
重大害虫长林小蠹入侵我国的首次发现与侵染特征*
2021-07-16 03:12:32任利利武海卫宗世祥王传珍刘漪舟骆有庆
林业科学 2021年5期
任利利 陶 静 武海卫 宗世祥 王传珍 华 德 石 娟 刘漪舟 骆有庆
(1.北京林业大学林木有害生物防治北京市重点实验室 北京 100083; 2.山东省林业科学研究院 济南 250014; 3.山东省烟台市森林资源监测保护服务中心 烟台 264003; 4.山东省烟台市牟平区自然资源局 烟台 264100)
长林小蠹Hylurgusligniperda(Fabricius, 1787),异名:BostrichusligniperdaFabricius, 1787;BostrichuselongatusHerbst, 1793;BostrichusflavipesPanzer, 1795;HylesinusflavipesPanzer, 1795;Hylesinusligniperda(Fabricius, 1792);Hylurguselongatus(Herbst, 1793);Hylurgusflavipes(Panzer, 1795);HylurguslongulusKolenati, 1846。属鞘翅目(Coleoptera)象虫科(Curculionidae)小蠹亚科(Scolytinae)切梢小蠹族(Tomicini)林小蠹属(HylurgusLatreille),英文名为 red-haired pine bark beetle。
林小蠹属在世界范围内已知有3种(Alonso-Zarazagaetal., 2017),寄主树种大多为松属(Pinusspp.)植物。欧洲报道一种为米林小蠹HylurgusmicklitziWachtl, 1881,主要危害阿勒颇松(Pinushalepensis)、 海岸松(P.pinaster)、 意大利松(P.pinea)和P.nigrasubsp.pallasiana(Wood, 1992; Brightetal., 1997; Alonso-Zarazagaetal., 2017); 印度分布一种(Wood, 1985),HylurgusindicusWood, 1985,作者描述的4只雌性标本采集于喜马拉雅长叶松(Pinusroxburghii)(原文中Pinuslongifolia为其异名)木段,除此新种记录外,该种无其他研究报道。我国无该属昆虫分布的报道。
长林小蠹原产欧洲南部及非洲北部的地中海沿岸。近90年来,已传播至各大洲,包括欧洲全部,非洲北部及其南端,亚洲的日本、韩国、土耳其、斯里兰卡,大洋洲的澳大利亚、新西兰,南美洲的巴西、智利、乌拉圭以及北美洲的美国。据报道,传入日本的时间推测早于1931年; 于 1942 年传入澳大利亚, 1956 年传入乌拉圭, 1973 年传入南非, 1974年传入新西兰, 1987年传入智利, 2000—2001年于纽约发现越冬种群并明确传入美国, 2007 年在加州发现; 于2007 年传入阿根廷, 2018年报道传入韩国(Parketal., 2017; Wood, 1992; Alonzo-Zarazagaetal., 2017; El-Khouryetal., 2019; Swan, 1942; Schedletal., 1966; Schedl, 1982; Ciesla, 1988; Liuetal., 2007)。我国没有长林小蠹分布的报道,但已将其列为进境植物检疫性有害生物,国际植物保护公约(IPPC)规定其为需要检疫除害处理的种类(International Plant Protection Commission, 2005; 国家质量监督检验检疫总局, 2017)。
该虫在世界范围的寄主树种超过20种(Eglitis, 2001),包括: 2针松的欧洲赤松(P.sylvestris)、意大利黑松(P.brutia)、阿勒颇松、土耳其松(P.halepensisvar.brutia)、欧洲黑松(P.nigra)、P.nigrasubsp.pallasiana、奥地利黑松 (P.nigravar.austriaca)、克里木松(P.nigravar.caramanica)、海岸松、日本黑松 (P.thunbergii)、赤松(P.densiflora), 2~3针松和3针松的湿地松(P.elliottii)、加那利松 (P.canariensis)、展叶松(P.patula)、辐射松(P.radiata), 5针松的山松(P.montezumae)、意大利松、北美乔松(P.strobus), 其他寄主还包括冷杉属(Abiesspp.)、落叶松属(Larixspp.)、云杉属(Piceaspp.)和黄杉属(Pseudotsugaspp.)的部分树种。
