
大兴安岭次生林区优势种落叶松分布格局及竞争作用
2021-07-12 00:07白小军谷会岩
生态学报 2021年10期
白小军,贾 琳,谷会岩,*
1 东北林业大学林学院, 哈尔滨 150040
2 陕西省神木中学, 榆林 719300
种群的空间格局是指种群内个体的空间分布方式或配置特点,是植物种群在群落中相对位置可以定量化描述的基本特征之一,能够反映种群在个体水平上彼此间的相互关系及组合方式[1- 4]。种群空间格局是在特定时间范围内种群相对静止的一种表现形式,一般可分为随机分布、均匀分布和集群分布3种类型[5]。
种群内个体的空间分布与形成该种群空间格局的生态学过程紧密相联[6],种群空间格局的形成既取决于种群自身的生物学特性,也与群落内不同物种之间的生物学关系以及群落内的环境等诸多其他因素有关,另外还与种群所处的发育阶段、 植被演替阶段、 以及研究尺度等密切相关[4,7]。因此对其空间分布格局研究不仅能够描绘植物个体分布格局,也有助于我们对产生这种分布模式的生态学过程如种内和种间竞争作用、种子扩散、干扰等的进一步理解、有利于我们阐明森林生态系统的形成与维持、群落的稳定性与演替规律[8]、有助于我们认识种群的生态学过程及其与生境条件的相互关系[9]、同时还可以揭示种群动态变化的格局以及成因、另外对研究群落中不同物种之间长期共存的维持机制也具有重要意义[10]、为植被及生物多样性的保护提供一定的理论依据[11]。
树种的空间分布和大小并不是相互独立的,而是受到种内、种间竞争过程影响[12],竞争是形成特定群落结构和种群分布格局的基本驱动力之一。森林中林木的竞争是一个普遍存在的自然现象[13],阐明森林群落中林木的竞争关系对于理解森林的结构和动态具有重要意义[14-15]。对森林群落竞争的研究, 广泛采用的是与距离和个体大小有关的竞争指数[16-17],检测群落中植物个体竞争作用的一个重要方法是使用空间点格局分析法对森林空间分布模式生成“快照”,将其二维特性保持不变,如果竞争在森林群落构造过程中是一个重要的工作机制,则相比于林木早期的演替阶段,林木的空间分布会随着时间变化而趋于均匀分布,例如前人在研究古田山中亚热带常绿阔叶林中的优势树种的分布格局时发现甜槠与木荷在空间上主要呈聚集分布,且随年龄阶段的增加,聚集程度有降低的趋势[10]。
理解竞争对植物空间分布格局的贡献和其导致的变化是一项重要挑战,有时竞争的强度、空间异质性或斑块性会导致空间格局分布模式与竞争间的关系模糊不清,这时则需要采用更加灵敏的方法来检测,而标记点格局分析能够使用反映树木生长情况的一些连续的附加信息(胸径、树高、冠幅和生长量等),并结合个体间的距离来分析群落内的竞争[8,18],相比于单一依靠树木间距离进行竞争作用的检测,标记点格局分析能够检测到森林群落内细微的竞争关系,可在植物个体空间分布上呈现出更加丰富的生长信息,这对于研究植物种群特定空间格局的形成机制和演替中相邻个体间的竞争非常重要。
优势种群的空间格局是群落结构的重要特征,既是群落结构、分布格局的体现,也是种群和群落动态与稳定性的标志[19]。目前, 天然次生林群落的次生演替动态受到了研究者的极大关注, 现已开展的研究主要包括次生林群落结构及优势种群的空间格局分析[20],但是目前对次生林恢复过程中建群种的种内竞争关系的定量研究比较少见,本文采用胸径为标记属性的点格局分析,分析种内不同龄级的个体间以及同一龄级的不同个体间的竞争关系,从而为揭示大兴安岭次生林群落结构、分布格局的形成与维持机制提供一定的理论基础,同时也希望能够为大兴安岭地区落叶松种群的保护和合理利用提供理论依据。
