
长白落叶松-水曲柳混交林不同混交方式单木冠长预测模型
2021-07-28 04:07:28贺梦莹董利虎李凤日
南京林业大学学报(自然科学版) 2021年4期
贺梦莹,董利虎,李凤日*
(1. 东北林业大学林学院,森林生态系统可持续经营教育部重点实验室,黑龙江 哈尔滨 150040; 2. 吉林省林业调查规划院,吉林 长春 130022)
长白落叶松(Larixolgensis)和水曲柳(Fraxinusmandshurica)是我国东北地区主要用材树种,长白落叶松-水曲柳混交林是我国东北地区较为成功的混交林类型。树冠作为树木的重要组成部分,其大小对树木的生长活力有重要的作用,树冠大小可以用来预测树木活力和木材质量[1]。冠长是研究树冠大小和树冠结构的重要因子之一,因此构建准确的冠长模型具有重要的意义。
国内外学者对冠长率进行了一定的研究,但对冠长的研究不多。许多学者通常采用Logistic函数、指数函数、Richards函数、Weibull函数、对数函数等构建冠长率模型[2-5]。Soares等[4]利用4种非线性模型构建了单木冠长率模型,研究结果表明冠长率与树木年龄和林分密度有关,竞争对树木冠长率具有一定的影响。Sattler等[6]利用三步最小二乘法构建了复杂林分中的冠长和树冠半径的联立方程组模型。计算冠长通常需要获得树高和枝下高的数据,国内外学者主要采用简单或者复杂的模型来构建树高曲线模型,树高曲线是指树高与胸径之间的相关曲线[7]。一般的树高曲线仅以胸径为自变量构建,对于不同林分的树高预测不够准确,因此在树高曲线中加入林分变量和竞争因子等不仅可以提高树高曲线的预测精度,还可以使其适用范围更广[8-9]。冠长与树高之间的相关性可以采用非线性联立方程组解决,不仅可以确保冠长与树高之间的约束关系,还可以解释冠长和树高之间的内在相关性[10-11]。以往有学者研究采用似乎不相关回归的方法构建枝下高和树高的联立方程组,不仅保证各方程误差的协方差在没有限制误差的情况下具有渐进有效性,促使变量的方差逐渐减小,而且考虑了枝下高与树高之间的高度内在相关性[12-14]。
植物间的竞争会导致个体对竞争的可塑生长及异速生长关系的改变[15]。张彦东等[16]研究表明,在带状幼龄混交林中,靠近落叶松带边行水曲柳的树高生长和胸径生长优于中间行水曲柳,靠近水曲柳带的落叶松树高生长和胸径生长优于中间行的落叶松。在人工混交林内,不同混交比例林分内树木的种内竞争和种间竞争强度不同,影响树木发育,导致树木冠长和树高不同。因此,构建冠长模型和树高曲线模型时考虑树木在混交带的位置和混交林的混交比例非常重要。
