
胁迫萌发对青稞籽粒中β-葡聚糖和γ-氨基丁酸含量的影响
2021-07-28 06:46李婷玉陈正行周文菊涂兆鑫
中国粮油学报 2021年6期
李婷玉 杜 艳 陈正行 周文菊 涂兆鑫 李 娟
(粮食发酵工艺与技术国家工程实验室;江南大学1,无锡 214122)(青海华实科技投资管理有限公司2,西宁 810016)(江苏省生物活性制品加工工程技术研究中心;江南大学3,无锡 214122)
青稞营养丰富,蛋白质含量适中,且脂肪含量较低,并含有丰富的矿质元素、维生素、膳食纤维以及β-葡聚糖、γ-氨基丁酸(GABA)和α-生育三烯醇等多种生理功效成分,其生态特性和营养保健作用都是其它粮食作物无法比拟的。其中,青稞中的β-葡聚糖平均含量为5.25%,是目前世界上β-葡聚糖含量最高的麦类作物。β-葡聚糖具有调节血糖、降低胆固醇、提高免疫力等功能[1]。GABA具有降血压、神经营养功能、抗癫痫病、生殖生理功能等多种生理功能[2]。
发芽可用于改善谷物的外观、质地、风味、口感及其营养价值。然而,发芽通常导致谷物中β-葡聚糖的分解,β-葡聚糖被内源性的β-葡聚糖酶水解。金属离子对谷物萌发过程中β-葡聚糖的含量和β-葡聚糖酶的活性有很大影响。钟正升[3]研究发现Mg2+、Zn2+、K+、Na+有助于β-葡聚糖酶的生成及β-葡聚糖的分解; Cu2+会抑制β-葡聚糖酶活性及β-葡聚糖的降解。毕静[4]的研究结果表明Cu2+和Fe2+对酶活有明显的抑制作用。彭维[5]的研究表明Na+、K+、Mg2+、Mn2+、Fe3+、Ca2+对β-葡聚糖酶具有激活作用,而Cu2+对其起抑制作用。富硒发芽可以抑制发芽青稞中β-葡聚糖的降解[6]。萌发可诱导生物活性成分的变化,尤其可以积累GABA等营养物质。研究表明,当植物暴露于许多不利条件时,如缺氧、干旱、低温、高温、高盐等,GABA的积累迅速增加,表明GABA在非生物胁迫中可能起主要作用[7]。低温胁迫[8-11]是促进GABA积累的有效途径。Wallace等[12]研究发现,低温胁迫通过改变植物细胞结构,降低pH,激活谷氨酸脱羧酶(GAD)活力从而促进GABA生成,同时抑制GABA转氨作用,使GABA得到积累,但低温胁迫萌发对β-葡聚糖含量的影响研究较少。
