
不同种源野生北沙参种子表型及萌发特性的研究
2021-07-16 06:02李丽霞逄玉娟周淑诺姜明媛张立培卞福花
种子 2021年6期
李丽霞, 逄玉娟, 高 燕, 周淑诺, 姜明媛, 冯 磊, 张立培, 卞福花
(1.烟台大学生命科学学院, 山东 烟台 264005; 2.昆嵛山林场, 山东 烟台 264112;3.烟台吉恩生物科技有限公司, 山东 烟台 264006)
北沙参又名珊瑚菜(GlehnialittoralisFr. Schmidt ex Miq.),隶属于伞形科珊瑚菜属,国内主要分布于山东、内蒙古、河北,国外在美洲东海岸、日本、朝鲜亦有分布[1]。北沙参作为临床常用中药,属补阴药,具有生津滋阴、益气养阴、利肺益胃的功能,用于治疗咽干口渴、肺热劳嗽、干咳痰血、胃阴不足等症[2],对2016年我国出现的寨卡病毒和2020年我国流行的新型冠状病毒所致症状均具有一定疗效,且作为针对气阴两虚证的基本方药之一,被应用于国家卫生健康委员会公布的诊疗方案中[3-4]。北沙参的年需求量在5 000 t左右,国内已有较大面积栽培,市场上主要是人工栽培品种[5]。目前对北沙参开展的研究中所用北沙参绝大多数为已有的栽培品种,且研究内容多集中于种植管理[6-7]、化学成分[8-9]、药理作用及临床应用[10-12]方面,而种子分级、质量标准和质量控制方面仍不完善,对北沙参野生种源开发以及选种育种等方面亦存较大的研究空间[13]。近年随着北沙参市场需求量的上涨,合理利用开发野生北沙参资源,挖掘具有优良潜力的种质进而培育高产优质新品种成为亟待研发的课题。因此,本研究从全国北沙参主要分布地区的山东和辽宁选取5个不同地区的野生北沙参种子,从种子表型特征及种子萌发特性方面展开研究,以期为深入了解北沙参种子的萌发机制以及为北沙参野生种质资源进一步开发利用、选育鉴定优质种源提供一定的理论依据。
1 材料与方法
1.1 材料采集
供试材料于2019年6-7月分别采自山东海阳凤城 (HY)、即墨田横岛(THD)、日照海滨国家森林公园(RZ)和辽宁瓦房店情人岛(QRD)、葫芦岛金石滩(JST)。
1.2 种子长、宽、面积及千粒重测定
随机选取不同种源果实各30粒,用智能考种分析系统测长、宽和面积,重复3次;以上果实每次测定完成后去掉果皮,得到的种子用同样的方法测定相应的参数。随机选取不同种源果实各1 000粒,用电子天平称量果实千粒重,剥掉果皮后继续称量种子的千粒重,重复3次。