为了揭示不同混交方式长白落叶松-水曲柳混交林林木冠长的生长规律,本研究以不同林分条件的长白落叶松-水曲柳混交林为研究对象,分别构建了不同混交方式下长白落叶松和水曲柳的冠长模型和树高曲线模型,在最优冠长模型和最优树高曲线模型的基础上,采用非线性似乎不相关回归(NSUR)的方法构建冠长模型和树高曲线模型的联立方程组,并对所构建的模型进行评价。
1 材料与方法
1.1 研究地概况
研究地位于黑龙江省尚志国有林场管理局小九林场(127°38′27″~127°51′37″E,45°11′29″~45°25′05″N)、一面坡林场(127°59′09″~128°18′43″E,44°52′43″~45°11′54″N)、帽儿山林场(127°18′00″~127°41′06″E,45°02′20″~45°18′16″N)、黑龙宫林场(127°37′45″~127°56′12″E,44°23′55″~45°35′08″N)和元宝林场(128°10′17″~128°33′07″E,45°08′26″~45°18′26″N)。各林场海拔为200~600 m,年平均气温在2.3 ℃左右,年平均降水量600~800 mm。
1.2数据来源
2017—2019年在黑龙江省尚志国有林场管理局小九林场、一面坡林场、帽儿山林场、黑龙宫林场和元宝林场不同林分条件的长白落叶松-水曲柳混交林内设置54块标准地。标准地形状为矩形,大小根据不同混交配置而定,要求覆盖3个重复的混交林带,长白落叶松与水曲柳行间混交比例分别为1∶1、2∶2、3∶3、5∶5,标准地大小为0.06~0.32 hm2。设置标准地后,对每块标准地内的样木按顺序进行逐一标号,并对样木进行定位,记录定位坐标;对每块标准地进行每木检尺,测定和记录标准地内各标号样木的树木因子,包括胸径、树高以及枝下高等。冠长由树高与枝下高的差计算所得。
本研究共收集54块标准地的6 118株长白落叶松样木和9 286株水曲柳样木数据,将标准地随机抽样按照2∶1的比例分别划分成拟合数据(36块标准地的长白落叶松样木4 108株、水曲柳样木6 195 株)和检验数据(18块标准地的长白落叶松样木2 010株、水曲柳样木3 091株),分别用于模型拟合和模型检验。长白落叶松-水曲柳混交林林分因子及样木因子特征详见表1和表2。

表2 长白落叶松和水曲柳样木因子特征

表2(续)
1.3 基础模型


表3 冠长基础模型
2)树高曲线基础模型。以往针对树高曲线的研究,大多数是以胸径为自变量,再将其他林分变量和竞争因子加到模型中,通常采用幂函数模型、单分子式模型、理查德模型、逻辑斯蒂模型等构建[9,17-21]。本研究选取单分子式模型作为构建长白落叶松和水曲柳树高曲线的基础模型,基础模型形式如下:
H=a1[1-exp(a2DBH)]。
(1)
1.4 模型影响因子及联立方程组
1)混交比例和树木在混交带中的位置。 长白落叶松与水曲柳混交林的混交比例分别为1∶1、2∶2、3∶3、5∶5。在混交林中,根据长白落叶松距水曲柳带的距离可分为边行长白落叶松、中间行长白落叶松;边行长白落叶松两侧相邻行均为水曲柳或者一侧相邻行为水曲柳,中间行长白落叶松两侧相邻行均为长白落叶松。同样水曲柳可分为边行水曲柳、中间行水曲柳。
由于不同混交方式林分的树木冠长和树高大小具有差异,为了更精确预测混交林内长白落叶松和水曲柳的冠长及树高,将树木在混交带内位置K(边行、中间行)和混交比例Zi(长白落叶松与水曲柳的混交比例分别为1∶1、2∶2、3∶3、5∶5)作为哑变量引入模型中,建立效果更好的哑变量模型。其中:K=1时为边行树木,K=0时为中间行树木;则Z1=1、Z2=0、Z3=0、Z4=0时混交比例为1∶1,Z1=0、Z2=1、Z3=0、Z4=0时混交比例为2∶2,Z1=0、Z2=0、Z3=1、Z4=0时混交比例为3∶3,Z1=0、Z2=0、Z3=0、Z4=1时混交比例为5∶5。
2)与距离无关的竞争因子。 考虑外业成本及实际应用,本研究选择与距离无关的竞争因子来量化林木竞争状态,分析竞争对长白落叶松和水曲柳冠长的影响。竞争因子包括:大于对象木的胸高断面积之和、林木树高与林分优势木平均高之比,以及林木胸径与林分优势木平均胸径之比[22-23]:
HDH=H/H0;
(2)
DDH=DBH/D0;
(3)
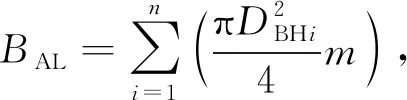

(4)
式中:HDH为林木树高与林分优势木平均高之比;DDH为林木胸径与林分优势木平均胸径之比;BAL为大于对象木的胸高断面积之和;H0为优势木平均高;D0为优势木平均胸径;DBHi为第i株竞争木的胸径;n为株数;DBH0为对象木胸径。
3)其他影响因子。 树木变量为树高、高径比和冠长率;林分变量为林分密度、林分年龄、优势木平均胸径、优势木平均高、林分平均胸径、林分平均树高、长白落叶松平均树高、水曲柳平均树高、长白落叶松优势木平均胸径、水曲柳优势木平均胸径、长白落叶松优势木平均高、水曲柳优势木平均高等。
考虑模型稳定性及拟合效果,避免过度参数化和变量间的共线性问题,并考虑选取的变量在应用中地面调查获取数据的难易程度及变量在模型中的意义,采用全子集回归法进行自变量的筛选,选取拟合优度和检验结果最优的冠长和树高曲线的哑变量模型。模型的构建及变量的选取通过SAS 9.4软件实现。
为了考虑冠长与树高之间的内在相关性问题,本研究采用长白落叶松和水曲柳的冠长模型与树高曲线模型建立联立方程组,利用非线性似乎不相关回归(NSUR)方法进行参数估计,模型基本形式如下:
(5)
式中:fCL为冠长函数;fH为树高函数;x为协变量;β为模型参数;ε为模型误差项。
以上联立方程组用SAS/ETS模块的SUR进行拟合。
1.5 模型评价与检验
(6)
(7)
σ(AIC)=2p+nln(SSR/n);
(8)
(9)
(10)
(11)
2 结果与分析
2.1 基础模型选定
2.2 哑变量模型构建
由于在混交林中边行、中间行和不同混交比例林分的长白落叶松、水曲柳冠长大小具有差异,因此将树木的混交带内位置K(边行、中间行)和混交比例Zi(长白落叶松与水曲柳混交比例分别为1∶1、2∶2、3∶3、5∶5)作为哑变量引入长白落叶松和水曲柳冠长基础模型中,通过比较不同的参数组合哑变量模型的收敛情况和拟合效果,本研究选择将rHD、HDH加到长白落叶松冠长模型中,将rHD和DDH加到水曲柳冠长模型中,加入哑变量和竞争因子的长白落叶松和水曲柳的最优冠长模型形式如下:
LLar=H/{1+exp[-(bl1DBH+bl2rHD+bl3HDH+sl1Z1+sl2Z2+sl3Z3+sl4Z4+plK)]}+εl;
(12)
LFra=H/{1+exp[-(bf1DBH+bf2rHD+bf3DDH+sf1Z1+sf2Z2+sf3Z3+sf4Z4+pfK)]}+εf。
(13)
式中:LLar为长白落叶松冠长;LFra为水曲柳冠长;sl1、sl2、sl3、sl4、sf1、sf2、sf3、sf4、bl1、bl2、bl3、bf1、bf2、bf3、pl、pf为模型参数;εl、εf为模型误差。
由于在混交林中边行、中间行和不同混交比例林分的长白落叶松、水曲柳的树高大小具有差异,因此将树木在混交带内位置K和混交比例Zi作为哑变量引入长白落叶松和水曲柳树高曲线基础模型中,通过比较不同的参数组合哑变量模型的收敛情况和拟合效果,本研究选择将HgLar加到长白落叶松树高曲线模型中,将H0Fra加到水曲柳树高曲线模型中,加入哑变量和竞争因子的长白落叶松和水曲柳的树高曲线最优哑变量模型形式如下:
HLar=(bl1+bl3HgLar)[1-exp(-bl2DBH+sl1Z1+sl2Z2+sl3Z3+sl4Z4+plK)]+εl;
(14)
HFra=bf1[1-exp(-bf2DBH+bf3H0Fra+sf1Z1+sf2Z2+sf3Z3+sf4Z4+pfK)]+εf。
(15)
根据模型(12)—(15),利用非线性最小二乘法分别计算长白落叶松和水曲柳冠长模型的参数估计值,如表4所示。所有模型参数估计值的标准差均较小,说明所引入变量均差异显著,模型参数估计值比较稳定。