据国外文献报道,长林小蠹属树皮小蠹类,通常侵染新伐木、伐桩、衰弱木,健康树也可受危害,是次期性害虫中的先锋种; 钻蛀坑道于树干基部、根颈和根部。受害树木树皮和根部外表可见长林小蠹的侵入孔或羽化孔,幼虫在坑道内取食时,树皮表面有暗色蛀屑排出(Browne, 1968; Ciesla, 1988)。长林小蠹在法国一般1年2代,在智利1年3代,世代重叠使其在全年多数时间均能传播与危害; 成虫的活动受到温度、湿度和光照的影响。羽化后出孔前的成虫具有蔽光性; 出孔后扬飞期温度高于20 ℃时具有正趋光性,侵入寄主或入土后恢复蔽光性; 湿度同时也会影响其趋光习性; 性成熟前有在新生松枝上钻蛀补充营养的习性(Fabreetal., 1974; Ciesla, 1988; Eglitis, 2001)。长林小蠹雌虫首先侵入根皮下,在根皮的形成层中筑造一个交配室并诱集雄虫进入,雌虫与雄虫交配后(一夫一妻制),雌虫建造 1 个长而弯曲的母坑道,并在坑道两端的刻痕中产卵,据报道每雌虫最多产卵500粒(Fabreetal., 1974)。雌虫可钻蛀数条母坑道,单头雌虫蛀母坑道总长度可达3 m。幼虫在根皮下取食韧皮部与边材,子坑道不规则,不会形成特定的坑道结构; 幼虫共4龄,老熟后化蛹,以幼虫或成虫越冬,春季羽化飞出后,时有群聚现象(Browne, 1968)。
另据报道,除了直接危害松树外,长林小蠹可携带能侵染与危害寄主树木的半帚霉属(Leptographium)、蛇口壳属(Ophiostoma)、镰刀菌属(Fusarium)、链格孢属(Alternaria)、球壳孢属(Sphaeropsis)等真菌(Davydenkoetal., 2014; Jankowiaketal., 2013; Kimetal., 2011; Rayetal.2006),不仅易导致树木的死亡,且严重影响成品木材的品质。在欧洲,其是伞滑韧属(Bursaphelenchus)几种线虫,如Bursaphelenchushellenicu、B.sexdentati和B.tusciae等的传播媒介(Ambrogionietal., 1998; Mejrietal., 2016; Carletti, 2008; Carlettietal., 2008; Penasetal., 2006; 2004; Torrinietal., 2020)。长林小蠹成虫具有很强的自然传播能力,可飞行达数千米; 在澳大利亚,该虫 18个月内传播了 25 km(杨晓军等, 2002); 在智利,自 20世纪 80年代传入该虫后,到 1991年就已扩散至辐射松的所有分布区(Mauseletal., 2006)。
长林小蠹是我国口岸检疫中截获次数最多的林业害虫之一,公开报道的海关截获记录显示,多个省市口岸在来自智利、新西兰、澳大利亚、巴西、乌拉圭等多国的原木、锯材和集装箱中,均截获到了该虫。最早于1986年4月14日,天津动植物检疫所曾在进口的智利原木上截获到该虫(魏厚德等, 1991)。2019年4月19日,河北曹妃甸海关在进口的新西兰辐射松原木中截获此虫。2005—2015年间,我国口岸共截获长林小蠹4 049批次(梁振等, 2017年),被认为是重要的“林木杀手”。
目前我国对长林小蠹生物学特性、适生性、防治技术等方面的研究少,认知度低,极大地增加了检疫、监测和防治的难度。本研究利用形态学、分子生物学和危害特征分析的方法对其进行了鉴定,首次发现并明确了该虫在我国的入侵、定殖并严重危害,初步调查了其危害特性。
1 材料与方法
1.1 虫源采集
样品采集于山东省烟台市牟平区沿海防护林带(37.457 241°N,121.851 217°E)。2020年10月至11月,在日本黑松根皮下,采集到一种严重危害根部的小蠹,带回北京林业大学森林保护学科检疫实验室。
1.2 形态特征观察
使用蔡司AxioZoom V16体式显微镜对该小蠹的幼虫、蛹及成虫形态特征进行观察、拍照。参考Balachowsky (1949)、Wood (1986)、袁克等(2007)、王仿等(2006)以及《中华人民共和国出入境检验检疫行业标准〈长林小蠹检疫鉴定方法〉》(中华人民共和国国家质量监督检验检疫总局, 2006)对该虫形态特征进行检索与比对。
1.3 分子鉴定