1 材料和方法
1.1 研究区概况
大兴安岭平均海拔1200—1300m,最高峰达2035m。山脉北段较宽,达306km,南段仅宽97km。形成于侏罗纪造山运动时期,沿东侧的走向断层掀升翘起,整个山体呈倾斜断块,造成东西两坡的斜度不对称。东面坡度较陡,西坡则向蒙古高原和缓倾斜,海拔790—1000m。大兴安岭大部为火成岩,地形平滑,山顶浑圆,山坡较平缓。山脉东坡被嫩江及松花江的许多支流深深地切割。夏季海洋季风受阻于山地东坡,东坡降水多,西坡干旱,二者呈明显的对比,但整个山区的气候比较湿润,年降水500mm以上。山脉北段是中国东部地区最冷之地,冬季严寒(平均气温-28℃),有大面积多年冻土区。山脉中段与南段温暖干燥得多,1月气温约-21℃,年降水量250—300mm,雪量也较少。北段的针叶林在南段逐渐转变成阔叶林,最后是点缀于林地之间的草场。 本文所研究的样地属于1987年火灾后的天然次生林,物种组成较为简单,群落结构也较为明显。
1.2 研究方法
1.2.1样地的建立
2014年7—10月,在大兴安岭呼中林区,经过实地查看,选择一块几乎无人为因素干扰经林火后恢复的天然次生林地带,设置一块3.24hm2(180m×180m)长期固定检测样地,采用全站仪(TopconGTS- 336)将整个样地按相邻格子法划分为81个面积为20m×20m的样方,再将每一样方拉绳划为4个10m×10m的小样方,以东西方向为X轴,南北方向为Y轴,以每个样方的左下角为坐标原点,检尺记录每一调查乔木个体在各个小样方中的具体坐标(坐标值精确到0.1m),同时记录样方中出现的所有乔木物种名、对每棵乔木进行检尺记录胸径、树高、并在每一调查乔木个体胸径处挂铝牌以用长期研究,进行格局分析时将上述调查到的植物个体点坐标换算为其在整个样地内的相对坐标。
1.2.2数据处理与层次划分
使用Excel进行数据的初步处理,然后用统计软件R(version3.1.0)进行点空间格局分析、空间相关性、种内竞争作用检测等分析;径级划分根据样地实际调查数据划分为幼龄(下层)0
1.2.3空间格局的分析方法
(1)基于距离的点格局分析
用R软件中Spatstat程序包中的最近邻体距离G(r)函数、双关联函数g(r)和基于个体胸径的标记点格局分析Mark函数对调查到的植物种进行了点格局分布分析,相关参考公式如下:
G(r)=1-e(-λ×π×γ2)
式中,A为样方面积,λ为模型估计参数,指样方内物种个体的密度或单位面积内期望点数,可以用n/A来估计,n为总点数(植物个体数),wij为边界效应修正,dij指两随机点之间的距离,I为指数函数,当dij≤γ时,I=1.0,当dij≥γ时,I=0;PCF函数是从k函数中衍生出来的,主要以环代替k函数中的圆,计算过程没有累积过程,当g(r)=1时,个体分布显示完全随机分布,当g(r)>1时为聚集分布,相反呈均匀分布。
(2)基于距离和个体属性的标记的点格局