1.3 种子萌发实验
随机选取足量的不同种源北沙参果实,置于4 ℃冰箱内湿沙贮藏4个月,使实验的种子均经过低温后熟的过程,翌年4月剥去果皮,进行发芽实验。利用恒温培养箱设置5个温度梯度(10 ℃、15 ℃、20 ℃、25 ℃和30 ℃),每个温度放置种子30粒,置于平皿内润湿的双层滤纸上进行种子萌发,胚根长至种子2倍长度为发芽标准。每个处理设3个重复。
分别在种子播种后3 d、5 d、7 d时,对20 ℃、25 ℃和30 ℃处理的5个种源种子的发芽粒数进行统计,15 ℃处理所有种子继续统计发芽数至第10天,10 ℃处理所有种子持续统计第13天、第16天直至第19天,以连续3 d无新种子发芽视为萌发结束。根据发芽种子数与供试种子总数的比值来计算发芽率。
1.4 数据统计分析方法
利用SPSS 22.0统计软件对各数据均值进行单因素方差分析(多重比较采用Duncan检验)及相关性分析,测定数据以均值±标准差表示,p<0.05表示差异显著,p<0.01表示差异极显著。
2 结果与分析
2.1 不同种源北沙参果实和种子的特征
由表1可知,不同种源北沙参的果实面积、果实长度、果实宽度大小排序依次为:田横岛>日照>海阳>情人岛>金石滩;果实千粒重大小排序依次为:日照>田横岛>海阳>情人岛>金石滩。统计分析表明,在果实大小的多个测定指标包括果实长度、面积及重量上,金石滩种源的北沙参测定值显著低于其他种源的果实的相应值(p<0.05)。果实较大的为日照及田横岛种源的北沙参,但二者千粒重数值接近,其差异并未达到显著水平(p>0.05),而与其他种源的果实千粒重差异均达到显著水平(p<0.05)。

表1 不同种源北沙参果实形态指标
不同种源北沙参种子的形态指标见表2。分析可知,种子面积大小及种子长度排序依次为:田横岛>日照>海阳>情人岛>金石滩;种子宽度由大小排序依次为:日照>田横岛>情人岛>海阳>金石滩;种子千粒重大小排序依次为:田横岛>日照>海阳>金石滩>情人岛,其中田横岛种子千粒重为最大值9.71 g,其重量达到最小值即情人岛种子千粒重的1.42倍。通过差异显著性分析可知,在种子宽度和面积及千粒重指标测定中,情人岛及金石滩种源的北沙参种子均显著低于田横岛及日照种源的种子(p<0.05)。

表2 不同种源北沙参种子形态指标
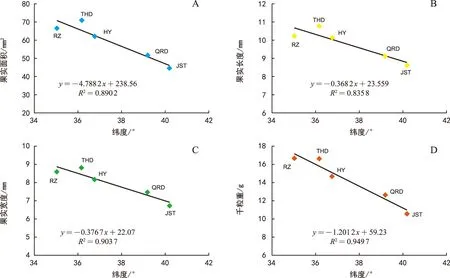