随机取不同寄主单株中的成虫12头和幼虫5头,分别提取单头小蠹的基因组 DNA,用于分子鉴定。
DNA 提取: 使用微量 DNA 提取试剂盒 AxyPrep Multisource Genomic DNA Miniprep Kit,(Axygen,美国),按说明书步骤提取样品 DNA; 使用超微量紫外线分光光度计 NanoDrop 2000(Thermo Fisher Scientific Inc.,美国)检测样品 DNA 浓度。
PCR 扩增: 本试验以上述方法提取的 DNA 为模板,扩增核糖体28S和线粒体 COⅠ基因片段。引物序列见表1。PCR 扩增体系为: 2×Es Taq MasterMix(CWBIO,CW0690M)20 μL,正向引物 1 μL,反向引物 1 μL,DNA 模板 2 μL 和超纯水 16 μL 共 40 μL。PCR 反应程序为: 94 ℃预变性 3 min; 94 ℃变性 45 s,参照表1中温度退火 1 min,72 ℃延伸 1 min,共进 行 40 次循环; 72 ℃延伸 10 min; 4 ℃保存。

表1 引物序列信息Tab.1 Primer sequence information
PCR 扩增产物测序: PCR 扩增完成的产物送北京睿博兴科生物技术有限公司进行双向测序。
序列比对: 拼接后的样本28S和COⅠ序列进行了Blast(NCBI)比对,使用Blastn、 Standard database、High similar sequences (mega blast 程序)。
面对激烈的市场竞争,HM国际货代企业应当努力增强实力,提高市场话语权以及服务专业性,为客户争取更多的利益。
基于28S序列建树: 参考黄蓬英等(2018)、 McKenna等 (2009)、Sequeira等 (2002)、Jordal等 (2011)、Susoy等(2014)、Pistone等 (2016) 以及Pistone等 (2018) 的研究,随机选取3条试验样本序列(NCBI 登录号MW725426、MW725427、MW725428),下载 NCBI 数据库中相关序列(表2),使用Bothrosternusfoveatus作为外群。

表2 相关建树序列信息Tab.2 Sequence information in tree construction
使用PhyloSuite (Zhangetal., 2020)及相关插件程序建树,使用MAFFT (Katohetal., 2013)进行多重序列比对,使用ModelFinder (Kalyaanamoorthyetal., 2017)选择最适建树模型,使用IQ-TREE (Nguyenetal., 2015)构建ML树(Maximum likelihood),参数如下: 5 000 ultrafast bootstraps, approximate Bayes 检验和 Shimodaira-Hasegawa-like approximate likelihood-ratio 检验(Hoangetal., 2018); 进化树文件由iTOL(https:∥itol.embl.de)使用Root mid point 模式成图。
基于COⅠ序列的单倍型分析: 下载 NCBI 数据库中长林小蠹COⅠ序列JQ015131、JQ015128、JQ015130和JQ015129 (经通讯联系,明确为美国研究者上传,样本采集地不详), AY040292(新西兰奥克兰样本), HM002621(经通讯联系,明确为深圳口岸截获来自新西兰辐射松原木携带的样本)。随机选取样品12条成虫序列和5条幼虫序列(NCBI 登录号为MW726016、MW726017、MW726018、MW726019、MW726020、MW726021、MW726022、MW726023、MW726024、MW726025、MW726026、MW726027、MW726028、MW726029、MW726030、MW726031和MW726032),多重序列比对方法同上,除去前后冗余碱基序列,所得长林小蠹COⅠ序列长度为676 bp。使用软件DNAsp v6(Libradoetal., 2009)计算遗传多样性的参数,包括DNA序列变异位点数量、核苷酸多样性、单倍型数量、单倍型多样性以及各单倍型的种群组成。使用PopART(http:∥PopART.otago.ac.nz)构建单倍型网络图。
1.4 危害特征观察