本实验以青稞籽粒为原料,在萌发过程中采用金属离子、亚硒酸钠和低温胁迫处理的方式,研究不同胁迫方式下青稞籽粒中主要功能性成分β-葡聚糖和GABA含量的变化情况和规律,以期为功能性青稞产品的开发提供参考。
1 材料与方法
1.1 原料与试剂
青稞(瓦蓝,2019年9月收获);七水合硫酸镁、硫酸锌、五水合硫酸铜、六水合氯化亚铁、十二水合氯化铁、亚硒酸钠、四氢呋喃、三乙胺、盐酸、结晶乙酸钠、四硼酸钠、2-巯基乙醇、三氯乙酸(TCA)、冰醋酸、氢氧化钠、无水乙醇,均为分析纯;次氯酸钠(有效氯6%~14%)、邻苯二甲醛(OPA)、γ-氨基丁酸标准品(色谱级)、Megazyme β-葡聚糖混联检测试剂盒、甲醇(色谱纯)、乙腈(色谱纯)。
1.2 仪器设备
BSC-250型恒温恒湿箱, GZX-9246 MBE型数显鼓风干燥箱, 150型高速多功能粉碎机, LXJ-IIB型低速大容量多管离心机, UV-3200型紫外分光光度计, KQ-50E型超声波清洗器, NexIon 350D型电感耦合等离子体质谱仪, Agilent 1260 series 型高效液相色谱仪, Agilent Hypersil ODS柱(4.0 mm×250 mm,5 μm)。
1.3 方法
1.3.1 发芽青稞的制备
选取饱满的青稞籽粒,用体积分数为0.2%~1%次氯酸钠溶液浸泡消毒30 min后,去离子水清洗3~4次,再用去离子水在25 ℃下浸泡10 h。随后,在直9 cm 的培养皿底部铺四层纱布,然后均匀铺放200粒青稞籽粒,在上面盖2 层湿纱布。在温度为15 ℃、相对湿度95%、避光的条件下发芽培养,每隔6 h洒100 mL去离子水。分别于发芽24、48、72 h时取样,用去离子水洗涤3次后,用80 ℃热水灭酶处理10 min,40 ℃干燥12 h,粉碎(过50目筛)。
1.3.2 金属离子胁迫萌发
发芽48 h,在培养过程中喷洒不同浓度(0.04、0.08、0.16 g/L)金属离子溶液(Fe2+、Fe3+、Cu2+、Mg2+、Zn2+)[5]。其余具体胁迫萌发方法同1.3.1。
1.3.3 亚硒酸钠胁迫萌发