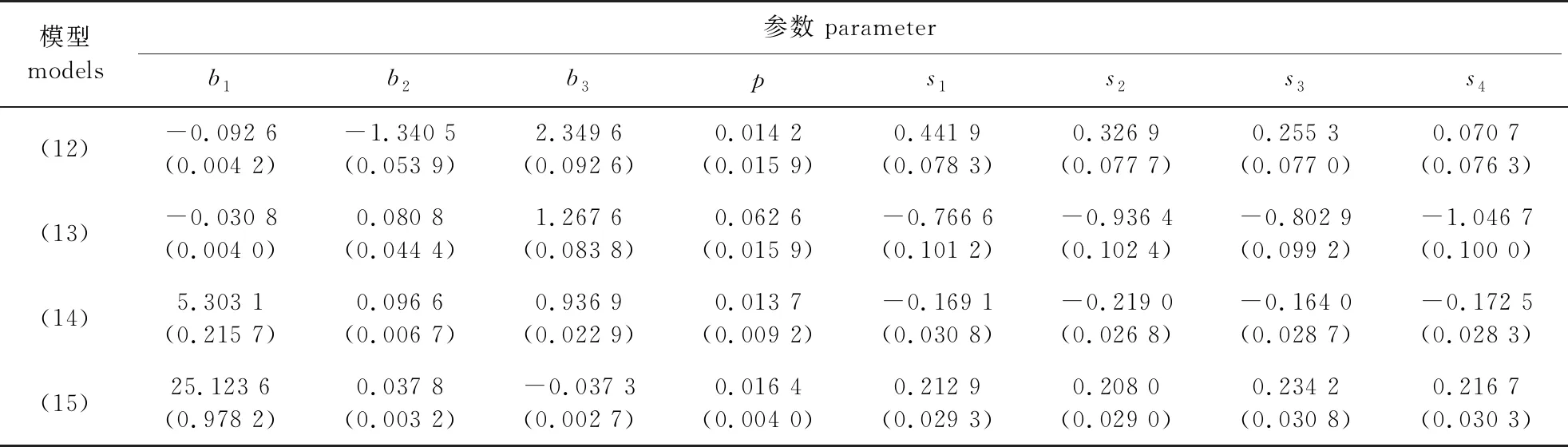
表4 长白落叶松和水曲柳冠长、树高曲线模型的参数估计结果
长白落叶松和水曲柳冠长模型、树高曲线模型的拟合优度及检验结果见表5。

表5 长白落叶松和水曲柳冠长、树高曲线模型的拟合优度及检验结果

2.3 联立方程组及预测结果
由于单独构建冠长模型不能解释冠长与树高之间的内在相关性,所以利用以上构建的长白落叶松、水曲柳的最优冠长模型和最优树高曲线模型建立联立方程组,采用NSUR方法估计模型参数,模型形式如下:
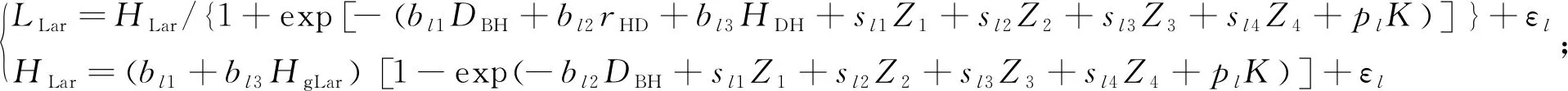
(16)

(17)
长白落叶松冠长模型与树高曲线模型联立方程组(16)和水曲柳冠长模型与树高曲线模型联立方程组(17)的参数估计值如表6所示,所有模型参数估计值的标准差均较小,说明所引入变量均差异显著,模型参数估计值比较稳定。

表6 联立方程组的参数估计结果
长白落叶松冠长模型与树高曲线模型联立方程组(16)和水曲柳冠长模型与树高曲线模型联立方程组(17)的拟合优度及检验结果如表7所示。
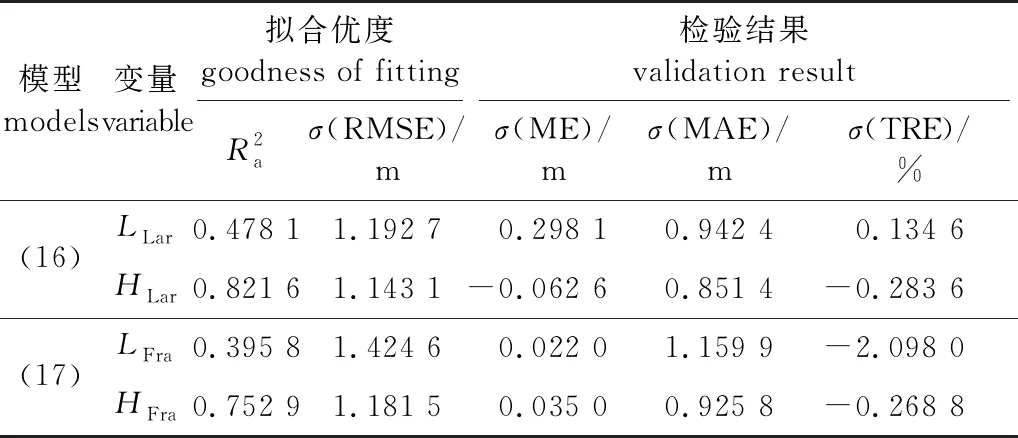
表7 联立方程组(16)和(17)的拟合优度及检验结果
基于检验数据,根据所构建的冠长和树高联立方程组(16)和(17)分别绘制出长白落叶松和水曲柳的冠长、树高的残差分布图,如图1所示,残差的散点是随机分布的,没有明显的趋势,说明模型的预测效果较好。

图1 长白落叶松和水曲柳的冠长及树高的预估值及残差分布
基于所构建的长白落叶松和水曲柳的冠长模型、树高曲线联立方程组,以混交比例为5∶5林分内的中间行树木为例,绘制不同条件下冠长与胸径的关系曲线。如图2所示,长白落叶松冠长随着高径比的增大而减小,随着HDH的增大而增大;水曲柳冠长随着高径比的增大而减小,随着DDH的增大而增大。
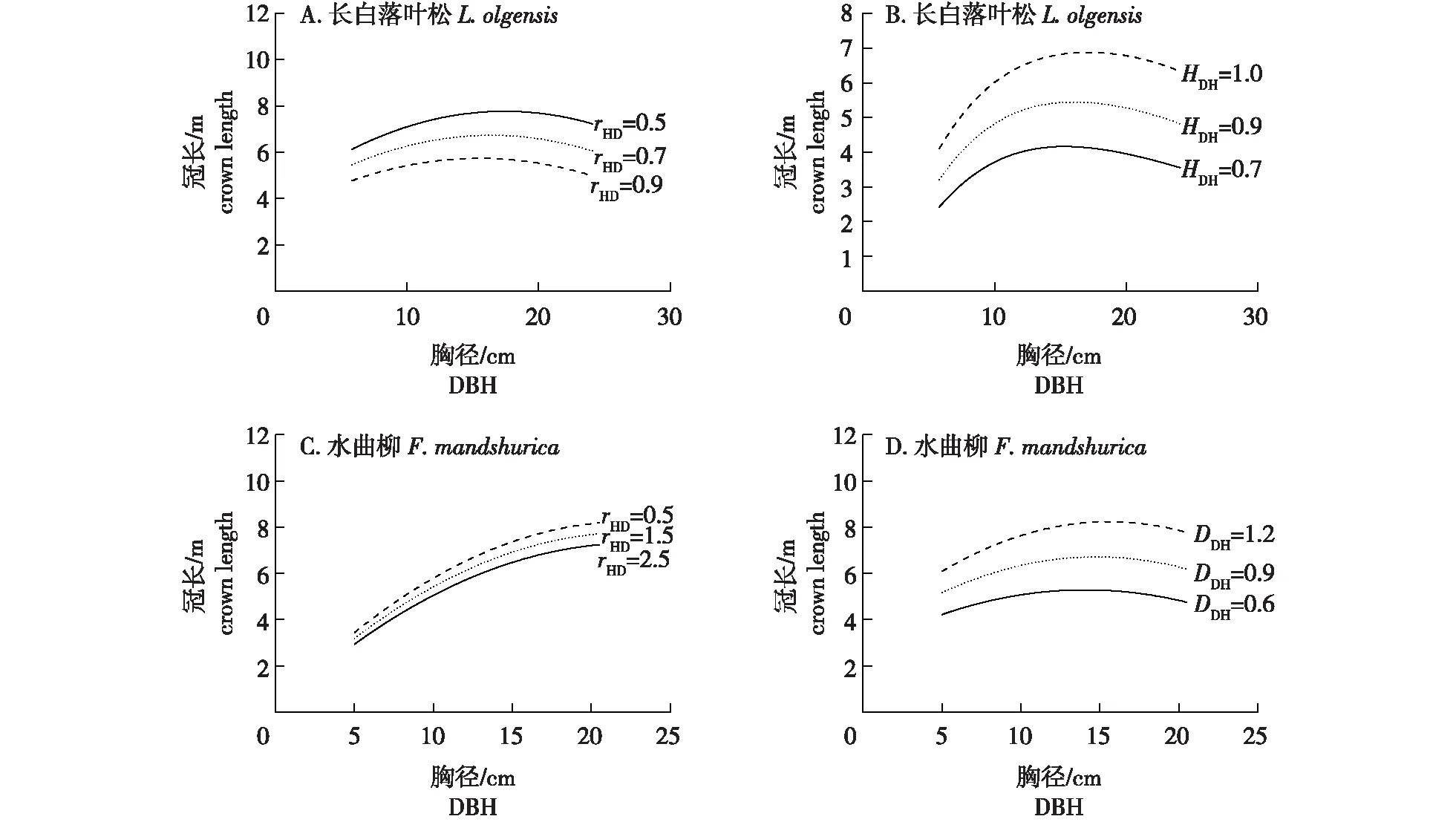
图2 不同条件下长白落叶松和水曲柳的冠长与胸径的关系曲线
基于所构建的长白落叶松和水曲柳的冠长模型、树高曲线模型联立方程组,分别绘制4种混交比例林分的长白落叶松和水曲柳冠长与胸径的关系曲线见图3。如图3所示,长白落叶松在混交比例为 1∶1的林分内冠长最大,在混交比例5∶5的林分内冠长最小;水曲柳同样在混交比例为1∶1的林分内冠长最大,在混交比例5∶5的林分内冠长最小。总体上来讲,由于立地条件和林分条件的影响,林分内种间竞争的强度与种内竞争的强度具有差异,导致不同混交比例林分内的林木冠长具有差异。
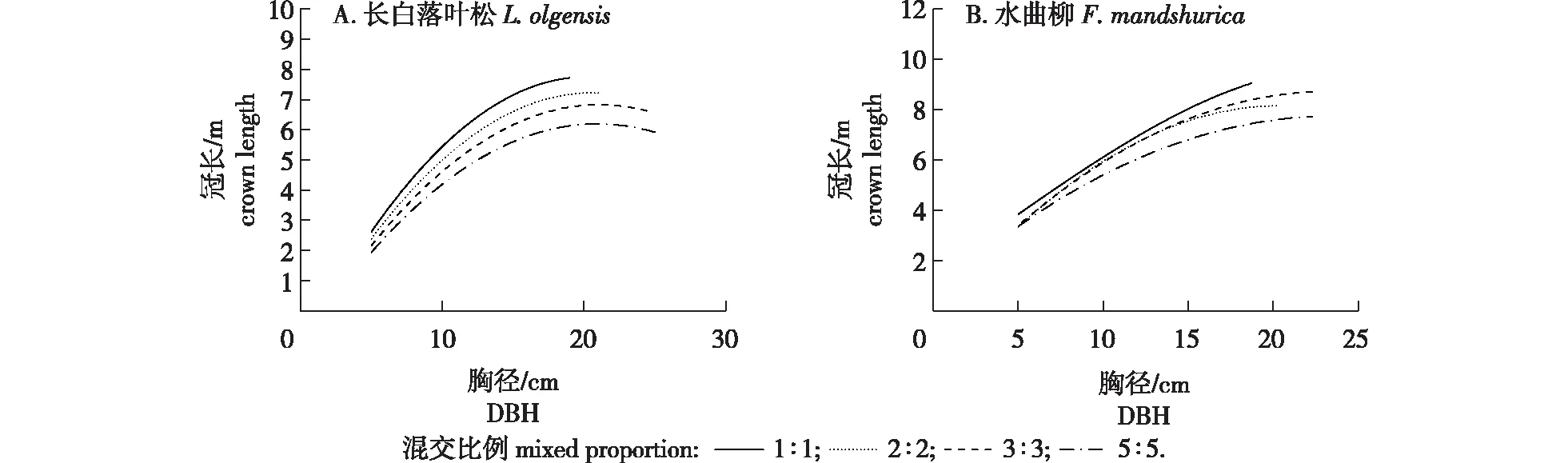
