
circLMNB1诱导调控内质网应激介导结直肠癌转移发生
2021-07-16 01:18:34何春萍黄超周瑞孙军阮鹏于红刚姚涛
实用医学杂志 2021年12期
何春萍 黄超 周瑞 孙军 阮鹏 于红刚 姚涛
武汉大学人民医院1消化内科,2神经内科(武汉430060)
结直肠癌(CRC)是最常见的恶性肿瘤之一[1],超过50%的CRC患者将在其一生中发生肝转移[2]。目前对于转移性晚期CRC 尚无有效治疗方法。绝大多数CRC 病例是散发性的和非遗传性的[3]。大量的CRC 零星的肿瘤前病变逐渐积累了遗传和表观遗传修饰,使细胞增殖和存活不受控制,并逐步发展成为具有典型的浸润和转移特性[4]。了解这些转变的分子机制,对于优化CRC 的治疗策略非常重要。环状RNA(circRNA)是一种保守且稳定的共价闭合的RNA,它可能是潜在的疾病标志物[5]。ZENG 等[6]发现,circHIPK3 在结直肠癌的生长与转移过程中起到关键的调控作用。越来越多的研究聚焦于内质网应激在肿瘤中的作用。内质网应激被证明可诱发细胞凋亡[7]。肿瘤的发生发展与细胞的内质网应激失调存在相关性[8]。内质网应激的激活可以诱导CRC 细胞凋亡,抑制肿瘤发生与发展[9]。而circRNA 与内质网应激之间存在调控关系。circHIPK2 可通过调控内质网应激机制,控制星形胶质细胞的激活[10]。然而,目前在结直肠癌中,通过circRNA 调控内质网应激机制,干预肿瘤发生进展的鲜有报道。本研究首先对circLMNB1 在CRC 组织和癌旁组织及细胞系中的表达特征进行探索,随后,在CRC 细胞中对circLMNB1 表达进行干预,进而检测细胞的迁移和侵袭能力,同时对内质网应激机制进行探索。本研究将明确circLMNB1 在结直肠癌的作用机制,为结直肠癌的治疗提供潜在的分子靶点。
1 资料与方法
1.1 一般资料研究收集2017年4月至2018年5月武汉大学人民医院胃肠外科20 例结直肠癌患者结直肠癌组织,并同时收集患者癌缘(3 ~5 cm)处组织作为癌旁组织。病例年龄在40 ~75 岁之间。纳入标准:a.经病理及影像检查确诊为结直肠癌;b. 未合并其他结直肠疾病;c. 未合并其他肿瘤;d.术前无放化疗史。排除标准:a. 病理确诊排除结直肠癌;b.合并其他肿瘤;c.术前经放化疗。研究开展前,经武汉大学人民医院伦理审查委员会批准,所有患者均签署知情同意书。