注:A为面积;B为长度;C为宽度;D为千粒重。下同。图1 不同种源北沙参果实特征与纬度的相关性分析Fig.1 Correlation analysis of fruit characteristics and latitude of G. littoralis from different provenances
2.2 不同种源北沙参果实和种子特征与纬度的相关性
相关分析结果(图1)表明,纬度与北沙参果实面积、长度和宽度均呈显著负相关 (p<0.05),与千粒重呈极显著负相关 (p<0.01) (图1)。对于北沙参种子来说,纬度则仅与种子千粒重呈显著负相关 (p<0.05),与种子面积、长度及宽度的相关性均未达到显著水平(p>0.05) (图2)。
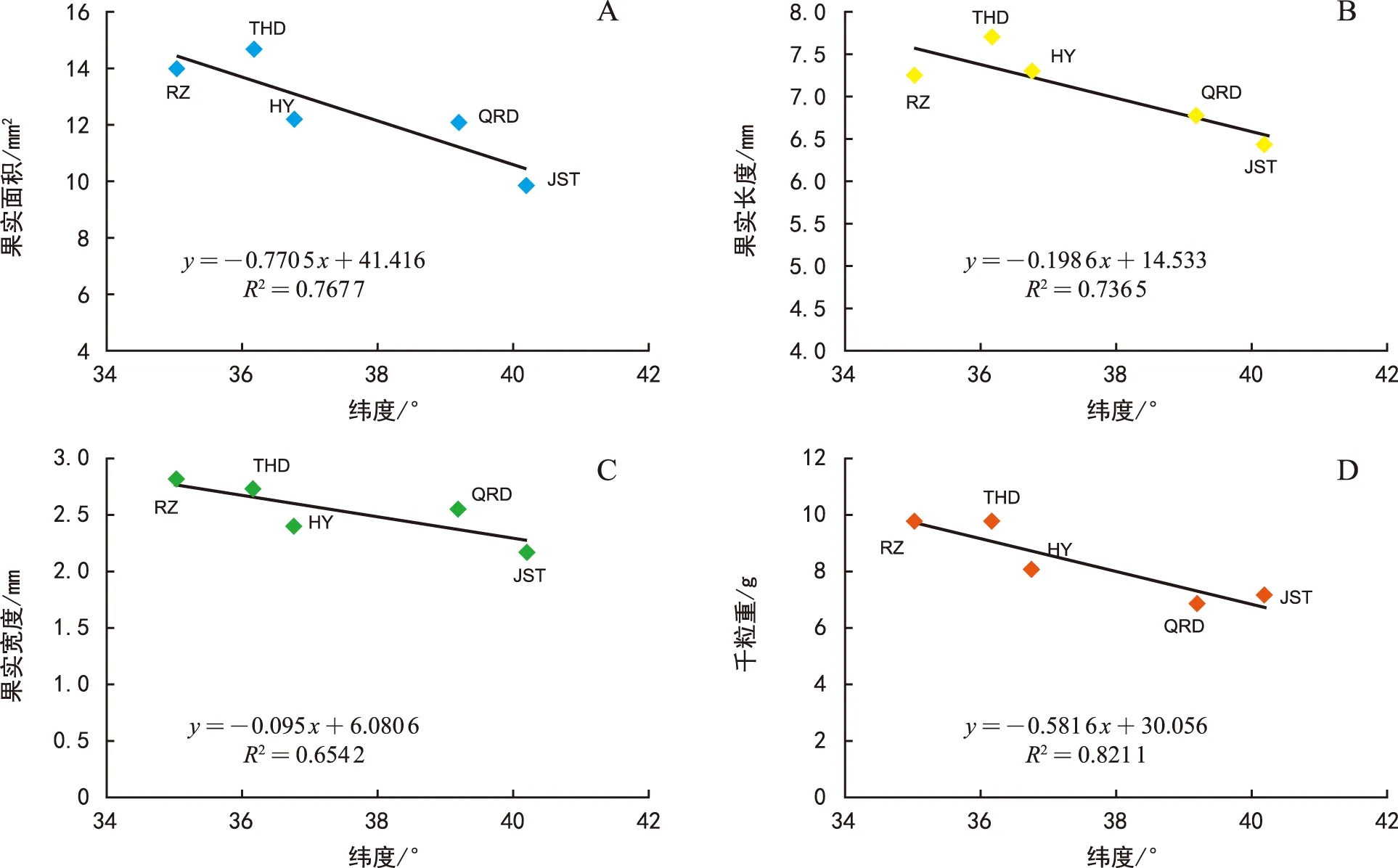
图2 不同种源北沙参种子特征与纬度的相关性分析Fig.2 Correlation analysis between seed characteristics and latitude of G. littoralis from different provenances
2.3 不同温度下各种源北沙参种子的发芽率差异分析
对不同种源北沙参种子在不同统计天数的发芽率数据进行单因素方差分析,由图3 A可知,在10 ℃处理下, 3 d时仅有金石滩极少量种子发芽;随着发芽时间延长,其他种源种子的发芽率缓慢增加,而金石滩种子发芽率增加较快,5~10 d统计的金石滩种子的发芽率均显著高于其他种子(p<0.05);至发芽13 d时,情人岛种子的发芽率亦稳定增加,与金石滩种子的发芽率愈来愈接近(p>0.05),而二者的发芽率明显高于其他种源种子的发芽率(p<0.05),16 d及19 d发芽统计结果同样如此(p<0.05)。
由图3 B可知,15 ℃处理下,各种源种子的3 d发芽率均很低,且无显著性差异(p>0.05);5 d时,金石滩种子的发芽率即显著高于其他种源种子的发芽率(p<0.05);至发芽7 d时,金石滩和情人岛种子的发芽率明显高于其他种源种子的发芽率(p<0.05),而二者之间差异并未达到显著水平(p>0.05)。统计至发芽10 d时,金石滩和情人岛种子的发芽率与其他种源发芽率均存在显著性差异,海阳种子发芽率则与田横岛及日照种源的发芽率亦达显著性差异(p<0.05)。
