
彭泽鲫♀×兴国红鲤♂杂交四倍体子代倍性鉴定及染色体核型分析
2021-07-15 12:13:48张庆飞郭青松虞炯莹操文杰王卫民
淡水渔业 2021年4期
张庆飞,郭青松,虞炯莹,操文杰,王卫民
(1.华中农业大学水产学院,农业动物遗传育种与繁育教育部重点实验室,农业农村部淡水生物繁育重点实验室,武汉 430070;2.武汉市江夏区农业综合执法大队,武汉 430200)
彭泽鲫(Carassiusauratusvar.pengsenensis)产于江西省九江市彭泽县,属于鲤形目(Cypriniformes)鲤科(Cyprinidae)鲫属(Carassius),具有生长快、个体大、抗病性及抗逆性强、营养丰富的特点,是淡水养殖的优良品种。彭泽鲫最初被认为是正常的二倍体(2n=166),营两性生殖[1],现已公认彭泽鲫为三倍体(3n=150+)鱼类,且生殖方式为两性型雌核发育[2]。兴国红鲤(Cyprinuscarpiouvar.singuonensis)产于江西省兴国县,属于鲤形目(Cypriniformes)鲤科(Cyprinidae)鲤属(Cyprinus),其背宽肉厚、肉质鲜美、生长快、食性广且抗逆性强,同时兴国红鲤杂交亲和力强,是重要的杂交亲本。
天然雌核发育的鱼类通常为三倍体或四倍体[3],在发育过程中精子只刺激卵子发育,本身并不融合,后代的生物学特性几乎与母本相同而不具有父本的遗传性状。目前已有研究表明,利用异精刺激天然雌核发育鱼类可产生正常的子代,但是不同种类的雄性精子产生的诱导效果差异显著。本实验室在兴国红鲤雄鱼精子刺激彭泽鲫雌核发育的子代中发现了四倍体,并于2020年5月进行扩繁得到子二代,推测该四倍体发生了两性融合,产生了远缘杂交四倍体。远缘杂交鱼类通常结合双亲的优势性状,具有较高的育种潜力,但能否稳定遗传仍存疑问。染色体作为遗传信息的载体,可以直观地了解其遗传变化和遗传特征,进一步判断子代的亲缘关系,因而对其子代进行倍性鉴定及染色体核型分析极为重要。
本研究通过对彭泽鲫♀×兴国红鲤♂杂交四倍体子代进行倍性鉴定和染色体核型分析,旨在为远缘杂交多倍体育种提供理论依据,探讨新品种的应用前景,同时也为原位杂交等研究提供数据支持。
1 材料与方法
1.1 实验材料
彭泽鲫亲本来源于江西省水产科学研究所,于华中农业大学南湖水产养殖基地进行产前培育,并在2020年5月挑选性腺成熟且健康无病的亲本进行人工繁殖得到子代;实验室前期在兴国红鲤精子刺激彭泽鲫卵子雌核发育的子代中发现四倍体,并于2020年5月挑选性腺成熟且健康无病的亲本进行繁殖得到健康子代。