经典的空间点格局分析方法仅基于距离来描述树种个体间的空间结构,然而,树木个体的胸径、树高、冠幅和生长量等附加信息,在空间上往往存在相关关系,例如在个体间相互抑制的情况下距离较近的植物个体的大小会低于整个群落的平均水平[21],所以对于同一种类型的植物群落来说,单变量标记点格局分析可以用来检测种内竞争。本文以的胸径标记属性,采用单变量标记点格局分析方法研究种群内植物个体空间分布格局关系,使用k函数进行计算,其可以用来探索因为密度效应引起的植物个体在生长上的降低[22],公式如下:
式中,t(mi,mj)是植物个体的标记mi和mj的假设检验函数,对于单变量分析,mi和mj分别是同一物种(或同层)两植物个体的胸径和树高。对于单变量标记点格局分析,当kmi,mi(r) =1时,距离为r的物种i和j的标记之间没有相关关系;当kmi,mi(r)>1时,两者呈现正相关关系;当kmi,mi(r)<1时,标记之间为负相关关系,即距离为r的两植物个体i和j的胸径或树高的大小低于整个群落的平均水平,其结果可以用来检测种内不同个体间的竞争关系。测试结果进行Monte-Carlo 拟合检验计算上下包迹线,即置信区间 拟合检验。用种群实际分布数据计算得到不同尺度下的g(r)值以生成置信区间,若检测结果在包迹线之间为随机分布,高于上包迹线为显著聚集性分布,低于下包迹线为显著均匀分布。本文通过99 次 Monte Carlo 随机模拟计算99%的置信区间。
2 结果与分析
2.1 不同龄级个体基于距离的分布格局及关联性
幼龄个体在0—5m的研究尺度下成显著的聚集分布,随着研究尺度的增加逐渐趋于随机分布格局;成龄个体在0—4m的小尺度下也呈聚集分布格局,但聚集强度较幼龄个体有所减弱;落叶松幼龄个体聚集分布在成龄个体的周围并与成龄个体有着相近的聚集尺度(图1、2);上层老龄个体在整个研究尺度下几乎都属于随机分布格局(图1、2);整体上来看落叶松从幼龄到成龄、老龄,随着龄级的增加,分布格局由聚集分布逐渐趋于随机分布;基于个体间距离的关联性分析显示:老龄与成龄个体之间在整个研究尺度下都没有关联、老龄与幼龄个体在0—3m的研究尺度下有一定的正相关关系、成龄与幼龄个体在0—5m的尺度下显著正相关(图5)。

图1 落叶松所有个体空间分布
2.2 基于胸径标记同一龄级个体间的竞争格局
基于胸径的单变量标记点格局分析发现,落叶松种群内存在明显的竞争(图3),上层的老龄个体在0—8m 和10—13m的尺度下存在显著的竞争作用(图3),这与张昊楠等人的上层个体检测不到显著的竞争作用这一结论相反[23];中层成龄个体在20—28m的研究尺度下出现了显著负相关关系,即中层个体只在较大的尺度下存在一定潜在的竞争作用(图3);下层幼龄个体在小尺度下未检测到这种竞争作用(图3),而在22—33m的尺度下幼龄个体也出现了竞争,下层幼龄个体出现竞争作用时的尺度与中层个体出现竞争作用的尺度是相接近的(图3)。

表1 样地内调查植物生物指标统计

表2 落叶松生活史的划分

图3 基于胸径标记的点格局
2.3 不同龄级个体间的竞争格局
2.3.1基于胸径标记的竞争格局
基于胸径的双变量标记点格局分析发现,落叶松种内上层老龄个体和中层成龄个体间在整个研究尺度下基本都呈现显著的负相关关系,说明在整个研究尺度上层老龄个体对中层成龄个体的生长存在显著影响即老龄个体与成龄个体间存在显著的竞争作用;上层老龄个体在较小(0—8m)的研究尺度下对下层幼龄个体也存在显著的抑制作用,从而显著影响下层幼龄个体的生长;在较小的研究尺度下中层成龄个体对下层幼龄个体基本没有影响,而在r>20m的尺度下,中层成龄与下层幼龄个体之间也出现了显著的竞争作用从而影响下层幼龄个体的生长(图4)。