2020年11月,在山东省烟台市牟平区沿海防护林带的被害日本黑松林内进行踏查和树干、根解析。依据被害木树冠颜色,选择绿色树冠5棵,黄绿色树冠21棵,以及红冠木5棵; 伐倒后将样木根系用吊车从沙地中整体拔出,对该小蠹的危害特征进行观察与统计。
2 结果与分析
2.1 形态特征鉴别
2.1.1 切梢小蠹族的主要形态鉴别特征 复眼完整,纵椭圆形; 前胸背板前半部无瘤齿; 鞘翅2基缘呈并列双凸弧线(图1)。

图1 长林小蠹符合切梢小蠹族的关键形态特征Fig. 1 Hylurgus ligniperda conforms to the key morphological characteristics of the Tribe Tomicinia. 复眼椭圆形、完整(长林小蠹)Compound eye oval, complete (H. ligniperda); b. 前胸背板及鞘翅基缘(长林小蠹)Pronotum and elytra base (H. ligniperda).
2.1.2 长林小蠹与常见相似属种类的区别 1)与切梢小蠹属(Tomicus)的区别 林小蠹属前足基节窝相连; 前胸背板较狭长,长为宽的1倍左右,前1/3轻微缢缩; 鞘翅沟间部毛多列。切梢小蠹属前足基节窝微分离; 前胸背板较粗壮,长为宽的0.85倍,前1/3强烈缢缩; 鞘翅沟间部毛单列(图2)。

图2 长林小蠹与切梢小蠹属形态特征的对比Fig. 2 Comparison of the morphological characteristics of Hylurgus ligniperda and the Genus Tomicusa. 前足基节窝相连(长林小蠹)Procoxae contiguous (H. ligniperda); b. 前足基节窝微分离(横坑切梢小蠹)Procoxae moderately separated (Tomicus minor); c. 前胸背板较狭长(长林小蠹)Pronotum more slender (H. ligniperda); d. 前胸背板较粗壮(横坑切梢小蠹)Pronotum stouter (T. minor); e. 鞘翅沟间部毛多列(长林小蠹)Erect interstrial setae abundant, confused (H. ligniperda); f. 鞘翅沟间部毛单列(横坑切梢小蠹)Erect interstrial setae in uniseriate rows (T. minor).

图3 长林小蠹与大小蠹属形态特征的对比Fig. 3 Comparison of the morphological characteristics of Hylurgus ligniperda and the Genus Dendroctonusa. 触角鞭节6节(长林小蠹)Antennal funicle 6 segmented (H. ligniperda);b. 触角鞭节5节(红脂大小蠹) Antennal funicle 5 segmented (Dendroctonus valens).
3)与根小蠹属(Hylastes)的区别 林小蠹属鞘翅基缘隆起; 触角鞭节6节; 体表被长毛,无鳞片。根小蠹属鞘翅基缘不隆起; 触角鞭节7节; 体表被短刚毛或鳞片(图4)。

图4 长林小蠹与根小蠹属形态特征的对比Fig. 4 Comparison of the morphological characteristics of Hylurgus ligniperda and the Genus Hylastesa. 鞘翅基缘隆起,体表被长毛(长林小蠹)Crenulations on elytral bases usually well developed, body surface setae usually hairlike (H. ligniperda); b. 鞘翅基缘不隆起,体表被短刚毛或鳞片(黑根小蠹)Crenulations on elytral bases usually poorly developed,body surface setae scalelike or bristle (Hylastes parallelus);c. 触角鞭节6节(长林小蠹)Antennal funicle 6 segmented (H. ligniperda); d. 触角鞭节7节(黑根小蠹)Antennal funicle 7 segmented (H. parallelus).