1.2 实验方法
1.2.1 血液采集
用EDTA抗凝剂浸润一次性注射器内壁,采用尾静脉抽血的方式,从鱼体侧线下方扎针,碰到脊柱时向后轻拉针头,抽血1 mL置于2 mL离心管中,4 ℃保存,用于测定DNA相对含量,未加抗凝剂的血液直接用于血涂片的制备。
1.2.2 DNA相对含量测定
选用三倍体彭泽鲫血细胞作为内参,通过流式细胞仪检测倍性。取抗凝血剂10 μL置于2 mL离心管中,加入500 μL DAPI染液,再加入500 μL PBS缓冲液,用300目滤膜过滤,避光染色5 min后,上机检测。
1.2.3 血涂片制备
取一滴未加抗凝剂血液滴在光滑平整的载玻片上,将载玻片缓慢靠近血液,待血液散开后,以30°~40°的角度匀速向前推片形成血膜,待血膜完全晾干后进行染色。将Giemsa母液用PBS缓冲液稀释10倍后配置成工作液,滴加Giemsa染液染色1 min,再滴加等量的PBS缓冲液,染色30 min。30 min后流水冲洗,室温干燥。每种鱼制作10张血涂片,在Olympus正置显微镜下观察拍照,每张血涂片测量20个红细胞的长轴a和短轴b,根据公式表面积S=πab/4、体积V=a2b/1.91计算红细胞和细胞核的表面积与体积。
1.2.4 组织块原代细胞培养
取两种子代幼鱼并用75%酒精消毒体表,剪取鳍条及肌肉置于盛有AIM的培养皿中浸泡2 h,每半小时更换AIM,期间切除刮去多余组织。将组织块切碎成1 mm2小块均匀铺散在25 cm2细胞培养瓶中,用移液管从细胞培养瓶侧面缓慢加入4~5 mL原代培养基,置于28 ℃恒温培养箱中倒置6~8 h,翻转后静置培养。当细胞长满时,可消化传代,原瓶可保留继续生长细胞。
1.2.5 染色体标本制备
参考祝冬梅[4]染色体标本制备方法,取铺满细胞的细胞瓶,加入秋水仙素至终浓度为1.5 μg/mL,于28 ℃恒温培养箱处理3 h。吸出秋水仙素,用0.25%胰酶消化,再加入等量原代培养基,中止消化。将细胞混合液转入离心管,1 000 r/min离心7 min,弃上清液。冰水低渗10 min,1 000 r/min离心7 min。用卡诺固定液(甲醇 ∶冰乙酸=3 1,现配现用)固定2~3次,每次15 min,1 000 r/min离心7 min。吹打混匀细胞,滴片后自然风干,用Giemsa染液染色15 min,冲洗后自然晾干。