图4 三种高度层次(上层,中层和下层)/三种龄级个体间(老龄、成龄、幼龄)基于胸径(DBH)的双变量标记点格局分析
2.3.2基于距离标记的竞争格局
基于距离的双变量点格局分析(图5)显示:不同龄级的老龄个体与成龄和幼龄个体间并没有检测到明显的负相关性即基于距离标记的点格局没有发现老龄个体与成龄和幼龄个体间存在明显的竞争作用;成龄个体和幼龄个体在小尺度下呈显著的正相关关系即成龄个体与幼龄个体之间不存在竞争作用,这一点与基于胸径标记的竞争格局分析结果相同;基于距离的先决条件零模型(图6)的分析显示:在0—2m和3—10m的尺度下,成龄个体对幼龄个体有显著的庇护作用即在小尺度下成龄与幼龄个体之间并不存在明显的竞争作用,这也与基于胸径标记的点格局分析结果相同。

图5 基于距离的完全随机模型(CRS)下不同龄级个体相关性

图6 落叶松幼龄成龄个体的分布格局
3 结论与讨论
很多研究表明,森林中较为高大的乔木上层个体即老龄个体几乎在所有尺度下均呈现随机或者均匀分布格局,这其中的一些原因被解释为是由于上层老龄个体间对来自于光照、水分和养分等资源的竞争作用所导致[10,24- 25],而较小径级的幼龄个体在小尺度下的聚集分布格局被认为是由于种子散布的限制、以及需要的资源较少、竞争相对较弱,从而导致小径级的幼龄个体更易出现聚集性分布格局[25]。本文的研究结果(图2)显示:落叶松老龄个体在整个研究尺度下几乎均属于随机分布格局、落叶松幼龄个体在0—5m较小研究尺度下成显著的聚集分布格局,但是随着研究尺度的增大,同样是幼龄个体其分布格局也发生着变化,这与上述前人的研究结论高度吻合[4,7,10,24];而对于形成这种分布格局的原因,结合前人的研究结论以及本文的研究结果来分析(图3),本文也认为老龄个体随机分布格局的形成与长期演替过程中随着龄级的增加老龄个体间出现显著的竞争作用从而导致老龄个体间的自疏即负密度制约效应有关[8,26- 27];而对于落叶松幼龄个体在小尺度下呈聚集分布格局的原因,根据本文(图2)的分析是由于受到种子扩散限制作用所导致的,这一结论也与前人通过大量研究得出的结论相同[8,24,27- 30],另据本文研究发现落叶松幼龄个体与成龄个体之间有着相近的聚集尺度(图2)、成龄个体对幼龄个体在较小的研究尺度下又有着较强的庇护作用(图6)、老龄个体与幼龄和成龄个体在小尺度下存在着显著的竞争作用、落叶松成龄个体与幼龄个体之间在小尺度下并没有检测到明显竞争关系(图4),且同龄级的幼龄个体间在小尺度下也未检测到竞争作用(图3),结合以上这些分析结果,本人认为幼龄个体的聚集分布格局的形成除了是受扩散限制作用的影响外与来自老龄个体的竞争排斥作用应该也有一定的关系,幼龄个体在与老龄个体之间在竞争过程中并不占优势,从而导致幼龄个体以及幼龄个体与成龄个体间的“抱团取暖”,形成所谓的聚集分布格局,这种分布格局形成有利于中下层成龄和幼龄个体在这种不利的竞争环境中生存[31];至于落叶松幼龄个体在较大的尺度下却又呈随机分布格局,应该与落叶松幼龄个体间以及成龄与幼龄个体间在较大尺度下出现的竞争作用有关(图3,图4)。
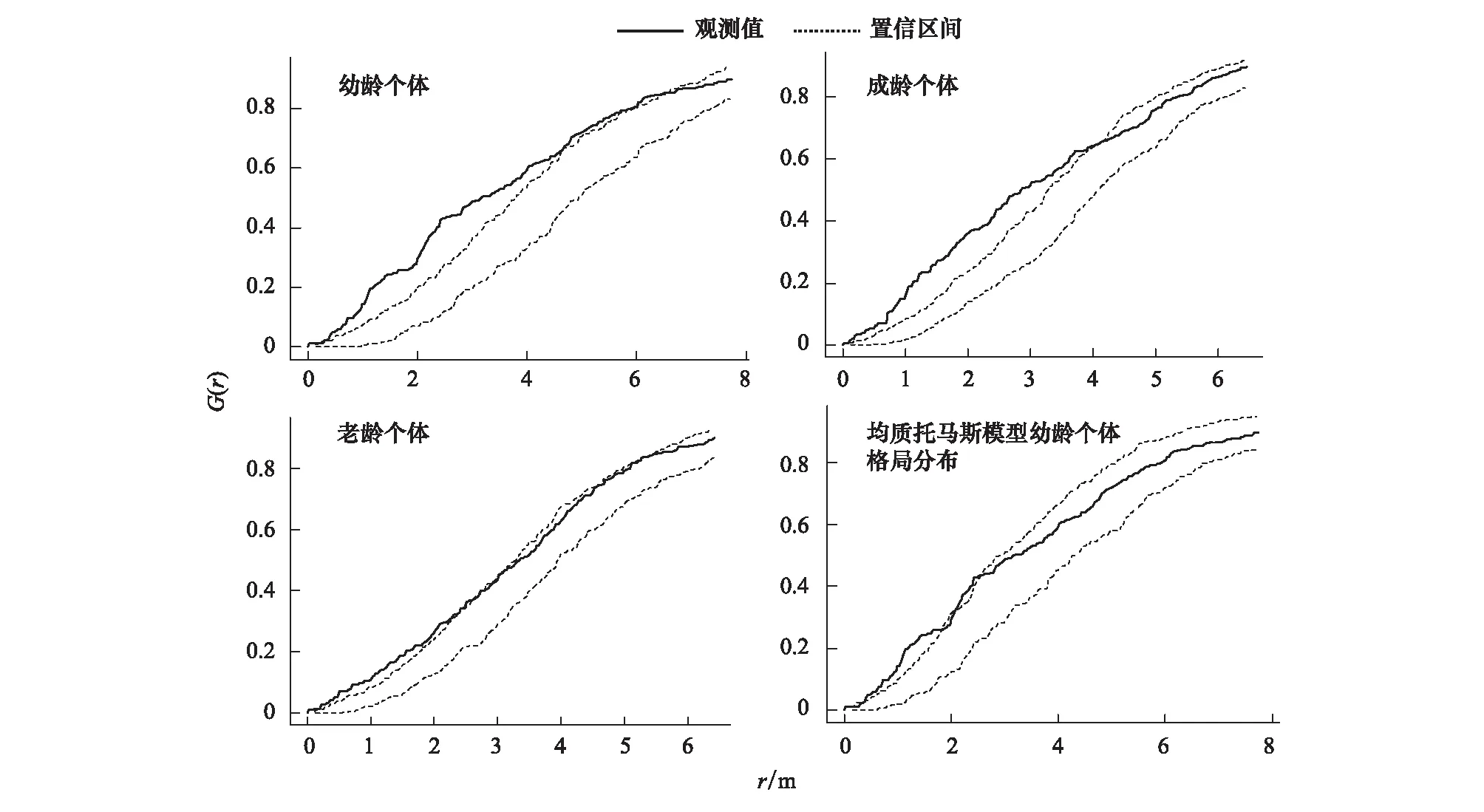
图2 完全随机模型下不同龄级个体的分布格局。
