研究中,杨树女贞混交林各土层的土壤微生物生物量氮含量最高,这与董敏慧等[30]的研究结果一致。这可能是由于混交林改善了林地的土壤质地和环境[31],凋落物归还量增加,土壤养分提高,促进了土壤微生物的繁殖;此外,土壤温度、水分、根系分泌物等也是引起土壤微生物生物量氮含量发生变化的重要因素[32]。虽然表层土壤具有较高的有机质含量和营养元素含量,但在本试验地植物细根主要聚集在土壤亚表层[25],根系分泌物相应增多,导致混交林微生物生物量氮含量在亚表层土壤达到最高;而深层土壤根系逐渐减少和营养物质匮乏,土壤水分、pH等因子受到限制,不利于微生物的生存,微生物生物量氮含量也随之减少。
土壤氮转化相关酶活性可以反映土壤氮素转化与供给状况[33],主要受土壤水分、温度、pH等因素影响[16,34]。本研究结果表明,杨树女贞混交林土壤脲酶活性高于杨树纯林,与彭春菊等[35]的研究结果一致。这可能是由于杨树女贞混交林凋落物分解、根系活动向土壤输入养分,增加了土壤微生物可利用的营养物质,使微生物数量增加、活性提高,从而导致脲酶活性增强[32]。此外,植物根系分泌物含量[36-37]和土壤颗粒组成的吸附作用[38],也可能会影响脲酶活性。杨树女贞混交林的除深层土壤外,总体呈增强趋势,杨树石楠混交林的则先降低后升高,与Tabatabai等[39]的研究结论不一致,这可能与土壤类型、植被类型和养分状况有关[40-41]。杨树女贞混交林相比杨树纯林,土壤表层和深层的硝酸还原酶活性降低,这可能是由于杨树女贞混交林地为偏酸性土壤(表2),通气条件差,硝化作用受到抑制[26],不利于微生物繁衍和植物根系活动。
综上所述,与杨树纯林相比,杨树女贞混交林和杨树石楠混交林在增加氮源的输入和提高土壤速效养分含量的同时,能有效吸收并削减土壤中硝态氮含量。这可能意味着混交林能够降低硝酸盐向土壤深层淋溶、污染浅层地下水的风险,建议今后在太湖流域多推广以乔灌复层混交林为主的森林经营模式。
参考文献(reference):
[1]FANG X M, ZHANG X F, CHEN F S, et al. Phosphorus addition alters the response of soil organic carbon decomposition to nitrogen deposition in a subtropical forest[J]. Soil Biology and Biochemistry, 2019, 133:119-128. DOI: 10.1016/j.soilbio.2019.03.005.
[2]LEENA F,TIMO D,SEID M D,et al. Conifer proportion explains fine root biomass more than tree species diversity and site factors in major European forest types[J]. Forest Ecology and Management, 2017, 406:330-350. DOI: 10.1016/j.foreco.2017.09.017.
[3]TILMAN D,CASSMAN K G,MATSON P A,et al.Agricultural sustainability and intensive production practices[J].Nature,2002,418(6898):671-677.DOI:10.1038/nature01014.
[4]ZHANG X, DAVIDSON E A, MAUZERALL D L, et al. Mana-ging nitrogen for sustainable development[J]. Nature, 2015, 528(7580):51-59. DOI: 10.1038/nature15743.
[5]VALVERDE-BARRANTES O J, ROCHA O J. Logging impacts on forest structure and seedling dynamics in aPrioriacopaifera(Fabaceae) dominated tropical rain forest (Talamanca, Costa Rica)[J]. Revista De Biologia Tropical,2014,62(1):347-357. DOI:10.15517/rbt.v62i1.8504.
[6]RUEL J C, FORTIN D, POTHER D. Partial cutting in old-growth boreal stands: an integrated experiment[J]. The Forestry Chronicle, 2013, 89(3): 360-369. DOI: 10.5558/tfc2013-066.
[7]周焘,王传宽,周正虎,等.抚育间伐对长白落叶松人工林土壤碳、氮及其组分的影响[J].应用生态学报,2019,30(5):1651-1658.ZHOU T,WANG C K,ZHOU Z H,et al.Effects of thinning on soil carbon and nitrogen fractions in aLarixolgensisplantation[J].Chin J Appl Ecol,2019,30(5):1651-1658.DOI:10.13287/j.1001-9332.201905.020.