20 ℃处理下,各种源种子的3 d发芽率即表现出差异,其中,金石滩和情人岛种子的发芽率显著高于其他种源种子(p<0.05);5 d时,金石滩种子的发芽率明显增加,与其他种源种子的发芽率均存在显著性差异(p<0.05);统计至7 d时,金石滩和情人岛种子的发芽率明显高于其他种源发芽率(p<0.05),而二者之间
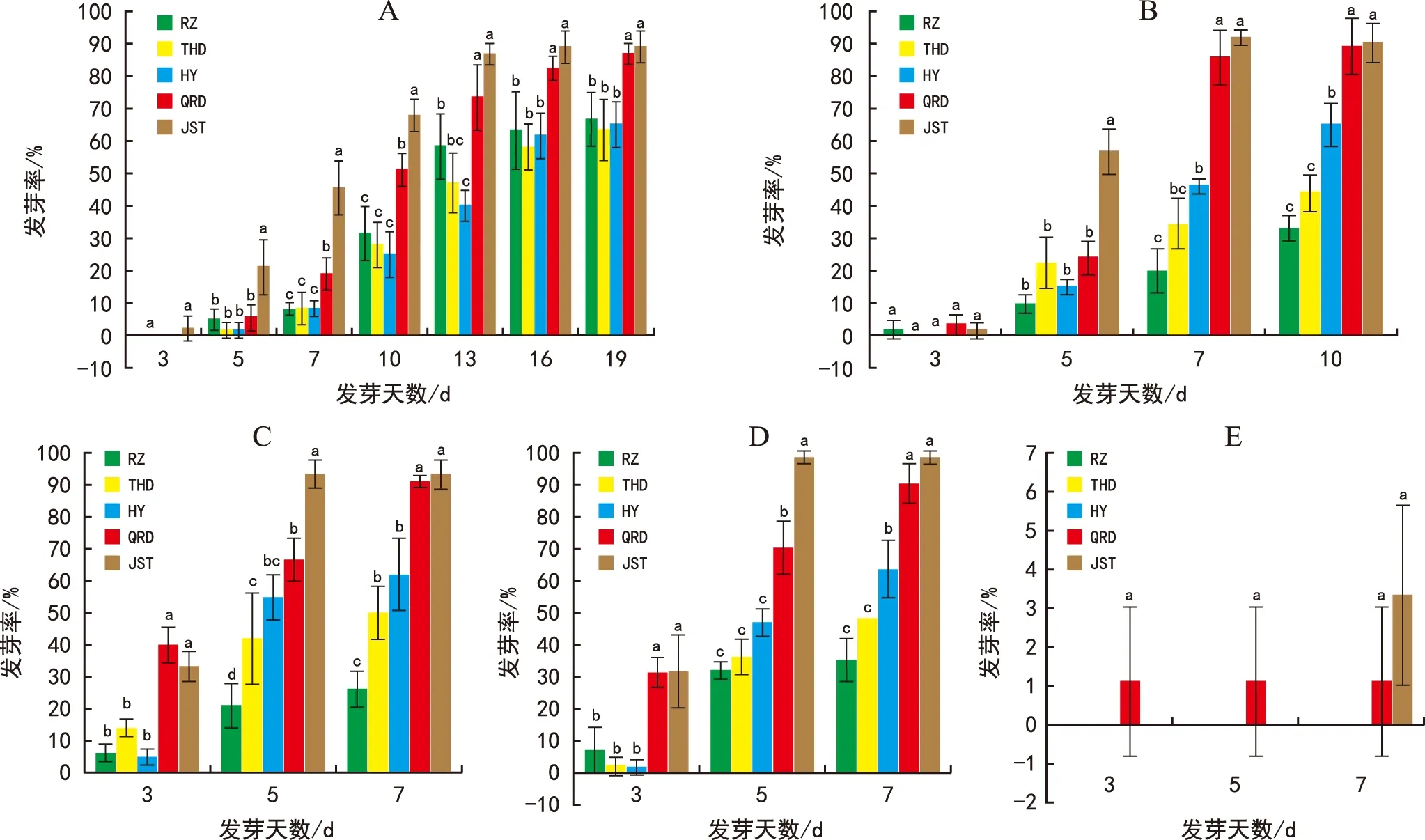
注:A为10 ℃;B为15 ℃;C为20 ℃;D为25 ℃;E为30 ℃。图3 不同种源北沙参种子在不同温度及不同发芽天数的发芽率 Fig.3 Comparison of germination rate of G. littoralis seeds from different provenances at different temperatures and germinating days
无显著性差异(p>0.05),日照种子发芽率最低,与其他种源种子发芽率均存在显著性差异(p<0.05)(图3 C)。
不同种源北沙参在25 ℃处理下随发芽天数的发芽率见图3 D。由图可知,各种源种子的3 d发芽率中,金石滩和情人岛种子的发芽率显著高于其他种源种子的发芽率(p<0.05);5 d时,金石滩种子的发芽率继续快速增加,和情人岛种子发芽率差异亦达到显著水平(p<0.05),而日照、海阳及田横岛种子的发芽率之间无显著性差异(p>0.05);7 d时,金石滩和情人岛种子的发芽率明显高于其他种源种子的发芽率(p<0.05),而二者之间差异未达到显著水平(p>0.05)。
30 ℃较高温度下,3 d与5 d时仅有情人岛的种子发芽且发芽率在5 d和7 d时未发生变化,7 d时金石滩种子的发芽率略高于情人岛种子的发芽率,但相互之间并无显著性差异(p>0.05)(图3 E)。
2.4 不同温度下各种源北沙参种子的最终发芽率及其与纬度的相关性
本研究的最终发芽率为10 ℃、15 ℃、20 ℃、25 ℃各处理下种子萌发试验结束时间对应的发芽率的平均值,用以综合直观的体现各温度下的发芽率。由图4可知,不同种源种子的最终发芽率与纬度呈显著正相关(p<0.05)。

图4 纬度与北沙参种子最终发芽率的相关性Fig.4 Correlation analysis of latitude and final germination rate of G. littoralis seeds
3 讨 论