图3 长白落叶松和水曲柳在不同混交比例林分下冠长与胸径的关系曲线
3 讨 论
本研究在长白落叶松冠长模型中加入了高径比(rHD)和林木树高与林分优势木平均高之比(HDH),冠长与rHD为负相关关系,与HDH为正相关关系;在水曲柳冠长模型中加入了rHD和林木胸径与林分优势木平均胸径之比(DDH),冠长与rHD为负相关关系,与DDH为正相关关系。rHD与林分密度密切相关,是反映树木和林分稳定性的重要指标[21]。rHD越大,树木越容易受到外部条件的破坏,因此rHD越小,树冠越大,而且与针叶树相比,rHD对阔叶树种的稳定性影响较小[21-22]。rHD也反映了树木的竞争情况,竞争压力越大,rHD越大,导致树冠衰退,冠长越小[23]。树木的HDH、DDH越大,说明树木的生长活力越好,在竞争中处于更有利的地位,所以树冠发育越好,冠长越大,同时也说明与距离无关的竞争因子可以很好地反映树木受到的竞争压力,具有代表性[24-25]。在长白落叶松和水曲柳的树高曲线方程中分别加入了林分变量长白落叶松平均树高(HgLar)和水曲柳优势木平均高(H0Fra),林分平均树高和优势木平均树高是反映立地条件和树木生长状况的重要指标,林分平均树高和优势木平均树高越大,说明立地条件越好,越有利于树木的生长发育,树木越高。基于构建的长白落叶松和水曲柳冠长模型、树高曲线联立方程组,分别模拟了不同混交比例林分下的两树种冠长的变化规律,结果表明,由于立地条件、林分条件以及竞争因素的影响,导致不同混交比例林分的长白落叶松和水曲柳的冠长具有差异。
本研究利用不同混交方式林分内的长白落叶松-水曲柳混交林54块标准地的6 118株长白落叶松样木和9 286株水曲柳样木的数据,分别构建了两个树种的冠长模型和树高曲线的联立方程组,模型具有较好的预测能力。本研究中,长白落叶松样木最大胸径为25.10 cm,水曲柳样木最大胸径为25.30 cm,缺乏大径阶数据,在预估大径阶(DBH>30 cm)的冠长时,可能产生预测误差。由于树冠大小受立地条件、林分条件等因素的影响,随着更多数据的收集,今后可进一步研究不同林分条件下的混交林单木冠幅和冠长的联立方程模型。
参考文献(reference):
[1]VALENTINE H T,LUDLOW A R,FURNIVAL G M.Modeling crown rise in even-aged stands of Sitka spruce or Loblolly pine[J].For Ecol Manag,1994,69(1/2/3):189-197.DOI:10.1016/0378-1127(94)90228-3.