[8]WANG Q K,WANG S L.Soil microbial properties and nutrients in pure and mixed Chinese fir plantations[J].Journal of Forestry Research,2008,19(2):131-135.DOI:10.1007/s11676-008-0022-7.
[9]CHEN X L,WANG D,CHEN X,et al.Soil microbial functional diversity and biomass as affected by different thinning intensities in a Chinese fir plantation[J]. Appl Soil Ecol, 2015, 92: 35-44.DOI:10.1016/j.apsoil.2015.01.018.
[10]BOLAT I.The effect of thinning on microbial biomass C,N and basal respiration in black pine forest soils in Mudurnu,Turkey[J].European Journal of Forest Research,2014,133(1):131-139.DOI:10.1007/s10342-013-0752-8.
[11]BAI S H, DEMPSEY R, REVERCHON F, et al. Effects of forest thinning on soil-plant carbon and nitrogen dynamics[J]. Plant and Soil, 2017, 411(1/2):437-449. DOI: 10.1007/s11104-016-3052-5.
[12]吴殿鸣,薛建辉,罗英,等.杨麦间作系统硝态氮损失的原位研究[J].南京林业大学学报(自然科学版), 2012,36(2):111-116. WU D M, XUE J H, LUO Y, et al. Study on nitrate nitrogen leaching in poplar-wheat intercropping ecosystem byinsituanalysis[J]. J Nanjing For Univ (Nat Sci Ed), 2012, 36(2):111-116. DOI:10.3969/j.issn.1000-2006.2012.02.023.
[13]武志杰,隽英华,陈利军,等.一种检测土壤硝酸还原酶活性的分析方法:CN101271060A[P].2008-09-24. WU Z J, JIU Y H, CHEN L J, et al. A modified method for measuring soil nitrate reductase activity: CN101271060A[P].2008-09-24.
[14]鲁如坤. 土壤农业化学分析方法[M].北京: 中国农业科技出版社, 2000.LU R K. Soil argrochemistry analysis protocols[M]. Beijing: China Agriculture Science Press. 2000.
[15]杨万勤, 王开运. 森林土壤酶的研究进展[J]. 林业科学, 2004, 40(2):152-159. YANG W Q, WANG K Y. Advances in Forest Soil Enzymology[J]. Sci Silvae Sin, 2004, 40(2):152-159. DOI:10.11707/j.1001-7488.2004.02.027.
[16]鲍勇, 高颖, 曾晓敏, 等. 中亚热带3种典型森林土壤碳氮含量和酶活性的关系[J]. 植物生态学报,2018, 42(4)508-516. BAO Y, GAO Y, ZENG X M, et al. Relationships between carbon and nitrogen contents and enzyme activities in soil of three typical subtropical forests in China[J]. Chin J of Plant Ecol, 2018, 42(4):508-516. DOI:10.17521/cjpe.2017.0311.
[17]王娟娟, 朱莎, 靳士科, 等. 上海市3种森林类型土壤微生物生物量碳和氮的时空格局[J]. 生态与农村环境学报, 2016, 32(4):615-621. WANG J J,ZHU S,JIN S K, et al. Spatio-temporal distribution of soil microbial biomass carbon and nitrogen in three types of urban forest soils in Shanghai[J]. J of Ecol and Rural Environ, 2016, 32(4):615-621. DOI: 10.11934/j.issn.1673-4831.2016.04.016.
[18]李瑞霞. 间伐对马尾松细根生物量、形态和碳氮含量的影响[D]. 南京: 南京林业大学, 2014. LI R X. Effects of thinning on biomass, morphology and carbon and nitrogen content of fine roots ofPinusmassoniana[D]. Nanjing: Nanjing Forestry University, 2014.
[19]张志铭, 赵河, 杨建涛, 等. 太行山南麓山区不同植被恢复类型土壤理化和细根结构特征[J]. 生态学报, 2018, 38(23):8363-8370. ZHNAG Z M, ZHAO H, YANG J T, et al. Soil physicochemical properties and fine root characteristics of different restored vegetation types in southern Taihang Mountain[J]. Acta Ecol Sin, 2018, 38(23):8363-8370.DOI: 10.5846 /stxb201801190150.
[20]ZHU F F, DAI L M, ERIK A, et al. Uptake patterns of glycine, ammonium, and nitrate differ among four common tree species of northeast China[J]. Frontiers in Plant Science, 2019, 10:799. DOI: 10.3389/fpls.2019.00799.