发芽48 h,在培养时喷洒不同浓度(4、8、16 mg/L)的亚硒酸钠的溶液[6]。其余具体胁迫萌发方法同1.3.1。
1.3.4 低温胁迫萌发
将洗净浸泡后的青稞籽粒分别在5 ℃和-20 ℃胁迫24、48、72 h取出并在室温下回温30 min后萌发24 h。其余具体胁迫萌发方法同1.3.1。
1.3.5 β-葡聚糖含量测定
按照NY/T 2006—2011《谷物及其制品中β-葡聚糖含量的测定》。
1.3.6 GABA含量测定
采用OPA柱前衍生反相高效液相色谱-紫外检测法。
供试品溶液的制备:精确称取1.0 g青稞粉,加入5%三氯乙酸适量,摇匀,并定容至25 mL,常温超声40 min,离心(4 000 r/min,10 min)。取上清液,过0.22 μm滤膜。
色谱条件:色谱柱:Agilent Hypersil ODS柱(4.0 mm×250 mm,5 μm)。流动相A(pH=7.2)为27.6 mmol/L 乙酸钠-三乙胺-四氢呋喃(体积比为500∶0.11∶2.5),流动相B(pH=7.2)为80.9 mmol/L 乙酸钠-甲醇-乙腈(体积比为1∶2∶2),流速为1.0 mL/min,柱温为40 ℃。检测波长338 nm。采用梯度洗脱,洗脱程序为:0 min,8% B;17 min,50% B;20.1 min,100% B;24.0 min,0% B。
衍生条件:用高效液相色谱系统自动衍生装置,将200 μL 供试品溶液和400 μL衍生剂[13]反应2 min。
1.3.7 金属离子残留量
按照GB 5009.268—2016《食品中多元素的测定》。
1.3.8 数据分析
所有实验均重复2次,结果以平均值±标准差表示。应用SPSS软件进行方差分析,显著性分析采用Ducan’s多重检验,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果与讨论
2.1 发芽对青稞β-葡聚糖和GABA含量的影响