1.2.6 核型分析
用Olympus正置显微镜低倍镜查找分裂相,再用油镜拍照。选取126个分散良好、图像清晰、数目完整的中期分裂相统计染色体数目。并从中选取5个分散良好、形态清晰的中期分裂相,利用Photoshop、Image J软件放大图像并参考Levan等[5]提出的染色体类型划分标准对5个分裂相进行参数测量分析,染色体相对长度=(染色体长度/染色体组总长)×100;臂比=长臂/短臂,制作染色体核型图。
2 结果
2.1 红细胞DNA相对含量
以彭泽鲫红细胞为内参,利用流式细胞仪对杂交四倍体子代红细胞DNA相对含量进行测定,结果见图1。如图,横坐标表示红细胞荧光值,纵坐标表示细胞数目。彭泽鲫平均荧光值约为301.24±2.42,杂交四倍体子代荧光值约为405.79±3.18,二者DNA相对含量比为0.74,即杂交四倍体子代为四倍体。
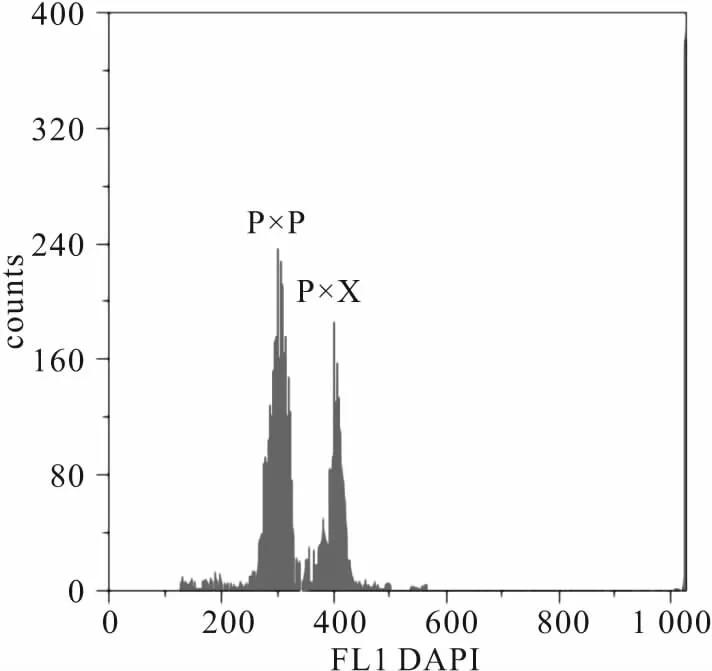
图1 两种子代红细胞DNA相对含量测量峰图
2.2 红细胞形态比较
彭泽鲫♀×兴国红鲤♂杂交四倍体子代、彭泽鲫雌核发育子代血涂片经Giemsa染液染色晾干后,在Olympus BX53正置显微镜油镜下观察红细胞形态,结果见图版1。如图所示,两种鱼红细胞经染色后细胞核颜色较深,核质分明。彭泽鲫雌核发育子代红细胞长轴略短于杂交四倍体子代,从形态上看杂交四倍体子代更长。由于倍性增加红细胞发生异形的概率增高,因而杂交四倍体子代血涂片中发现较高比例的异形红细胞,如哑铃型红细胞(黑色箭头)、无核红细胞(绿色箭头)、多核红细胞(蓝色箭头),而在彭泽鲫中异形红细胞较少。
2.3 红细胞大小比较
彭泽鲫雌核发育子代、彭泽鲫♀×兴国红鲤♂杂交四倍体子代红细胞测量结果见表1。结果显示:杂交四倍体子代红细胞长轴为彭泽鲫的1.13倍,差异极显著(P<0.01),但红细胞短轴与彭泽鲫差异不显著,四倍体子代红细胞表面积、体积均极显著地大于彭泽鲫(P<0.01),分别为彭泽鲫的1.08、1.22倍。因而在显微镜下可以观察到杂交四倍体子代红细胞更大、更长,彭泽鲫红细胞呈小且圆的椭圆形。在红细胞核方面,四倍体子代的长轴、核表面积、核体积极显著地大于彭泽鲫(P<0.01),分别为彭泽鲫的1.17、1.20、1.40倍,相同的是红细胞核短轴二者差异不显著。