[21]詹媛媛,薛梓瑜,任伟,等.干旱荒漠区不同灌木根际与非根际土壤氮素的含量特征[J]. 生态学报,2009,29(1): 59-66. ZHAN Y Y,XUE Z Y,REN W,et al.Characteristics of nitrogen content between rhizopshere and bulk soil under seven shrubs in arid desert area of China[J]. Acta Ecol Sin, 2009, 29(1): 59-66. DOI: 10.3321 /j.issn:1000-0933.2009.01.008.
[22]冯烨, 张焕朝, 杨瑞珍, 等. 杨桤混交林及其凋落物对土壤氮矿化的影响[J].南京林业大学学报(自然科学版), 2019, 43. FENG H, ZHANG H C, YANG R Z, et al. The influence of poplar-alder mixed forest and litter to soil nitrogen mineralization[J]. J Nanjing For Univ(Nat Sci Ed), 2019,43. DOI:10.3969/j.issn.1000-2006.201902025.
[23]叶钰倩, 赵家豪, 刘畅, 等. 间伐对马尾松人工林根际土壤氮含量及酶活性的影响[J]. 南京林业大学学报(自然科学版),2018, 42(3):193-198. YE Y Q, ZHAO J H, LIU C, et al. Effects of thinning on nitrogen contents and enzyme activities of rhizosphere soil inPinusmassonianaplantations[J]. J Nanjing For Univ(Nat Sci Ed), 2018, 42(3):193-198. DOI:10.3969/j.issn.1000-2006.201709026.
[24]王卫国, 童根平, 徐温新, 等. 3种常见速生灌木吸收红壤氮磷能力的比较[J]. 浙江林学院学报, 2009, 26(3):346-350. WANG W G, TONG G P, XU W X, et al. Absorption of soil N and P by three fast growing bush species[J]. J Zhejiang For Coll, 2009, 26(3):346-350. DOI:10.3969/j.issn.2095-0756.2009.03.009.
[25]芦建国, 梁同江, 孔凡海. 8种灌木根系分布对高速公路生态边坡的影响[J]. 南京林业大学学报(自然科学版), 2011, 35(5):155-159. LU J G, LIANG T J, KONG F H. Influence of eight species shrubs’ roots distribution on the ecological slope of highway[J]. J Nanjing For Univ(Nat Sci Ed),2011, 35(5):155-159.DOI: 10.3969/j.issn.1000-2006.2011.05.035.
[26]薛建辉, 阮宏华, 刘金根, 等. 太湖流域水岸生态防护林体系建设技术与对策[J]. 南京林业大学学报(自然科学版), 2008, 32(5):13-18.XUE J H, RUAN H H, LIU J G, et al. The construction techniques and strategies of riparian shelterbelt systems surrounding Taihu Lake watershed[J]. J Nanjing For Univ(Nat Sci Ed),2008, 32(5):13-18. DOI: 10.3969/j.issn.1000-2006.2008.05.004.
[27]周石磊,孙悦,黄廷林,等. 周村水库大气湿沉降氮磷及溶解性有机物特征[J]. 水资源保护,2020,36(3):52-59. ZHOU S L, SUN Y, HUANG T L, et al. Characteristics of nitrogen, phosphorus and dissolved organic matter in atmospheric wet deposition of Zhoucun Reservoir[J]. Water Resour Prot,2020,36(3):52-59. DOI:10.3880/j.issn.1004-6933.2020.03.010.
[28]朱兆良, 文启孝. 中国土壤氮素[M]. 南京:江苏科学技术出版社,1992. ZHU Z L, WEN Q X. Nitrogen in soil of China[M]. Nanjing: Jiangsu Science and Technology Press, 1992.
[29]XU X, THORNTON P E, POST W M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems[J]. Global Ecology and Biogeography, 2013, 22(6):737-749. DOI: 10.1111/geb.12029.
[30]董敏慧, 张良成, 文丽, 等. 松树-樟树混交林、纯林土壤微生物量碳、氮及多样性特征研究[J]. 中南林业科技大学学报, 2017, 37(11):146-153. DONG H M,ZHANG L C,WEN L,et al. Soil microbial biomass C, N and diversity characteristics in pure and mixed forest ofPinusandCinnamomun[J]. J Central South Univ For Technol, 2017,37(11):146-153. DOI: 10.14067/j.cnki.1673-923x.2017.11.024.