2.1.3 长林小蠹的形态特征 1)成虫 如图5所示,体长4~5.7 mm,体圆筒形,黄褐色至黑色。触角鞭节6节,锤状部4节,复眼长椭圆形; 额面正下方有一黑色瘤; 头腹面前咽缝三角区下陷。鞘翅基缘具颜色较深的锯齿状边缘; 鞘翅沟上毎刻点的中央各具一黄色短茸毛; 沟间粗糙,具横皱褶,每沟间具有4~5排排列不甚规则且向后斜伸的茸毛,鞘翅两侧的茸毛比前胸背板两侧的要短,且鞘翅上的茸毛愈向翅末愈长,在斜面上,茸毛形成簇状。

图5 长林小蠹识别的关键形态特征Fig. 5 The key morphological characteristics of the identification of Hylurgus ligniperdaa. 成虫背面观Adult dorsal view; b. 额面正下方有一黑色瘤Frons with a median tubercle; c. 头腹面前咽缝三角区下陷Gular triangle with depression.
2)其他虫态形态特征 老熟幼虫体长约4.1 ~ 5.9 mm; 体圆柱形,粗壮,弓曲,黄白色。头部黄褐色,胸节和腹节均无足(图6)。

图6 长林小蠹幼虫和蛹的形态Fig. 6 The morphological characteristics of the larvae and pupae of Hylurgus ligniperda a. 蛹Pupae; b. 老熟幼虫The last larval instar.
2.2 分子鉴定
采集于烟台日本黑松根部的12头成虫小蠹样本及5头幼虫样本,属同一单倍型。通过Blast比对,在28S 基因部分序列(647 bp)与已报道的长林小蠹序列JX263769(新西兰奥克兰样本),MH005064(厦门口岸截获来自于澳大利亚进口辐射松原木携带样本),KJ531125(德国样本)相似性分别为: 100%,99.85%和99.69%。在COⅠ 基因部分序列(676 bp)与已报道的长林小蠹序列HM002621(深圳口岸截获来自于新西兰辐射松样本)相似性达100%。