近年来,野生北沙参资源量大幅缩减,濒危状况日益严重,1999年被《国家重点保护野生植物名录(第一批)》列为Ⅱ级濒危重点保护植物[14]。北沙参分布数量锐减的原因除了由于生境破坏、野生种群较小及分布生境狭阈等原因外,种子萌发率低也是重要原因之一[7]。因此,合理利用野生北沙参资源,根据种子发芽率挖掘优良潜力的种质具有积极的现实意义。研究发现,北沙参种子的发芽率较低,而北沙参后续相关研究进展缓慢,亦主要受限于北沙参种子的发芽率不高[15-16]。本研究也发现,不同种源北沙参种子的发芽率不同且与纬度呈显著正相关(p<0.05),比如日照种源的种子发芽率较低,但随着纬度的增加,北沙参种子的发芽率呈增加趋势。由此可见,北沙参种子的发芽率与种源的地理特征之间,特别是与纬度有一定的联系。在种子分级的各项指标中,发芽率最为重要,可以较为直接地反映种子的田间出苗率[17]。因此,发芽率可以作为北沙参选种的重要依据,发芽率高的种源,即辽宁省瓦房店情人岛和葫芦岛金石滩两地的野生北沙参可作为具有潜力的优势种质加以重点关注。
本研究表明,不同种源北沙参果实和种子的形态指标存在一定的差异,低纬度种源如日照、田横岛的果实和种子的面积和质量均较大,高纬度种源如金石滩及情人岛的果实和种子的面积和质量均较小,即随着纬度的增加,北沙参种子的面积、长、宽和千粒重均呈减小的趋势。由此可见,北沙参种子的表型性状与纬度有一定的联系,但果实、种子与纬度相关性分析的结果表现不同。纬度与北沙参果实面积、长度和宽度均呈显著负相关(p<0.05),与千粒重呈极显著负相关 (p<0.01)。纬度则与种子面积、长度及宽度的相关性均未达到显著水平(p>0.05),仅与种子千粒重相关显著 (p<0.05)。可能是由于北沙参各种源间的差异,更多体现在果皮的大小及质量差异较大,所以导致剥掉后无法很好地体现各形态指标间与纬度的相关性。
温度对不同种源北沙参种子的发芽率均有较大影响。低温下各种源的北沙参种子萌发较为缓慢,因此15 ℃统计至10 d,而最低温度处理10 ℃统计至19 d后连续3 d未有新种子萌发,发芽率才达到恒定不变,且在低温胁迫下胚根生长极其缓慢,和正常温度下生长的状态具有较大差别。发芽速度较快、发芽率最高的温度为20 ℃和25 ℃,而在30 ℃下各种源种子均无法正常萌发。本研究结果与Picciau等[18]的研究结论较为一致。温度对北沙参种子萌发具有较大影响,又因为北沙参具有耐寒的特点[19],因此北沙参种子在萌发阶段温度可以略低,但必须要避免高温,尽量保持一个适宜的温度以保证较高的发芽率。