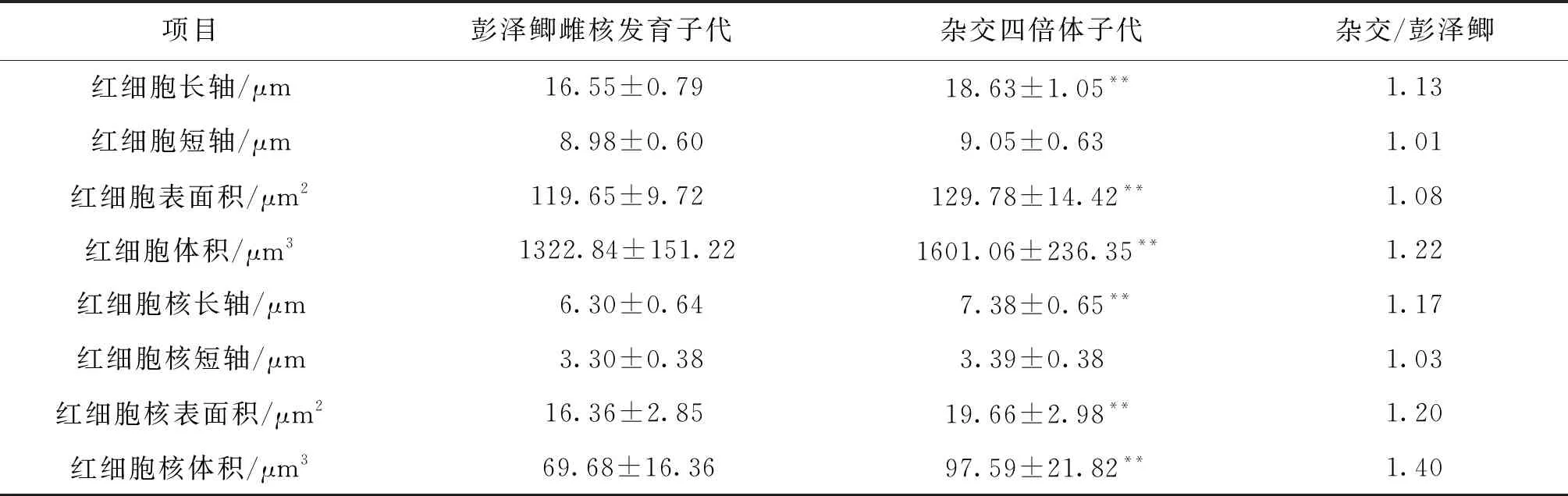
表1 彭泽鲫雌核发育子代、彭泽鲫♀×兴国红鲤♂杂交四倍体子代红细胞及核大小比较
2.4 肌肉、鳍条组织细胞培养
两种子代肌肉、鳍条组织块接种到T25培养瓶后生长情况见图版Ⅱ、Ⅲ。如图所示,两种子代肌肉、鳍条组织块贴壁情况较好,细胞围绕组织块呈发散状生长。彭泽鲫组织块3 d开始迁出细胞;7 d时在组织块周围有梭状细胞迅速生长;9 d时组织块之间细胞接触,生长迅速;15 d时,一些组织块周围细胞堆积变形,生长缓慢;18 d时基本铺满T25细胞瓶的80%,达到传代要求。杂交四倍体子代同样是3 d迁出细胞;5 d时已有组织块之间细胞接触生长;10 d时生长迅速;18 d时已经达到传代要求。传代结果见图版Ⅳ、Ⅴ,由于组织块周围细胞密集较难被胰酶消化下来,在传代过程中一些细胞被胰酶过度消化导致细胞死亡(黑色箭头),悬浮在培养基中。相比于原代培养,传代培养细胞增殖速度快,仅4~5 d就可以长满T25细胞培养瓶。截至目前,两种子代细胞已经传代到第10代。
图版Ⅳ 彭泽鲫子代肌肉、鳍条组织块传代培养(4×10)

图版Ⅱ 彭泽鲫雌核发育子代肌肉、鳍条组织块原代培养(4×10)
2.5 染色体数目统计
染色体数目统计结果见表2。经过观察统计,染色体数目在160以下有5个细胞;161~170有3个细胞;171~180的有9个细胞;181~190的有23个细胞;191~200的有15个细胞;大部分完整的中期分裂相染色体数目在201~210条,占55.56%。染色体数目分布范围广泛,但是在这个范围内并没有明显的众数。
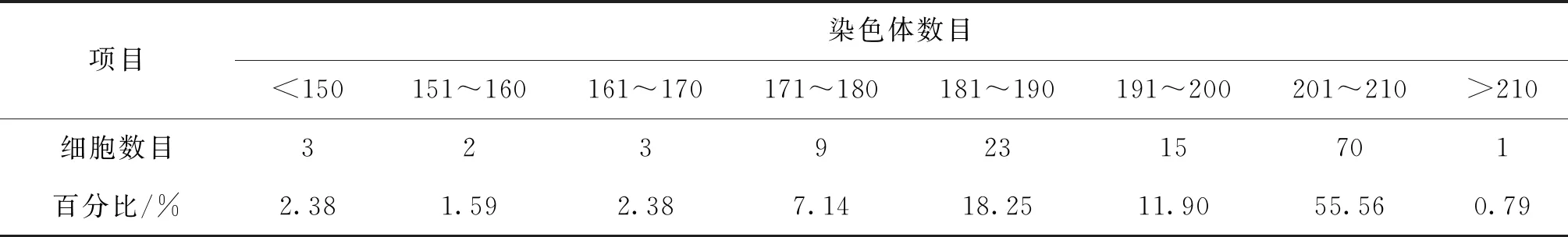
表2 彭泽鲫♀×兴国红鲤♂杂交四倍体子代染色体数目
2.6 染色体核型分析
按照Levan[6]的染色体划分标准,根据测量结果对染色体进行分型,结果见图版Ⅵ,其中包括42条中部着丝粒染色体(m)、64条亚中部着丝粒染色(sm)、62条亚端部着丝粒染色体(st)、32条端部着丝粒染色体(t),染色体臂数(NF)为368,其核型公式为4n=42m+64sm+62st+32t,NF=368,未发现异型性染色体。

图版Ⅵ 彭泽鲫♀×兴国红鲤♂杂交四倍体子代染色体
3 讨论
3.1 DNA相对含量与倍性