基于28S基因序列的最大似然树结果显示(图7),采集于山东烟台的样本序列(SDYT H1—3),与口岸截获来自于澳大利亚进口辐射松原木携带的样本序列、德国样本序列、新西兰奥克兰样本序列以100%的置信系数聚为一支。长林小蠹一支以99%的置信系数与同属种类米林小蠹聚为一支。

图7 基于28S 基因序列的ML 系统发育树 (圆圈代表ultrafast bootstrap置信系数)Fig. 7 ML phylogenetic tree based on 28S gene sequence (circle represents ultrafast bootstrap support)
参与单倍型数据分析的3个来源的长林小蠹样本共计23个,其中山东烟台17个样本。线粒体COⅠ基因序列长度为 676 bp,总体的核苷酸多样性(Pi)为0.008 95,共含有变异位点17个; 共有6个单倍型; 所有样本总体单倍型多样性为0.395 3。山东烟台样本均属于同一单倍型,与深圳口岸截获来自于新西兰辐射松的长林小蠹样本属于同一单倍型(图8)。
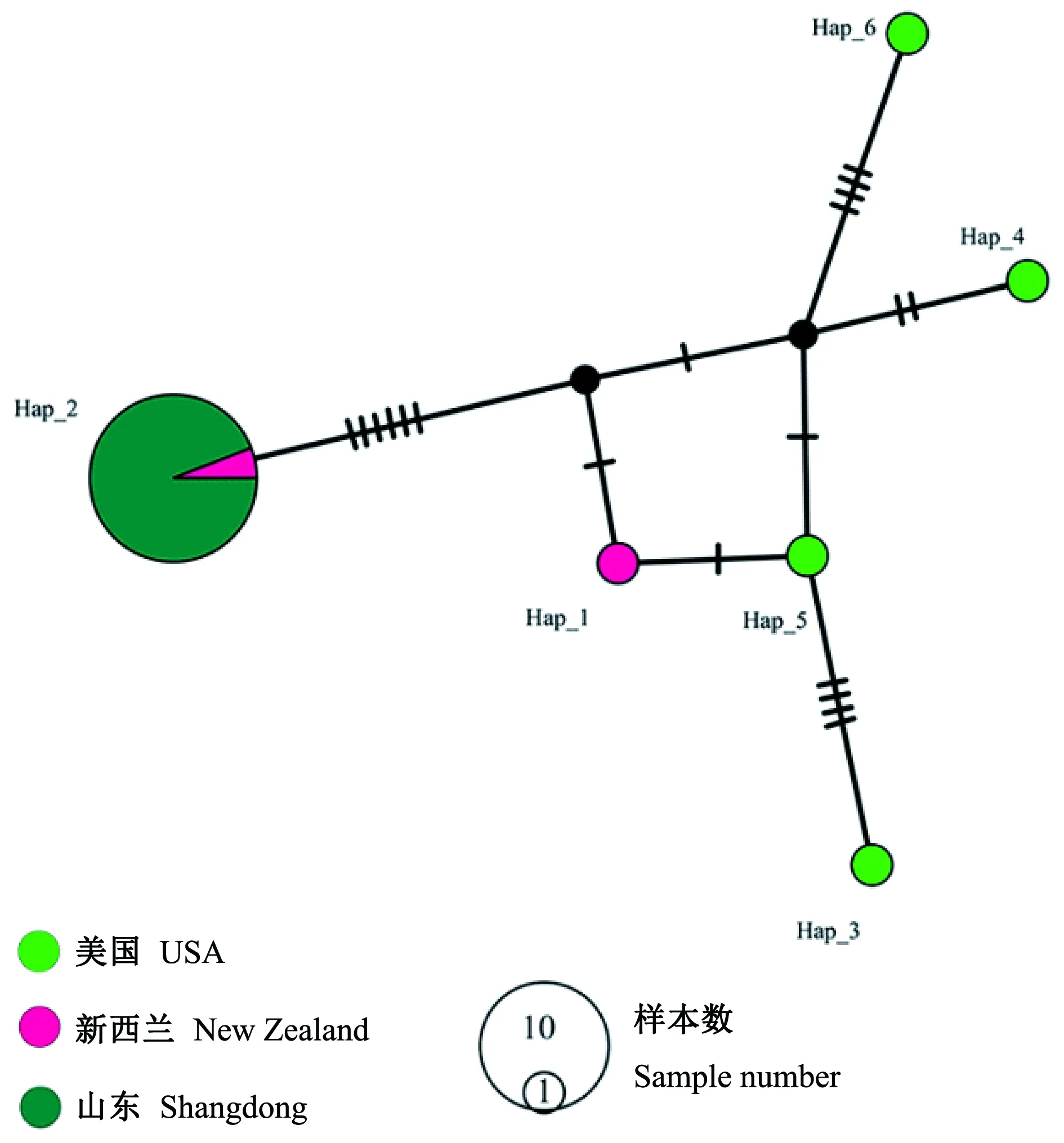
图8 基于676 bp COⅠ片段的23个长林小蠹样本单倍型网络Fig. 8 Haplotype network of 23 global Hylurgus ligniperda samples based on 676 bp COⅠ sequences Hap_1-6: 6个单倍型Six haplotypes.