[2]MONSERUD R A,STERBA H.A basal area increment model for individual trees growing in even- and uneven-aged forest stands in Austria[J].For Ecol Manag,1996,80(1/2/3):57-80.DOI:10.1016/0378-1127(95)03638-5.
[3]卢军,李凤日,张会儒,等.帽儿山天然次生林主要树种冠长率模型[J].林业科学,2011,47(6):70-76.LU J,LI F R,ZHANG H R,et al.A crown ratio model for dominant species in secondary forests in Mao’er Mountain[J].Sci Silvae Sin,2011,47(6):70-76.
[4]SOARES P,TOMÉ M.A tree crown ratio prediction equation forEucalyptplantations[J].Ann For Sci,2001,58(2):193-202.DOI:10.1051/forest:2001118.
[5]POPOOLA F S,ADESOYE P O.Crown ratio models forTectonagrandis(Linn.f) stands in Osho forest reserve,Oyo State,Nigeria[J].J For Environ Sci,2012,28(2):63-67.DOI:10.7747/jfs.2012.28.2.063.
[6]SATTLER D F,LEMAY V.A system of nonlinear simultaneous equations for crown length and crown radius for the forest dynamics model SORTIE-ND[J].Can J For Res,2011,41(8):1567-1576.DOI:10.1139/x11-078.
[7]胥辉,全宏波,王斌.思茅松标准树高曲线的研究[J].西南林学院学报,2000,20(2):74-77.XU H,QUAN H B,WANG B.A study on the model of theoretical height-diameter curve ofPinuskesiyavar.langbianensis[J].J Southwest For Coll,2000,20(2):74-77.DOI:10.3969/j.issn.2095-1914.2000.02.003.
[9]张鹏,王新杰,许昊.将乐地区马尾松标准树高曲线的研究[J].中南林业科技大学学报,2015,35(3):69-73.ZHANG P,WANG X J,XU H.Generalized height-diameter model ofPinusmassonianafor Jiangle State-owned Forest Farm,Fujian Province[J].J Central South Univ For Technol,2015,35(3):69-73.DOI:10.14067/j.cnki.1673-923x.2015.03.014.
[10]唐守正,李勇,符利勇.生物数学模型的统计学基础[M].2版.北京:高等教育出版社,2015.TANG S Z,LI Y,FU L Y.Statistical foundation for biomathematical models[M].2nd ed. Beijing:Higher Education Press,2015.
[11]TANG S Z,LI Y,WANG Y H.Simultaneous equations,error-in-variable models,and model integration in systems ecology[J].Ecol Model,2001,142(3):285-294.DOI:10.1016/S0304-3800(01)00326-X.
[12]李想,董利虎,李凤日.基于联立方程组的人工樟子松枝下高模型构建[J].北京林业大学学报,2018,40(6):9-18.LI X,DONG L H,LI F R.Building height to crown base models for Mongolian pine plantation based on simultaneous equations in Heilongjiang Province of northeastern China[J].J Beijing For Univ,2018,40(6):9-18.DOI:10.13332/j.1000-1522.20170428.
[13]李想.黑龙江省人工樟子松枝下高动态预测模型的研究[D].哈尔滨:东北林业大学,2018.LI X.The study of dynamic predicting model of height to crown base for Mongolian pine plantation in Heilongjiang Province[D].Harbin:Northeast Forestry University,2018.