1.2 主要试剂和细胞系实验中所采用的结直肠癌细胞系LoVO 和HCT116 均购于中科院细胞库。circLMNB1 siRNA 由苏州吉玛生物设计并合成,circLMNB1 过表达载体pcircRNA2.2 由广州伯信生物提供,用于细胞培养的RPMI 1640 细胞基础培养基购自赛默飞旗下品牌Gibco,Transwell 小室由美国康宁公司提供。CHOP(A00311),ATF6(A00655)和GAPDH(A00227)一抗由博士德生物提供,HRP标记的山羊抗兔(ab6721)二抗由Abcam 公司提供。
1.3 细胞培养复苏后的细胞,培养于含10%胎牛血清的RPMI1640 培养基中,在5%CO2、37℃、饱和湿度的培养箱培养,2 d 更换一次培养基。当密度达到85%以上时,进行细胞传代。
1.4 免疫印迹加入RIPA裂解液到细胞中充分混匀,于冰上裂解30 min 后离心,获得总蛋白液。测得样品总蛋白含量后,计算上样量。取20 μg 总蛋白进行SDS-PAGE 凝胶电泳,经转膜、5%脱脂奶粉封闭后,依次进行一抗孵育过夜、TBST 洗涤3 次、二抗室温下孵育1 h,随后进行显色和曝光。
1.5 Transwell 迁移和侵袭实验培养板中加入胰蛋白酶处理后制备单细胞悬液,接种到上室,并放入培养箱中进行培养。实验结束后,取出小室进行固定和染色。取下小室底置于载玻片上,中性树脂封片,观察记录多个视野下的迁移和侵袭的细胞数目。所有实验重复三次。
1.6 实时荧光PCR取50 mg 大小临床组织或5×105个细胞,进行总RNA 的提取。使用TRIzol 方法提取组织或细胞中的总RNA,并将提取的总RNA逆转成为cDNA。cDNA 模板采用如下引物进行qRT-PCR 实验。GAPDH:F,5′-CCAGCCGAGCCACATCGCTC-3′;R,5′-ATGAGCCCCAGCCTTCTCCAT-3′。circLMNB1:F,5′-GCCAAAATTGAATGCTGTCC-3′;R,5′-TGAGATAGCCCAGCAATCCT-3′。以GAPDH 的表达为内参,计算circLMNB1 相对表达水平。
