
荒漠草原向灌丛地转变过程中两种优势植物种子萌发及阈值特征
2021-07-12 01:39:58郭天斗孙忠超马彦平李志丽赵亚楠王红梅
生态学报 2021年10期
于 露,郭天斗,孙忠超,马彦平,李志丽,赵亚楠,王红梅,2,*
1 宁夏大学农学院, 银川 750021
2 西北退化生态系统恢复与重建教育部重点实验室, 银川 750021
土壤水分是干旱、半干旱地区主要限制因素,降水作为土壤水分的主要来源,对群落组成、物种生活及生理特性起主要调控作用[1]。据全球降雨模型预测,未来极端高温和降水发生更为频繁,出现“湿润区越发湿润,干旱区越发干旱”现象,特别是干旱地区因受全球气候变化和人为活动影响,降雨模式逐渐向降水频率降低、强度增大、总量减少等趋势变化[2- 4]。而草原灌丛化作为全球性生态问题,与全球气候变化联系紧密[5],作为一种“木本植物在密度、生物量及覆盖率的增加”现象,其扩张范围广、速度快,普遍发生在如美洲、非洲、亚洲等降雨量为200—850 mm的干旱半干旱地区,对植物、动物、土壤等均产生影响[6]。虽研究表明灌丛侵入增加土壤有机碳氮的积累[7],但在气候变化和人为因素的双重影响下造成一系列如土壤旱化、侵蚀加剧、草本多样性降低、碳氮去耦合化等草原功能弱化现象[8],均说明全球气候干旱趋势下的灌丛化是草原植物群落及土地退化的表现。相较于自然灌丛化,荒漠草原大量人为引入灌丛这一植被恢复措施实则加速了其灌丛化的进程。
而种子萌发过程是种子生活周期的关键阶段,作为植被更新过程中的瓶颈期,对环境变化高度敏感[9]。因此,确定灌丛化过程中限制植被更新的因素是理解荒漠草原灌木入侵植物优势种及植被变化的重要一步,特别是在灌丛和草原的交错区域,种子数量、种子萌发动态及早期定植均是植被更新的主要过程[8],而种子活力和萌发作为植被更新过程中的关键一步,调控着干旱区域内物种优势性、植被更新模式及速率[10- 12],受降雨与土壤水分影响较大。降水模式的改变可使土壤水分发生变化,并进一步影响着草地潜在量,有研究表明格兰马草(Boutelouaeriopoda)、冰草(Agropyroncristatum)等草原植物的萌发率和出苗率均随着降雨量增加而提高[13]。除降雨量影响作用外,生长季内的降雨强度、频率均会对种子萌发及植被更新过程产生显著影响[2,14]。土壤水分在一定范围内与种子萌发之间呈正相关[13],当含量到达一定阈值后方可打破干旱地区种子休眠[15]。干旱地区土壤水分除了受降雨的直接影响,还受到植被类型转变的间接影响,即使降雨量增加,仍会发生因植被蒸腾造成的生态干旱[5],如随着灌丛引入年限和密度的增加,加速土壤水分利用的同时又加剧深层土壤水分的消耗,出现地上草本生物量及丰富度均迅速下降,多年生草本土壤种子库密度呈下降趋势、萌发更新受到进一步抑制的现象[16]。因此,不定期且雨量减少的降雨模式变化会导致干旱区内的植被更新速率变缓,限制了当地草地恢复[11]。
宁夏东部荒漠草原地区自20世纪七八十年代起广泛应用旱生灌木锦鸡儿属柠条(Caraganakorshinskii)栽培种用于植被重建[17],在增加地上生物量的同时,多年生草本退化、土壤沙化、旱化等问题,荒漠草地正向人为灌丛地转变[18]。柠条锦鸡儿(Caraganakorshinskii)又名“柠条”,属于豆科植物,大多生长于荒漠半荒漠地区的固定、半固定沙地,具有防风固沙、保持水土等作用[19];沙芦草(Agropyronmongolicum)又名“蒙古冰草”,为典型多年生草本,广泛分布于宁夏荒漠草原区域[20],其耐旱耐风沙能力极强,但由于该地区大量柠条灌丛引入用以植被恢复,使得多年生草本沙芦草数量急剧递减,柠条锦鸡儿逐渐成为优势种。