[31]田大伦, 项文化, 闫文德. 马尾松与湿地松人工林生物量动态及养分循环特征[J]. 生态学报, 2004, 24(10):2207-2210. TIAN D L,XIANG W H,YAN W D,et al.Biomass dynamics and nutrient cycling characteristics ofPinusmassonianaandPinuselliottiiplantations[J]. Acta Ecol Sin, 2004,24(10):2207-2210. DOI:10.3321/j.issn:1000-0933.2004.10.017.
[32]胡亚林, 汪思龙, 颜绍馗, 等. 杉木人工林取代天然次生阔叶林对土壤生物活性的影响[J]. 应用生态学报, 2005, 16(8):1411-1416. HU Y L, WANG S L, YAN S K, et al. Effect of Chinese fir plantation replacing natural secondary broad-leaved forest on soil biological activity[J]. Chin J Appl Ecol, 2005,16(8):1411-1416. DOI: 10.13287/j.1001-9332.2005.0167.
[33]孙瑞莲, 赵秉强, 朱鲁生, 等. 长期定位施肥对土壤酶活性的影响及其调控土壤肥力的作用[J]. 植物营养与肥料学报, 2003, 9(4):406-410. SUN R L, ZHAO B Q, ZHU L S, et al. Effects of long-term fertilization on soil enzyme activity and its regulation of soil fertility[J]. Plant Nutr Fertil Sci, 2003,9(4):406-410. DOI:10.3321/j.issn:1008-505X.2003.04.005.
[34]刘捷豹, 陈光水, 郭剑芬,等. 森林土壤酶对环境变化的响应研究进展[J]. 生态学报, 2017, 37(1):110-117. LIU J B, CHEN G S, GUO J F, et al. Advances in research on the responses of forest soil enzymes to environmental change[J]. Acta Ecol Sin, 2017, 37(1):110-117. DOI: 10.5846/stxb201608011581.
[35]彭春菊, 李全, 顾鸿昊, 等. 模拟氮沉降及经营方式对毛竹林土壤酶活性的影响[J]. 应用生态学报, 2017, 28(2):423-429. PENG C J, LI Q, GU H H, et al. Effects of simulated nitrogen deposition and management type on soil enzyme activities in Moso bamboo forest[J]. Chin J Appl Ecol, 2017,28(2):423-429.DOI:10.13287/j.1001-9332.201702.001.
[36]万欣,李文斌,黄海涛,等. 扬州茱萸湾风景区森林土壤质量状况[J]. 江苏林业科技,2020,47(2):40-43,47.WAN X, LI W B, HUANG H T, et al. Soil quality of urban forest in different vegetationtypes in Zhuyuwan Scenic Spot, Yangzhou [J].Journal of Jiangsu Forestry Science & Technology,2020,47(2):40-43,47. DOI: 10.3969/j.issn.1001-7380.2020.02.008.
[37]杨万勤, 宋光煜, 韩玉萍. 土壤生态学的理论体系及其研究领域[J]. 生态学杂志, 2000, 19(4):53-56. YANG W Q, SONG G Y, HAN Y P. Theoretical system and research fields of soil ecology[J]. Chin J Ecol, 2000, 19(4):53-56. DOI: 10.13292/j.1000-4890.2000.0058.
[38]周礼恺.土壤的酶活性[J]. 土壤学进展, 1980, (4):9-15. ZHOU L K. Enzyme activity of soil[J]. Progress in Soil Science, 1980, (4):9-15.
[39]TABATABAI M A, DICK W A. Distribution and stability of pyrophosphatase in soils[J]. Soil Biol and Biochem, 1979, 11(6):655-659. DOI: 10.1016/0038-0717(79)90035-x.
[41]孟立君, 吴凤芝. 土壤酶研究进展[J]. 东北农业大学学报, 2004, 35(5):622-626.MENG L J, WU F Z, et al. Research progress of soil enzymes[J]. J Northeast Agric Univ, 2004, 35(5):622-626. DOI:10.3969/j.issn.1005-9369.2004.05.024.
猜你喜欢
读者(2023年2期)2023-02-20
花卉(2022年11期)2022-06-09
故事会(2022年10期)2022-05-20
知识窗(2019年7期)2019-08-12
浙江农业科学(2019年4期)2019-04-19
森林工程(2018年4期)2018-08-04
基层中医药(2018年1期)2018-03-01
方圆(2018年23期)2018-01-07
现代园艺(2017年11期)2017-06-28
上海农业学报(2016年2期)2016-10-27