[14]DONG L H,ZHANG L J,LI F R.A three-step proportional weighting system of nonlinear biomass equations[J].For Sci,2015,61(1):35-45.DOI:10.5849/forsci.13-193.
[15]安慧,上官周平.密度对刺槐幼苗生物量及异速生长模式的影响[J].林业科学,2008,44(3):151-155.AN H,SHANGGUAN Z P.Effects of density on biomass and allometric pattern ofRobiniapseudoacaciaseedling[J].Sci Silvae Sin,2008,44(3):151-155.DOI:10.3321/j.issn:1001-7488.2008.03.028.
[16]张彦东,王庆成,谷艳华.水曲柳落叶松人工幼龄混交林生长与种间竞争关系[J].东北林业大学学报,1999,27(2):6-9.ZHANG Y D,WANG Q C,GU Y H.The relationship between growth and interspecific competition within the ash-larch mixed stand[J].J Northeast For Univ,1999,27(2):6-9.DOI:10.3969/j.issn.1000-5382.1999.02.002.
[17]陈丽聪,邓华锋,黄国胜,等.不同起源马尾松与杉木林分树高曲线的拟合及对比[J].西北农林科技大学学报(自然科学版),2014,42(1):57-64.CHEN L C,DENG H F,HUANG G S,et al.Fitting and comparison of tree height curves ofPinusmassonianaLamb andCunninghamialanceolatawith different origins[J].J Northwest A F Univ (Nat Sci Ed),2014,42(1):57-64.DOI:10.13207/j.cnki.jnwafu.2014.01.006.
[18]赵俊卉,亢新刚,刘燕.长白山主要针叶树种最优树高曲线研究[J].北京林业大学学报,2009,31(4):13-18.ZHAO J H,KANG X G,LIU Y.Optimal height-diameter models for dominant coniferous species in Changbai Mountain,northeastern China[J].J Beijing For Univ,2009,31(4):13-18.DOI:10.3321/j.issn:1000-1522.2009.04.003.
[19]卢军,张会儒,雷相东,等.长白山云冷杉针阔混交林幼树树高:胸径模型[J].北京林业大学学报,2015,37(11):10-25.LU J,ZHANG H R,LEI X D,et al.Height-diameter models for saplings in a spruce-fir mixed forest in Changbai Mountains[J].J Beijing For Univ,2015,37(11):10-25.DOI:10.13332/j.1000-1522.20140429.
[21]朱万才,吴瑶,李亚洲,等. 不同保留密度对落叶松水曲柳混交林的影响[J]. 森林工程,2021,37(2): 50-56.ZHU W C,WU Y,LI Y Z,et al. Effect of different reserve density onLarixgmelinii-Fraxinusmandshuricamixed forest[J]. Forest Enginee-ring,2021,37(2): 50-56.
[22]VOSPERNIK S,MONSERUD R A,STERBA H.Do individual-tree growth models correctly represent height:diameter ratios of Norway spruce and Scots pine?[J].For Ecol Manage,2010,260(10):1735-1753.DOI:10.1016/j.foreco.2010.07.055.
[23]SHARMA R P,VACEK Z,VACEK S.Individual tree crown width models for Norway spruce and European beech in Czech Republic[J].For Ecol Manag,2016,366:208-220.DOI:10.1016/j.foreco.2016.01.040.
[24]BIGING G S,DOBBERTIN M.Evaluation of competition indices in individual tree growth models[J].For Sci,1995,41(2):360-377.DOI:10.1093/forestscience/41.2.360.
[25]赵俊卉.长白山云冷杉混交林生长模型的研究[D].北京:北京林业大学,2010.ZHAO J H.Growth modeling for spruce-fir fo-rest in Changbai Mountains[D].Beijing:Beijing Forestry University,2010.
猜你喜欢
现代交际(2022年10期)2022-11-01 08:06:02
当代音乐(2022年8期)2022-04-29 16:02:48
林业科技情报(2021年1期)2021-04-14 09:10:02
当代音乐(2019年11期)2019-11-11 02:23:50
山东林业科技(2019年2期)2019-06-03 10:10:54
农民致富之友(2019年11期)2019-05-23 01:14:52
森林工程(2018年4期)2018-08-04 03:23:10
农民致富之友(2017年23期)2018-01-02 02:02:06
河北经贸大学学报(2017年2期)2017-02-15 22:38:35
管理现代化(2016年6期)2016-01-23 02:10:52