一般来说,由于物种的生态学和生物学特性、以及种子扩散限制和生境异质性等机制的影响,物种处在较小径级时容易呈聚集分布,尤其是在小的研究尺度上更加明显,而随着径级增大种内和种间为争夺资源的竞争也大大增强,聚集强度会随径级的增大而降低,呈随机或者规则性的分布格局,本文研究发现落叶松的空间分布格局及竞争作用随龄级的变化也符合这一规律[10,24],因此通过本文的研究,进一步证明了竞争作用在种群分布格局的形成过程中发挥着重要的作用;大尺度与环境异质性相关联[32- 33]、因此本研究中植物幼龄和成龄个体间在较大尺度下出现的竞争作用应该是由于环境的异质性导致的,这也说明明了植物所处环境条件的差异会影响植物间的竞争关系,而环境的异质性与生物学特性的复杂性又造就了群落内部物种的多样性和群落结构的差异性[34]。
基于距离的双变量点格局分析在分析不同龄级个体间的关联性时(图5)发现:不同龄级老龄个体与成龄和幼龄个体间、成龄与幼龄个体间并没有检测到明显的负相关性即基于距离标记的点格局发现老龄个体与成龄和幼龄个体间以及成龄与幼龄个体间并不存在明显的竞争作用[8],而基于胸径标记的点格局分析(图4)却检测到老龄个体与成龄个体和幼龄个体之间以及成龄与幼龄个体在较大尺度下间存在明显的竞争关系,这说明标记点格局分析要比单一基于距离的点格局分析在检测植物间的竞争关系时更为灵敏,能够检测到物种间或种内不同龄级个体间较微弱的竞争关系。
总之,种群分布格局是森林群落中不同物种之间相互作用的最直接反映,也是生物与环境之间关系的最直接反映[35];竞争作用是群落内不同物种维持共存的主要机制、从而影响着群落内物种的多样性[36,37],群落内优势物种及物种的多样性影响着群落演替的进程[38],群落在演替过程中能够形成怎样的结构、呈现怎样的分布格局以及群落的演替规律如何与植物间的竞争作用以及群落所处的环境条件等因素密切相关;老龄个体间在0—8m尺度下存在显著的竞争作用从而导致老龄个体的随机分布,幼龄个体与成龄个体在较大的尺度下也存在明显的竞争作用以及不同龄级个体间在特定的研究尺度下也存在竞争作用,进一步印证了森林群落中竞争作用是普遍存在的这一结论[13];植物幼龄个体在小尺度下的聚集分布不仅与种子的扩散限制作用有关[27- 30],也与竞争作用有关;植物个体的分布格局与个体所处的发育阶段有关,随着种群的发育,分布格局由聚集分布向随机分布格局过渡,与前人的研究结论相同[39],进一步印证了种群在发育过程中随着龄级的增加若分布格局由聚集格局向随机分布格局转变则说明种群内存在潜在的竞争作用这一结论的正确性[10],这也告诉我们从一定程度上可以通过检测群落内种间或种内竞争作用的大小来推断群落所处的演替阶段;在检测植物个体间的竞争关系时,相对于单一依靠距离的空间点格局分析,标记点格局分析能充分利用野外调查得到的植物胸径、树高等属性,因此能检测到植物间更细微的竞争作用、在检测植物间竞争作用时具有更高的灵敏性。
猜你喜欢
热带作物学报(2022年9期)2022-10-17
老龄科学研究(2022年9期)2022-10-12
烟台果树(2021年2期)2021-07-21
烟台果树(2021年2期)2021-07-21
中国动物保健(2020年6期)2020-11-27
今日农业(2019年13期)2019-08-12
人民调解(2019年5期)2019-03-17
中国果菜(2016年9期)2016-03-01
体育科研(2016年2期)2016-02-28
小说月刊(2015年12期)2015-04-23