本研究发现,随着种子千粒重的不断增加,北沙参种子的发芽率不断明显降低,可能有以下几个原因:首先,北沙参种子属于深度休眠的类型,相关研究表明,北沙参的种子内含有大量的香豆素类成分,是北沙参主要的活性成分之一[20-21],包含多种化学物质,被视为伞形科中药特征性的成分,亦是最早一批被确定为诱导植物休眠的物质之一[22],可能会极大地抑制种子的萌发,因此可能因为种子千粒重较大的北沙参种源种子中香豆素类成分含量较千粒重小的种源高,所以导致种子萌发被极大地抑制,本实验室同时开展的北沙参种子切掉一半胚乳的对比萌发实验为本研究结果提供了佐证(另文),不同种源北沙参香豆素典型成分的含量测定工作亦正在进行之中;其次,北沙参种子在储存期间的含水量高低会对后续的萌发产生较大的影响,在种子的含水量过高时,种子可能会发热、变质易发生霉变,因此千粒重较大的北沙参种源种子中的含水量较千粒重小的种源高,致使部分种子变质或失活,从而导致发芽率较低[23];日照及田横岛种源的种子出现较高的霉变率,尤其在较高温度(20 ℃及25 ℃)下,而在较低温度下霉变情况较为轻微;此外,因为地理隔离和复杂多样的气候、土壤、地形等生态因子的差异,可能会导致不同种源种子间在某些性状上产生了一定的遗传分化,促使不同种源发生了种内变异[24],但其具体机制仍需要进一步研究分析。
综上所述,不同种源北沙参的种子表型特征与纬度相关,相比而言,高纬度种源北沙参种子及果实的面积和质量较小,且其千粒重与纬度均呈显著负相关(p<0.05);北沙参种子发芽率与纬度呈显著正相关(p<0.05),萌发的适宜温度范围为20~25 ℃;辽宁省两地区的野生北沙参种源发芽率要优于山东省三地区的野生北沙参种源。此结果可为优质北沙参种源的选育选材提供有效参考。
猜你喜欢
绿色科技(2021年21期)2021-11-26
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
金桥(2021年9期)2021-11-02
食品安全导刊(2021年21期)2021-08-30
学苑创造·A版(2020年6期)2020-07-07
齐鲁周刊(2017年29期)2017-08-08
中学政史地·教学指导版(2016年12期)2017-05-16
绿色科技(2016年17期)2016-12-28