鱼类远缘杂交可以获得不同倍性的杂交后代[6],但是仅通过鱼类体表特征很难鉴定鱼类的倍性。常用的倍性鉴定方法有相对DNA含量测定法、红细胞体积测量法、染色体数目及核型分析法[7]。DNA作为主要的遗传信息,其含量与倍性呈正比关系[8-9],利用流式细胞仪测定其相对含量可以迅速、直接地反映鱼类染色体倍性,这种方法的优势在于对鱼体的伤害小、自动化、快速高通量且准确度高。目前,通过流式细胞仪技术鉴定倍性已经被广泛运用于动、植物中,如陈落落等[10]通过此方法鉴定出二倍体、三倍体虹鳟;方礼豹等[8]采用流式细胞计数法鉴定出二倍体、四倍体泥鳅及大鳞副泥鳅杂交四倍体子代的倍性;刘少军[6]通过此方法鉴定鲫鲂、鲫鲌等多种四倍体鱼类的倍性。另外,在马铃薯[11]、毛竹[12]等多种植物倍性鉴定上也广泛运用此技术。本研究中,将三倍体彭泽鲫作为内参,测定杂交四倍体子代红细胞相对DNA含量约为内参的1.33倍,即该杂交四倍体子代为四倍体,这一结果与桂建芳等[13]利用异源精子诱导异育银鲫雌核发育其子代可能存在四倍体的研究结果一致。本研究也表明,通过选取合适的内参,流式细胞术是一种快速、可靠鉴定倍性的方法。
3.2 红细胞形态、大小的比较
红细胞体积测量法也是辅助鉴定鱼类倍性的常用方法,其原理是细胞中染色体数目或DNA的含量与细胞核的大小呈正比[14],且细胞总是维持稳定的核质比[15]。这种方法的优势在于材料易得、易行直观。本研究中杂交四倍体子代红细胞长径略大于彭泽鲫,其形态相较于彭泽鲫雌核发育子代更长。同时我们在杂交四倍体子代中发现了许多哑铃状核、无核、多核等变异红细胞。这些异形现象在多倍体红细胞中是客观存在的[16]。同时在现有的鲫、草鱼、鲤、鳙等硬骨鱼类血涂片研究中都发现有红细胞分裂的现象[16],因而推测这种异形红细胞可能是由于红细胞分裂导致;也可能由于倍性的增加,部分红细胞难以维持稳定,导致细胞变形;此外有研究表明,饥饿胁迫也会影响红细胞形态,导致产生不规则形状或异形核红细胞[17]。但是这种现象在彭泽鲫中极少,因而这一异形现象也可以成为判断倍性的依据。本研究中杂交四倍体子代红细胞体积是彭泽鲫的1.22倍,核体积是彭泽鲫的1.40倍,但与预测理论值1.33不符。这一结果与陈落落等[10]、俞小牧等[18]研究结果相似。导致测量结果与理论值偏差与红细胞的生长受到许多因素的影响有关,如鱼类年龄大小、性别、生活环境、生长代谢情况等。同时本研究在同一张血涂片中也观察到了大小不同的红细胞,这是因为这些细胞处在不同的生长时期。与本研究不同的是,邹曙明等[19]在同源四倍体和三倍体团头鲂红细胞研究中发现其体积并未随倍性增加而增大。这表明不同鱼类红细胞大小是有差异的,单纯依靠红细胞测量法鉴定倍性可能不准确,因而结合上述流式细胞术对鱼体进行倍性鉴定更为可靠。