经过基于COⅠ和 28S 基因序列的DNA条码分析,采集于山东烟台的小蠹样本为长林小蠹,系首次发现该虫在中国定殖。
2.3 危害特征
据初步调查,该虫在牟平区的寄主树种为我国引进的外来树种日本黑松。危害严重,在挖出的枯死松树根系中,侵害株率达100%。长林小蠹危害部位可从树干下部至根系,单坑道长达1.4 m; 距离干基1.8 m的主干上也可受害。成虫也可直接穿越土壤侵害根系和羽化出孔,从土壤入侵根部时,可向干基部或根端部的方向挖掘坑道; 在侧根上可侵害远离根际7.4 m处的根,可危害细至0.5 cm的根; 根系中危害深达土壤内1.5 m(图9a、b)。严重受害的树木根系中,半米段根(根径12 cm左右)的虫口数可达62头。被侵害的根皮表面可见侵入孔、羽化孔和通气孔; 被侵害的根皮下充满棕色细粉状虫粪; 母坑道长,数量较多时交错在一起; 无明显的子坑道; 蛹室明显,分布于母坑道两侧。可见成虫或幼虫在坑道内排队或聚集钻蛀; 也可见成虫在坑道内交配(图9c—g)。该虫可独立危害树冠呈黄绿色、流脂丰富的黑松根部,初步判断其侵入顺序早于与其混合危害的其他多种钻蛀性害虫,如褐梗天牛(Arthopalisrusticus)、梢小蠹(Cryphalussp.)、角胫象(Shirahoshizosp.)等; 发现受该虫侵染的树木中,蓝变菌侵染严重。寄主新鲜的伤口对成虫的诱集能力强, 2020年11月初仍有成虫在林间活动,已发现的越冬虫态有老熟幼虫、蛹和成虫, 2021年3月初始见成虫在林间活动。
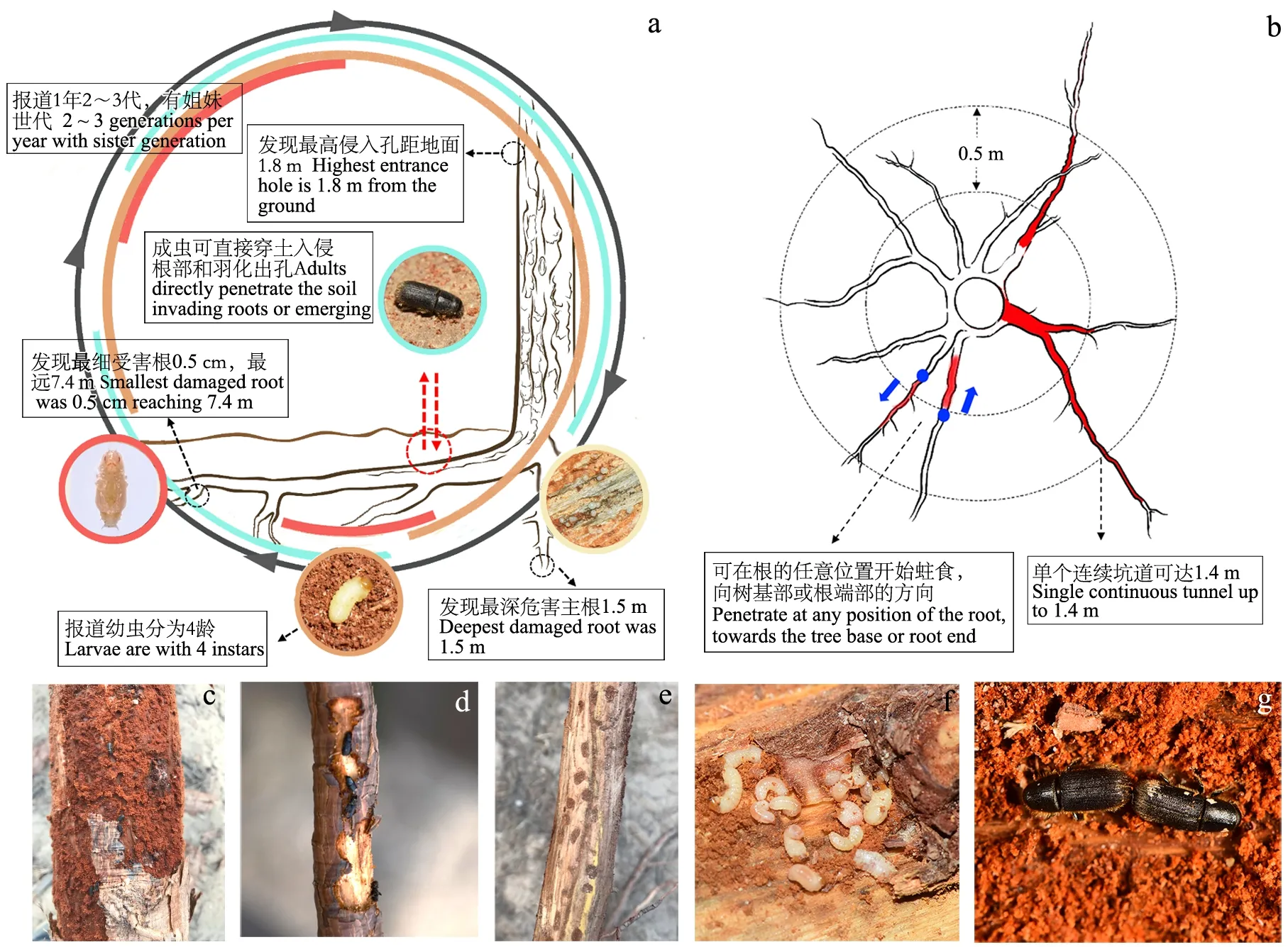
图9 长林小蠹危害特征初步观察Fig. 9 Preliminary observation results of the damaging characteristics of Hylurgus ligniperdaa. 长林小蠹危害特点示意Schematic diagram of the damaging characteristics of H. ligniperda; b. 长林小蠹危害根部特点示意Schematic diagram of the damaging characteristics of H. ligniperda to roots; c. 