1.7 患者临床特征分析对纳入的结直肠癌患者的基本信息及临床特征,包括性别、年龄、肿瘤大小、转移情况及TNM 分期情况。
1.8 统计学方法采用SPSS 22.0 进行数据分析。两组间数据分析采用t检验方法;多组间统计分析采用单因素方差分析进行差异分析。计量数据采用均数±标准差表示,使用Graphpad 8.0 作图。P<0.05 表示差异有统计学意义。
2 结果
2.1 circLMNB1 的表达特征qRT-PCR 结果显示,circLMNB1 在结直肠癌患者的癌组织中表达水平显著高于癌旁组织(P<0.01,图1A)。另外,circLMNB1 表达水平与临床患者特征之间的关系分析见表1。
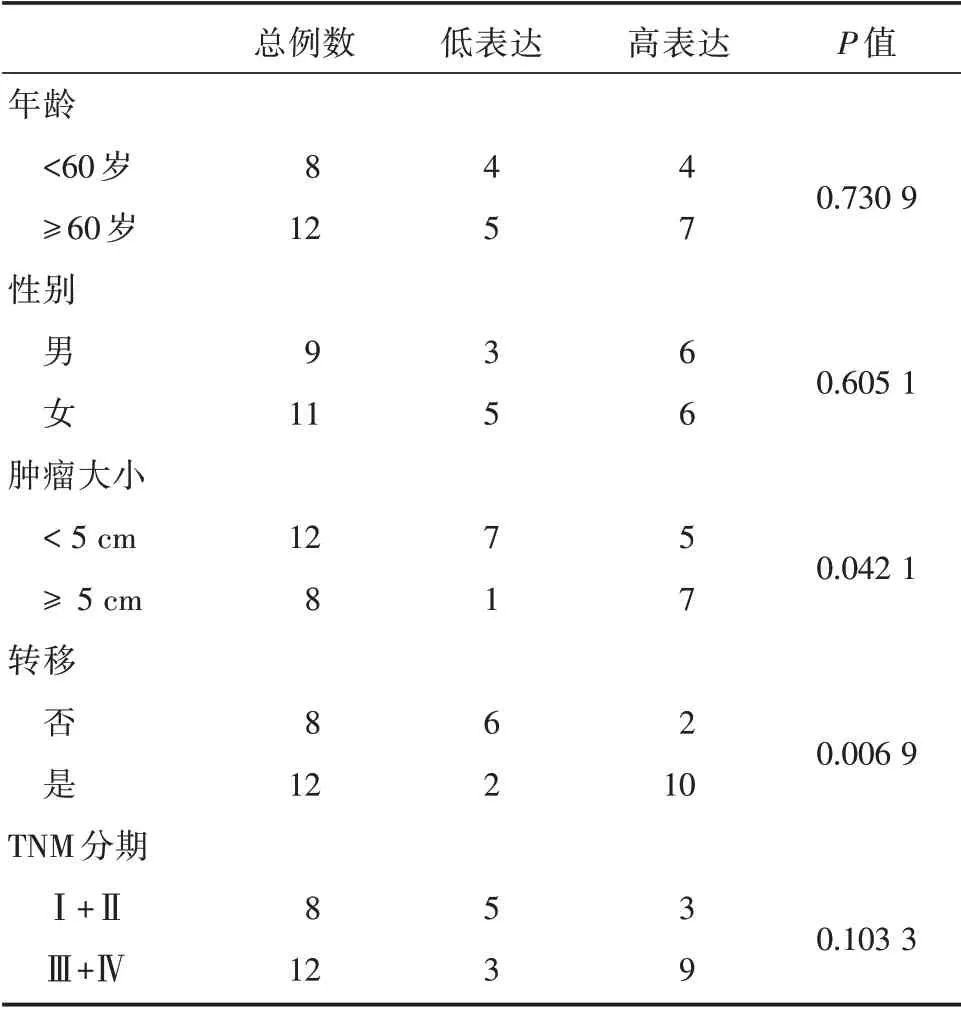
表1 CRC 患者特征Tab.1 Features of patients with CRC 例
circLMNB1 表达水平与年龄和性别均不存在显著性的差异(P>0.05)。而肿瘤大小、转移情况均与circLMNB1 表达水平存在显著的相关性(P<0.05)。然而,TNM 分期与circLMNB1 表达并不存在相关性(P>0.05)见表1。
同时,在人正常结直肠粘膜细胞FHC和结直肠癌细胞系HCT116 和LoVo 中,circLMNB1 的表达水平也存在显著差异。circLMNB1 在LoVo 和HCT116细胞系中的表达水平较FHC 细胞显著上调(P<0.05)。见图1B。
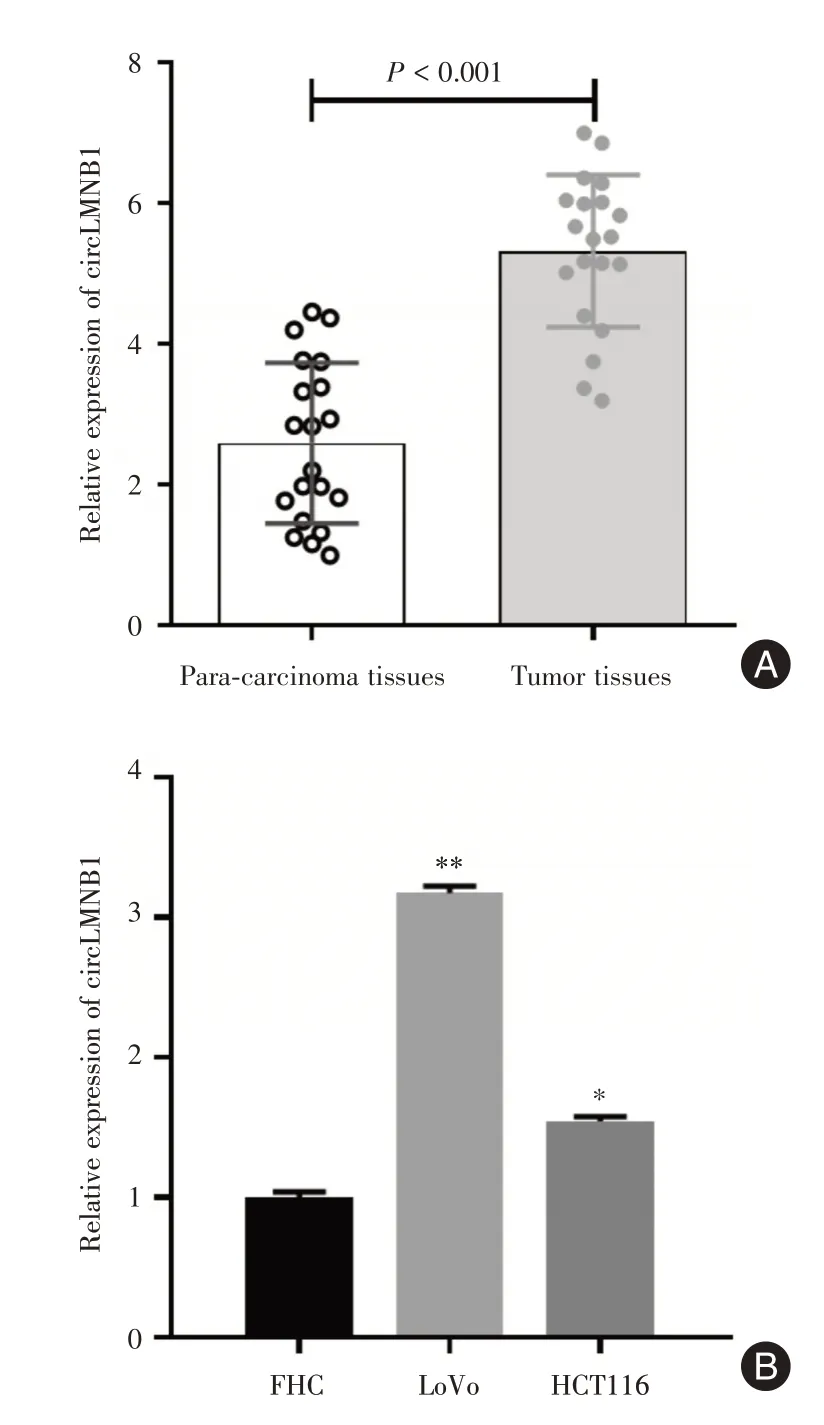
图1 circLMNB1 在结直肠癌临床组织和细胞系中的表达特征Fig.1 Expression characteristics of circLMNB1 in Clinical tissues and Cell Lines of Colorectal Cancer