3.3 组织细胞培养及染色体标本的制备
鱼类的肌肉和鳍条组织具有极强的再生能力,迁出细胞的效果较好,又易于贴壁生长,是培养原代细胞的理想材料,同时染色体数目以及核型分析也是细胞遗传学的基础[5]。在本实验中,因为两种子代规格较小,取头肾等方法均不适用,因而选取了肌肉、鳍条组织进行细胞培养。根据培养结果发现,这两种组织迁出的细胞具有较强的增殖能力,通常3 d就可以达到再次传代的要求,目前可以传代到10代以上,但能否建立细胞系仍需进一步研究。此外,有研究表明,不同浓度的秋水仙素、低渗、固定液比例、滴片高度、冷热滴片法等因素会影响染色体的形态。在制备染色体标本时本研究发现提前12 h更换培养基会使细胞状态较好,染色体中期分裂相更多,形态更清晰;冰水低渗效果在15 min为宜,否则离心后细胞较少,但是随着传代次数的增加,细胞越来越难涨开,可能是由于细胞在多次传代后衰老,已经过了对数生长期,细胞状态不好;由于染色体数目较多,固定液最佳比例为甲醇 ∶冰醋酸=2 ∶1;离心速度不能超过1 000 r/min,否则细胞容易胀破;滴片高度要大于1 m。
3.4 染色体数目及核型分析
染色体是遗传信息的载体,是生命体进化的基础,同时染色体分析也是鉴定杂交四倍体子代倍性、遗传组成最直接、最准确的方法[20]。近年来,许多研究者对不同地区鲫以及杂交鲫进行了倍性鉴定和染色体核型分析的研究,结果见表3。
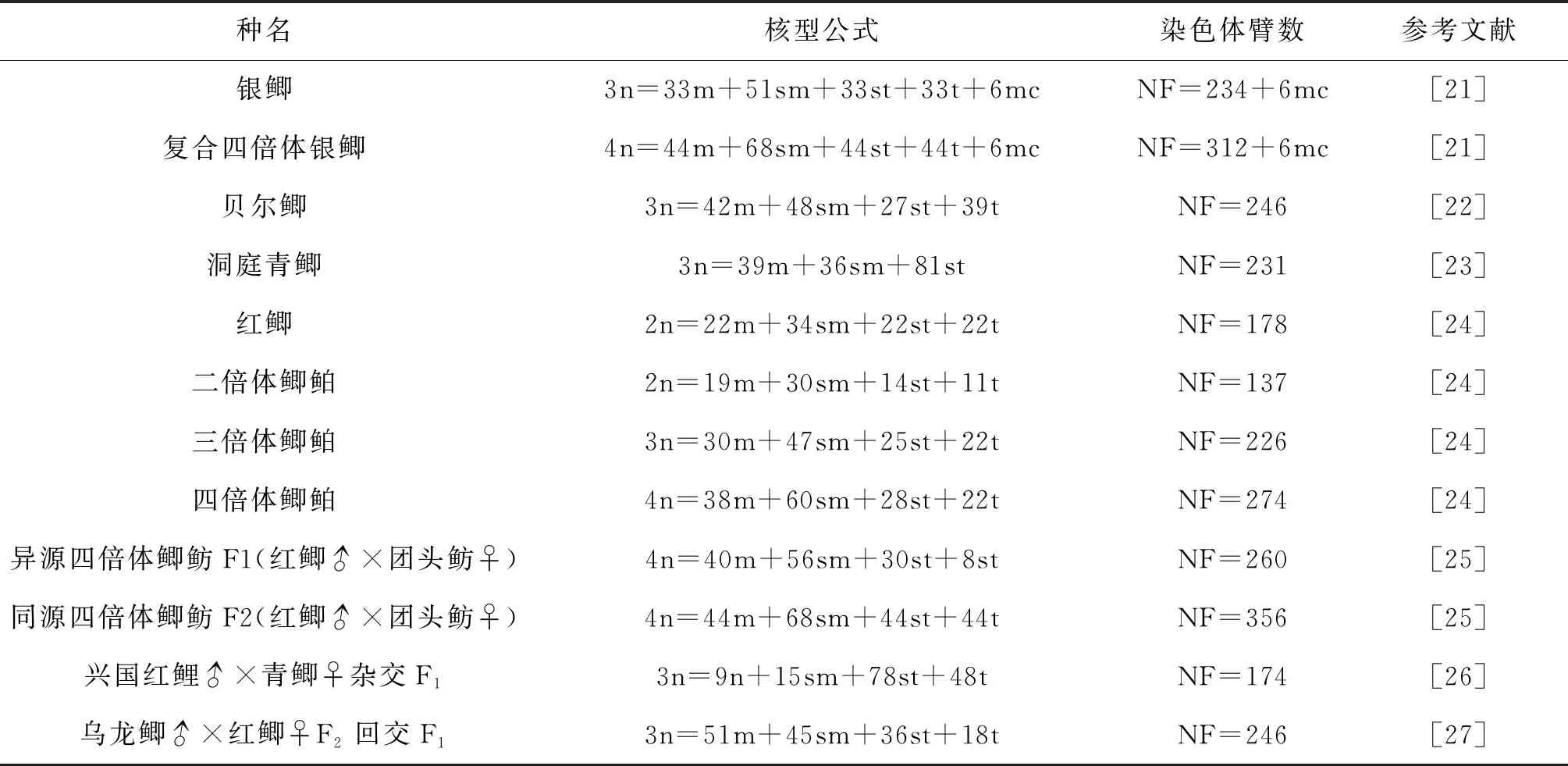
表3 不同地区鲫、杂交鲫核型分析结果
由表可知不同地区鲫以及杂交鲫的核型公式均有不同。例如贝尔鲫与银鲫均为三倍体但核型公式却不同;银鲫的染色体核型与野生鲫呈倍性关系;杂交鲫后代染色体核型公式会因父母本的不同而不同等,这些现象说明在鲫的进化过程中发生了染色体数目的加倍以及结构变异,同时这种核型的差异性也可能是因为地理种群的差异或研究方法、测量误差所致。本实验中,兴国红鲤是二倍体,核型公式为2n=28m+22sm+50st,NF=150;彭泽鲫是公认的三倍体,染色体数目为3n=150+,其核型公式为3n=33m+51sm+33st+33t[2]。