危害后根皮下的虫粪The beetle frass under the root bark; d. 成虫聚集钻蛀坑道Adults gathering and boring the tunnel; e. 坑道内蛹室Pupal chambers in the tunnel; f. 聚集危害的幼虫Larvae gathering and damaging; g. 坑道内成虫交配Mating of adults in the tunnel.
3 结论
经过资料查证、近缘种形态鉴别、基因条码比对、危害特征分析,以及原产地与入侵发现地自然生态地理和寄主树种的一致性等一系列的核实比较与综合判断,可明确长林小蠹这种外来有害物种已入侵我国,在我国山东烟台定殖,并严重危害日本黑松。长林小蠹危害性强,成虫可直接进入土壤侵入树木根部,甚至可危害远离根基约7 m以上0.5 cm的细根,可先期侵害亚健康树木并致死。该虫隐蔽性危害,地上无明显虫粪等特殊易辨的危害特征,监测、防控难度极大。长林小蠹在世界各大洲之间的入侵历史具有国际重大入侵生物的基本特点,即传播扩散时间长、入侵地分布广,且寄主树种多、环境适应性强。本文为深入研究长林小蠹的特征和快速鉴定方法奠定了基础,便于提高鉴定、检验检疫的针对性和有效性; 初步调查了该虫的危害特征,可为有效防控技术的研发提供路径。同时,该虫在胶东半岛与松材线虫混合发生的关系等也亟待研究。
猜你喜欢
中国机械工程(2024年1期)2024-02-10 16:20:12
青海农林科技(2022年3期)2022-09-26 03:16:18
Acta Mathematica Scientia(English Series)(2022年3期)2022-06-25 02:13:00
中国银幕(2022年4期)2022-04-07 21:25:47
海外星云(2021年14期)2021-10-17 15:55:34
海外星云 (2021年14期)2021-10-03 11:25:48
林业科学(2021年12期)2021-02-12 06:46:22
浙江林业科技(2019年5期)2019-01-10 16:43:51
中南林业科技大学学报(2017年9期)2017-12-11 01:25:02
江苏大学学报(自然科学版)(2013年1期)2013-08-22 06:23:52