2.2 circLMNB1 对结直肠癌细胞迁移和侵袭能力的影响分别采用circLMNB1 和circLMNB1 siRNA转染细胞,以对circLMNB1 进行过表达和干扰表达(图2A)。在HCT116 进行circLMNB1 过表达后,其迁移和侵袭能力均出现显著增强(P<0.05);而在LoVo 细胞中,采 用siRNA 对circLMNB1 进行敲降后,细胞的迁移和侵袭能力均显著减弱(P<0.05)。
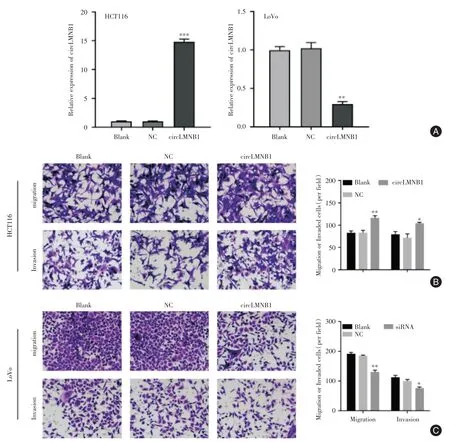
图2 circLMNB1 对结直肠癌细胞迁移、侵袭能力的影响Fig.2 Cell migration and invasion was changed by circLMNB1 knockdown or overexpression in CRC cell line
2.3 circLMNB1 对结直肠癌细胞内质网应激机制影响在HCT116 中对circLMNB1 进行过表达后,CHOP 和ATF6 的表达水平出现显著的下调;而在LoVo 细 胞中,使 用siRNA 对circLMNB1 进行表达干扰后,CHOP 和ATF6 的表达水平则出现显著的上调(图3)。
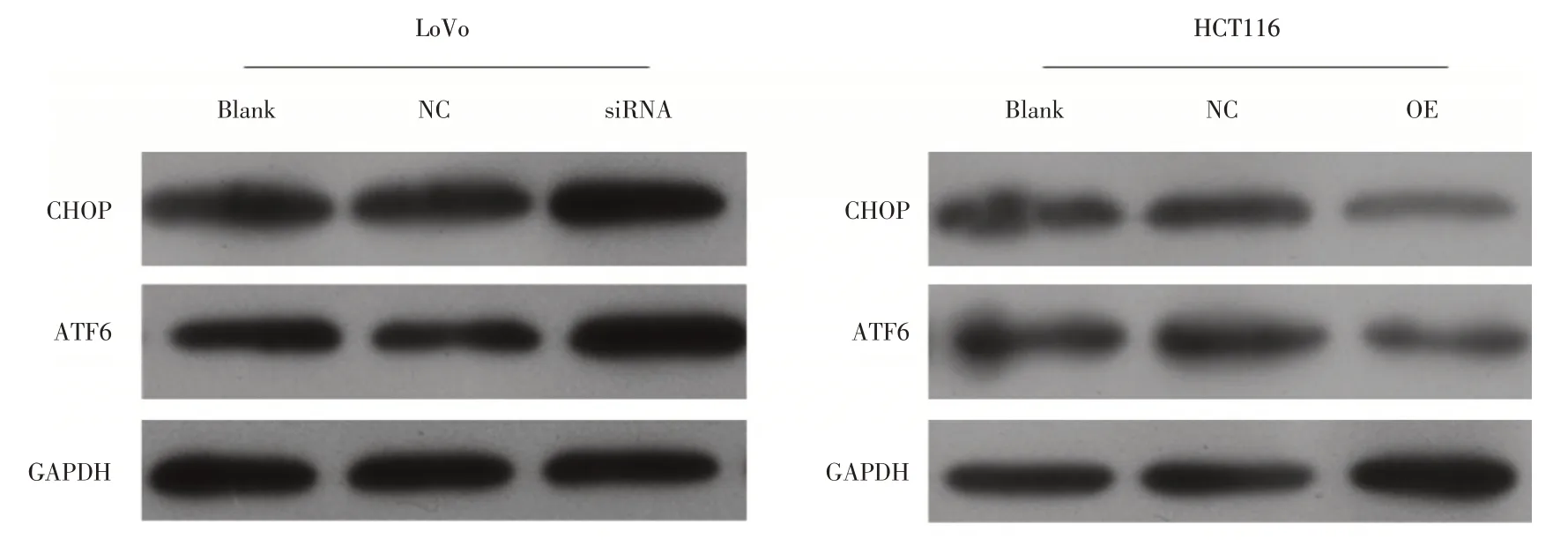
图3 circLMNB1 对结直肠癌细胞内质网相关蛋白CHOP 和ATF6 表达水平影响Fig.3 Expression of CHOP and ATF6 was changed by circLMNB1 knockdown or overexpression in CRC cell line
3 讨论
circRNAs 是一种通过选择性剪接形成的内源性RNA,广泛分布于真核细胞中[11]。环状RNA 由于其封闭的环状结构,不易被内切酶降解,比线性RNA 更稳定[12]。目前被当作肿瘤诊断标记物而进行广泛的研究[13]。如has_circRNA_102209 被证明其在结直肠癌中表达上调,并可通过miR-761对肿瘤的增长与转移进行调控[14]。本研究结果发现,circLMNB1 在结直肠癌组织中的表达较癌旁组织高,同时,在结直肠癌细胞系中的表达水平较正常细胞高;进一步的分析初步得知,circLMNB1表达水平与患者肿瘤的大小、转移情况存在显著的相关性,但仍有待进一步扩大样本,进行综合分析。