本实验通过对126个染色体中期分裂相计数、测量分析得知:杂交四倍体子代的染色体数目分布范围较广,在150~210均有分布,其中200~210条占55.56%,但没有明显的众数。因而我们推测杂交四倍体子代的染色体继承了彭泽鲫的150+染色体以及本兴国红鲤配子的染色体,其核型公式为4n=42m+64sm+62st+32t,NF=368。造成分布范围广的原因可能是在制备染色体标本时细胞叠加导致染色体增加或者滴片时染色体丢失[28]。在鲫的染色体研究中并不缺乏染色体数目不确定的情况,如银鲫。有研究表明二倍体银鲫染色体数目为100;三倍体为150或由150条基本染色体和6条或8~12条超数染色体组成;四倍体则为206条[29-30],这些结果与本实验结果一致。但是我们发现杂交四倍体子代核型公式与彭泽鲫加兴国红鲤配子核型公式并不相同,这可能由以下原因导致:在细胞生长发育的过程中,染色体呈现周期性变化,尽管有丝分裂中期是最好的观察时期,但仍受外界环境影响,例如制备染色体过程中秋水仙素的浓度、培养温度等因素会导致染色体缩短,细胞状态、低渗液种类及处理时间、固定液比例、滴片高度、冷热滴片法等因素会影响染色体的形态;精卵结合后,染色体发生重组;实验方法的不同以及测量过程中的误差等[23]。
小岛吉雄[31]将真骨鱼类划分为低位类、中位类和高位类3个演化类群。从低位类群到高位类群,染色体越收敛,端部着丝粒染色体越多,染色体臂数越少。根据吴政安[32]研究结果:中部、亚中部着丝粒染色体较多属于低位类;反之,为高位类型。在本实验结果中,中部、亚中部染色体共101条,占50.5%,即彭泽鲫♀×兴国红鲤♂杂交四倍体子代属于中位类群。
4 小结
综上所述,采用三种方法对彭泽鲫♀×兴国红鲤♂杂交四倍体子代倍性鉴定结果为四倍体,其染色体核型公式为4n=42m+64sm+62st+32t,NF=368。理论上该杂交四倍体子代遗传了彭泽鲫和兴国红鲤的染色体,但仍需要对其染色体组成进一步分析。由于远缘杂交四倍体子代可能会体现出杂种优势,有关杂交四倍体子代在生长、形态、抗病等优势有待进一步研究。

图版Ⅲ 彭泽鲫♀×兴国红鲤♂杂交四倍体子代肌肉、鳍条组织块原代培养(4×10)
Plate Ⅲ Primary culture of muscle and fin tissue blocks ofC.auratusvar.pengsenensis×C.carpiovar.singuonensis(4×10)