由图1可知,随着胁迫萌发时间延长,青稞中β-葡聚糖含量呈现下降的趋势。这可能是因为青稞在发芽时,胚上皮层细胞分泌了赤霉酸,刺激胚乳糊粉层合成β-葡聚糖酶,水解胚乳细胞的β-葡聚糖[14],从而使得β-葡聚糖含量减少。其中,β-葡聚糖含量在发芽48~72 h时降解速率最快。这可能是因为发芽早期,β-葡聚糖酶活性较小,β-葡聚糖降解较慢。随着发芽时间延长,β-葡聚糖酶活性增加,进而加快了β-葡聚糖降解。在整个发芽过程中,青稞中GABA含量总体呈先增加后减少的趋势。GABA在发芽前期(0~24 h)积累较慢,到了发芽中期(24~48 h)GABA积累最快,而到了发芽后期(48~72 h) GABA含量减少。这是因为青稞籽粒在发芽初期,蛋白酶未被充分激活,随着发芽时间延长,蛋白质水解形成了谷氨酸,在谷氨酸脱羧酶的作用下使得谷氨酸转化为GABA[15]。而GABA的生成与消耗是一个动态平衡的过程[16],随着GABA的积累,反遏抑制了GAD酶的活性而促进了GABA转氨酶的活性,使GABA转化为琥珀酸半醛,从而导致GABA含量的下降[17]。
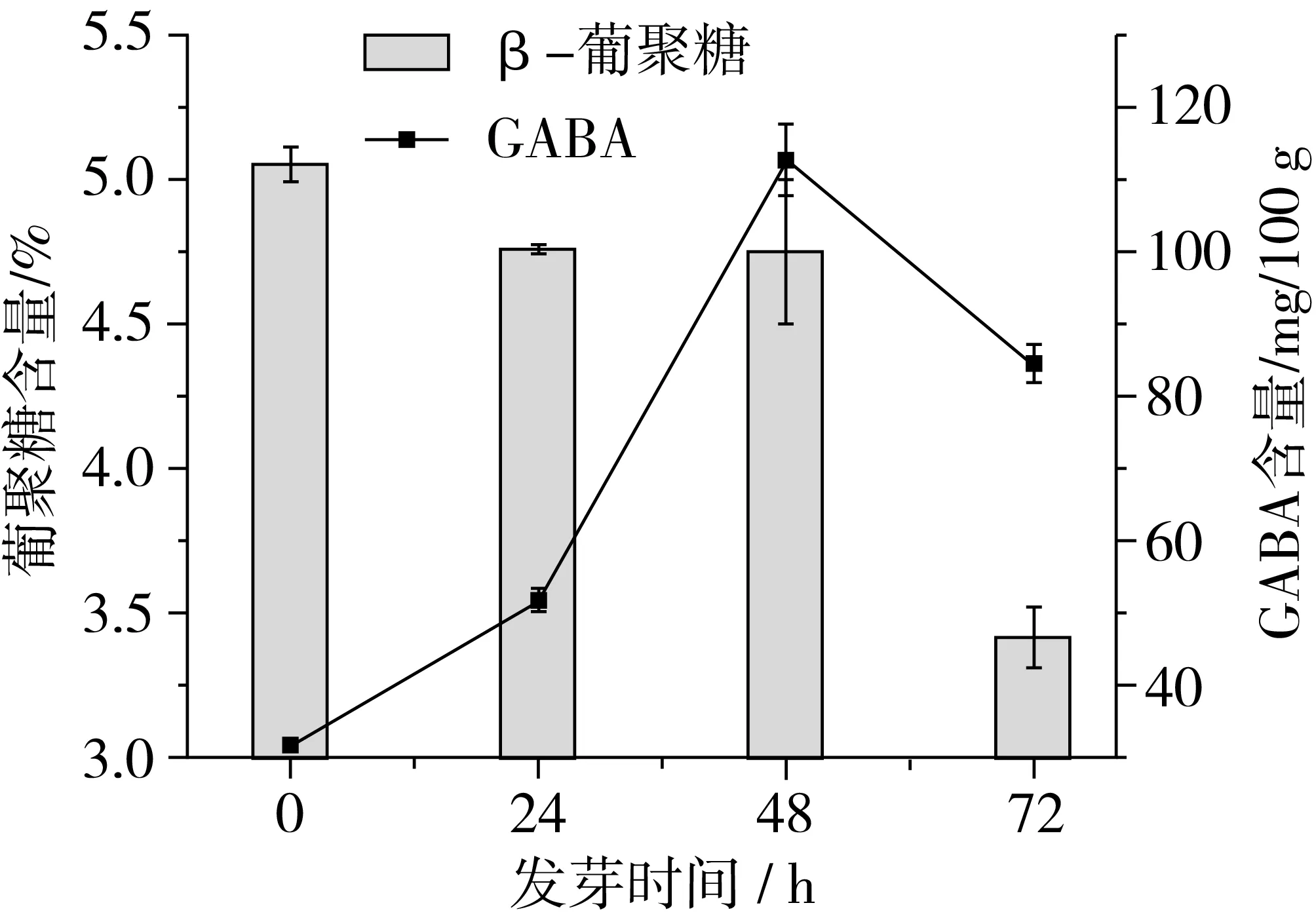
图1 发芽对青稞β-葡聚糖和GABA含量的影响
2.2 金属离子胁迫萌发对青稞β-葡聚糖和GABA含量的影响
由表1可知,采用5种金属离子胁迫萌发48 h β-葡聚糖含量低于对照组,明显可以促进β-葡聚糖降解,可能是因为Fe2+、Fe3+、Cu2+、Mg2+、Zn2+可以促进β-葡聚糖酶的活性[5-7]。而且采用5种金属离子胁迫青稞萌发48 h 后GABA含量均低于对照组,表明Fe2+、Fe3+、Cu2+、Mg2+、Zn2+胁迫萌发抑制了青稞中GABA的积累。在5种金属离子中,用Zn2+胁迫萌发48 h GABA含量增加量最少,最不利于GABA富集。关于Fe2+、Fe3+、Cu2+、Mg2+、Zn2+胁迫萌发对青稞籽粒 GABA富集效果的研究还鲜见报道,可能是因为金属毒害导致青稞萌发能力丧失[17]。
用不同浓度的Fe2+、Fe3+、Cu2+、Mg2+、Zn2+溶液胁迫萌发青稞种子会造成金属离子进入种子,形成富集,而这些离子摄入过多对人体造成一定的伤害。因此本实验对经不同浓度(0.04、0.08、0.16 g/L)金属离子溶液(Fe2+、Fe3+、Cu2+、Mg2+、Zn2+)胁迫48 h的发芽青稞中的Fe、Cu、Mg、Zn残留量进行了检测。