内质网应激被证实参与结直肠癌的转移过程[15]。内质网建立了独特的方式来对抗应激,统称为未折叠蛋白反应(unfolded protein response,UPR)[16]。在应对来自外界的刺激时,细胞通常通过一系列的信号分子进行调控,如ATF6、CHOP等[16]。研究表明,ATF6/CHOP 信号参与内质网应激调控,并诱导细胞发生凋亡[17]。XU 等[18]的研究证实,FOXD3 可通过激活内质网应激,促进结直肠癌细胞的凋亡。HUANG 等[19]的最新研究表明,TSPYL5 可通过诱导内质网应激CHOP 等内质网相关蛋白表达,促进结直肠癌细胞的凋亡,抑制细胞的迁移、侵袭等生物学行为。本研究发现,circLMNB1 的过表达引起结直肠癌细胞内质网应激变弱,同时伴随迁移和侵袭能力增强。但是对circLMNB1 进行干扰,内质网应激则增强,同时,细胞迁移和侵袭能力变弱。结果表明,circLMNB1 可以调控内质网应激,具有潜在的调控肿瘤细胞迁移和侵袭能力的作用。
总之,circLMNB1 可作为结直肠癌诊断的生物标记物,它可能通过抑制内质网应激的方式参与结直肠癌的发生与进展。circLMNB1 的这种潜在价值,有望成为临床诊断的标记物及治疗靶标。
猜你喜欢
解放军医学杂志(2021年12期)2022-01-18 03:53:24
现代临床医学(2021年1期)2021-01-26 00:55:52
中国医药指南(2017年3期)2017-11-13 02:56:23
安徽医科大学学报(2016年12期)2017-01-15 14:21:55
西南军医(2016年3期)2016-01-23 02:17:57
山东医药(2015年14期)2016-01-12 00:39:43
中国继续医学教育(2015年6期)2016-01-07 07:38:47
江苏大学学报(医学版)(2015年2期)2015-04-17 06:49:51
中国当代医药(2015年33期)2015-03-01 02:09:08
中国当代医药(2015年30期)2015-03-01 02:08:01