1.2 d肌肉组织块贴壁;2.2 d鳍条组织快贴壁;3.3 d开始迁出细胞;4.5 d细胞生长迅速;5.10 d时组织块迁出细胞接触,生长迅速;6.15 d组织块周围细胞密集,变形;7.18 d细胞铺满培养瓶80%

图版Ⅴ 彭泽鲫♀×兴国红鲤♂杂交四倍体子代肌肉、鳍条组织块传代培养(4×10)
Plate ⅤC.auratusvar.pengsenensis×C.carpiovar.singuonensissubculture of muscle and fin tissue of hybrid progeny (4×10)
1.代传1 d;2.传代2 d;3.传代4 d;4.传代5 d;黑色箭头为死亡细胞
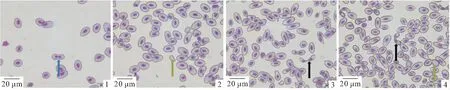
图版Ⅰ 彭泽鲫雌核发育子代、彭泽鲫♀×兴国红鲤♂杂交四倍体子代红细胞(100×10)
Plate I red blood cells (100 × 10) of inbred progenies ofC.auratusvar.pengsenensisand hybrid progeny ofC.auratusvar.pengsenensis×C.carpiovar.singuonensis
图1~2.彭泽鲫雌核发育子代红细胞图;3~4.彭泽鲫♀×兴国红鲤♂杂交四倍体子代红细胞图;蓝色箭头为多核红细胞;绿色箭头为无核红细胞;黑色箭头为哑铃型红细胞
猜你喜欢
High Technology Letters(2023年3期)2023-09-12 07:29:56
农业科技通讯(2023年1期)2023-02-12 07:09:08
心声歌刊(2022年4期)2022-12-16 07:11:00
丝绸(2022年5期)2022-05-25 18:02:26
Chinese Physics B(2022年3期)2022-03-12 07:48:34
Chinese Physics B(2021年11期)2021-11-23 07:28:34
音乐天地(音乐创作版)(2021年3期)2021-05-25 12:05:08
心声歌刊(2018年1期)2018-04-17 07:22:54
现代园艺(2017年13期)2018-01-19 02:28:17
中成药(2016年4期)2016-05-17 06:07:54