经测定,用Fe2+、Fe3+、Cu2+、Mg2+、Zn2+胁迫发芽48 h的发芽青稞中各元素的残留量如表2所示,Cu、Zn残留量低于NY 861—2004 粮食(含谷物、豆类、薯类)及制品中铅、镉、铬、汞、硒、砷、铜、锌等8种元素限量中规定的上限(10.50 mg/kg)。
2.3 亚硒酸钠胁迫萌发对青稞β-葡聚糖和GABA含量的影响
Marco等[19]研究发现,亚硒酸钠会影响小麦籽粒的发育,减缓胚根的生长,一定浓度的亚硒酸钠溶液会抑制植物籽粒的生长,进而减缓了β-葡聚糖的降解速率。由图2可知,4 mg/L的亚硒酸钠溶液胁迫下的发芽青稞中β-葡聚糖含量高于对照组,8、16 mg/L亚硒酸钠溶液胁迫下的发芽青稞中β-葡聚糖含量明显低于对照组,表明亚硒酸钠溶液浓度较低时可以抑制发芽青稞中β-葡聚糖的降解,亚硒酸钠溶液浓度较高时可以促进发芽青稞中β-葡聚糖的降解。4、8、16 mg/L亚硒酸钠溶液作用下的发芽青稞GABA含量低于对照组,表明亚硒酸钠不利于GABA积累。可能是由于一定浓度的亚硒酸钠溶液会抑制植物籽粒的生长。
用不同浓度的亚硒酸钠溶液胁迫萌发青稞种子,会造成硒进入种子,形成富集,而这些元素摄入过多对人体造成一定的伤害。因此本实验对经不同浓度(4、8、16 mg/L)亚硒酸钠溶液胁迫48 h的发芽青稞中的硒残留量进行了检测。经测定,用亚硒酸钠胁迫发芽48 h的发芽青稞中硒的残留量如表3所示,硒残留量低于NY 861—2004 粮食(含谷物、豆类、薯类)及制品中铅、镉、铬、汞、硒、砷、铜、锌等8种元素限量中规定的上限(300 μg/kg)。
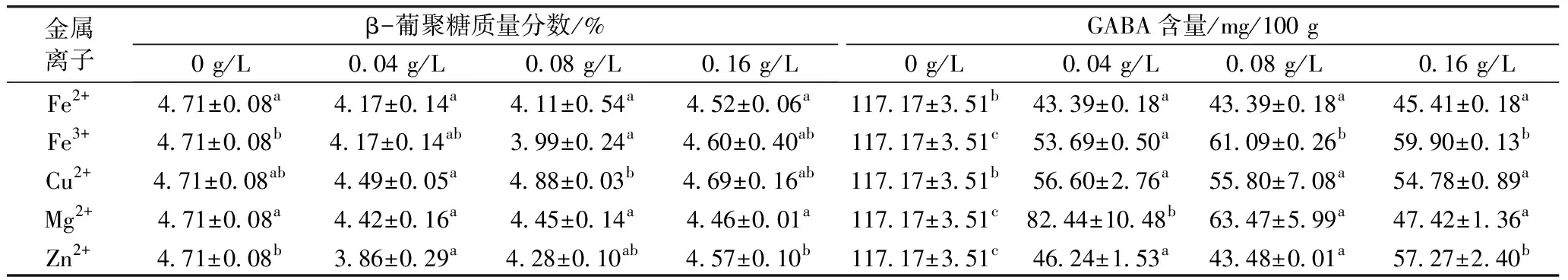
表1 金属离子胁迫萌发对青稞中β-葡聚糖和GABA含量的影响
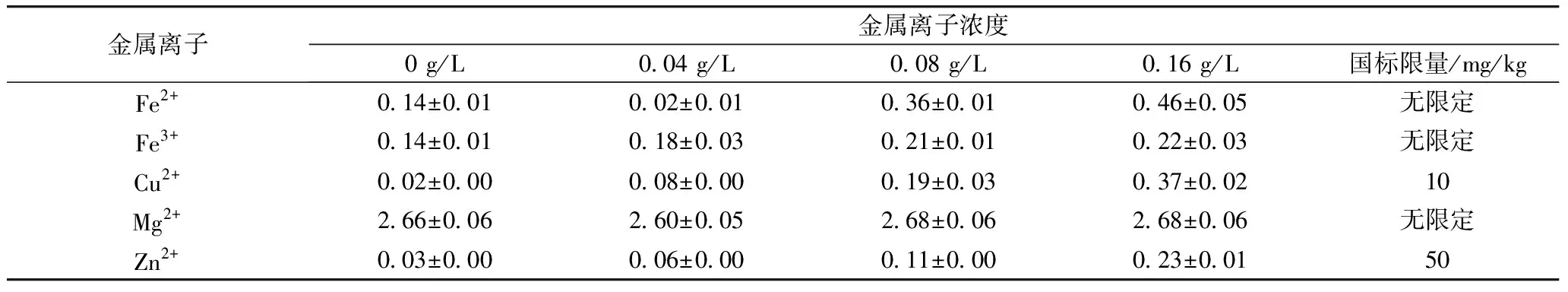
表2 发芽青稞中金属的残留量/mg/kg

表4 低温胁迫萌发对青稞中β-葡聚糖和GABA含量的影响
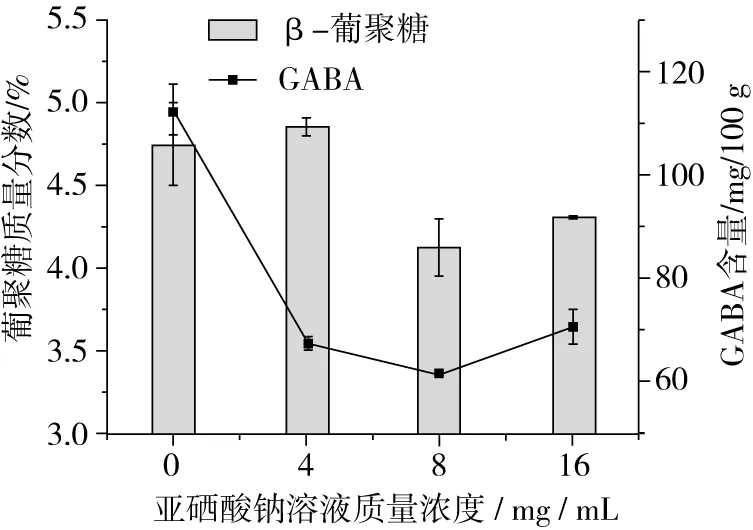
图2 亚硒酸钠胁迫萌发对青稞β-葡聚糖和GABA含量的影响
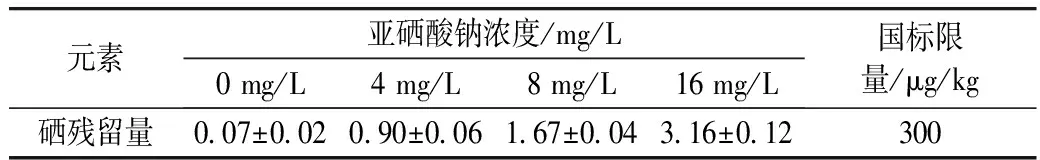
表3 发芽青稞中硒的残留量/μg/kg
2.4 低温胁迫对青稞β-葡聚糖和GABA含量的影响
由表4可知,-20 ℃胁迫条件下β-葡聚糖含量比5、15 ℃高,这可能是由于过低温度破坏了青稞细胞结构导致的[8],青稞发芽率严重下降,抑制了青稞的发芽活力,钝化了大部分β-葡聚糖酶,β-葡聚糖降解很少。5 ℃胁迫条件下β-葡聚糖含量比15 ℃高,这可能是较低的温度抑制了β-葡聚糖酶的活性,β-葡聚糖降解越少。
-20 ℃胁迫条件下发芽青稞中GABA含量显著高于15 ℃的原因是由于胁迫温度过低形成了冰晶体,破坏了植物细胞内部结构,Ca2+、H+等渗入细胞质激活细胞质中钙调素结合区的GAD活性,催化谷氨酸脱羧生成GABA[20],同时催化其转氨降解的酶活性受到抑制,GABA转氨作用减弱,使GABA得以大量积累。15 ℃发芽青稞中GABA含量显著高于5 ℃的原因可能是由于胁迫温度较高时,青稞对外界逆境环境反应不强,GAD激活不彻底,同时GABA转氨作用增强,将GABA氧化降解形成琥珀酸,为TCA循环提供所需碳源,造成了GABA及其合成原料的消耗[21],使得发芽青稞中GABA含量有所下降。
3 结论
青稞籽粒在萌发过程中可以产生GABA,提高了GABA含量,但青稞中β-葡聚糖则逐渐被降解。低温-20 ℃胁迫萌发,可以促进青稞中GABA的大量富集,且部分胁迫萌发方式(低温-20 ℃)可以抑制β-葡聚糖的降解。此外,部分胁迫萌发方式(5种金属离子、亚硒酸钠、低温5 ℃)却促进了β-葡聚糖的降解。
猜你喜欢
星星·诗歌原创(2022年1期)2022-04-16
翠苑(2019年4期)2019-11-11
广东茶业(2019年2期)2019-06-18
农药科学与管理(2019年12期)2019-05-20
中成药(2018年1期)2018-02-02
广东饲料(2016年3期)2016-12-01
西藏科技(2016年8期)2016-09-26
现代农业(2016年6期)2016-02-28
中国药业(2014年24期)2014-05-26
食品工业科技(2014年15期)2014-03-11