基于此,本研究选取宁夏东部荒漠草原-灌丛转变的镶嵌体区域的典型地段收集柠条锦鸡儿、沙芦草两种优势植物种子,开展实验室模拟干旱胁迫试验,探究全球气候降雨模式改变趋势下的干旱胁迫对其种子萌发影响,以期了解宁夏荒漠草原向灌丛地转变过程中的灌丛(柠条锦鸡儿)与多年生草本(沙芦草)在萌发阶段的竞争机制及对干旱的响应特征,进一步阐明全球气候变化背景下草原灌丛化的两种植被种子更新萌发竞争机制,为荒漠草原恢复和重建提供理论和依据。
1 材料与方法
1.1 种子收集区概况
研究区位于宁夏回族自治区盐池县中北部(37°5′—38°10′N,106°30′—107°39′E),与陕、甘、蒙三省区相临,北部与毛乌素沙地接壤,属于典型温带大陆性季风气候,草原类型属于荒漠草原,其植被主要以沙芦草、狗尾草(Setariaviridis)、猪毛菜(Salsolacollina)、赖草(Aneurolepidiumdasystachys)、画眉草(Eragrostispilosa)、柠条锦鸡儿、牛枝子(Lespedezapotaninii)为主;常年干旱少雨,且风大沙多,年均无霜期大约160 d,年降水量250—350 mm,雨季大多集中在7—9月,日照充足。土壤则以风沙土、灰钙土为主[18]。种子收集区主要位于研究区内四墩子野外观测基地(37°56′27″N,107°00′37″E)的大量柠条灌丛引入地段。
1.2 研究方法
1.2.1试验材料收集及处理
试验材料于2018年7—9月于宁夏盐池县四墩子基地荒漠草原试验区进行种子采集。供试沙芦草种子主要是由该试验区内野生沙芦草繁育基地采集,柠条锦鸡儿种子则通过野外随机收集方式获得,特征详见表1。将采集后的种子放置于4 ℃下贮藏供试验所用。试验前除去杂质并通过物理方法去除柠条锦鸡儿种皮,避免种皮过厚对试验有所影响。同时试验过程中为防止发生种子发霉现象,用0.52%HClO4消毒30—60 s后用蒸馏水反复冲洗。

表1 柠条锦鸡儿和沙芦草的种子特性
1.2.2适宜萌发条件下种子吸水及发芽模式
采用培养皿纸上发芽床,并将其放置于培养箱内恒温(25℃)、黑暗条件。共设置3个重复,每个重复内放置50粒种子,试验期间逐日统计发芽数,测定种子的发芽动态。另外,在培养6、12、18、24、30、36、42、48、54、60、66、72、78、84、90 h时,用以测定种子萌发所需水量。测定方法为取出全部种子(50粒)用滤纸吸干种皮表面水分并称重(精确至小数点后3位),以种子吸水后重量占吸水试验前重量的百分数表示[21]。
1.2.3种子胁迫处理
根据Michel[22]配制方法,采用PEG- 6000聚乙二醇对柠条锦鸡儿和沙芦草种子进行干旱胁迫处理,设置0(对照组CK)、-0.02、-0.1、-0.3、-0.6、-0.7、-0.9、-1.2 MPa(分别相当于2.9%、8.2%、12.5%、17.5%、20%、21.1%、23%、23.8%的PEG溶液浓度)共8个水势梯度,每一梯度设置3个重复,每个重复中各放置50粒大小一致、饱满种子,放置在双层滤纸上并盖上培养皿上盖以避免水分蒸发过快。准确称重后将所有处理放置在人工气候箱内(试验条件同1.2.2一致)进行培养15 d。每隔48 h采用恒重法向内注入蒸馏水来维持培养皿内溶液浓度,当胚根生长至4 mm左右时,可认为其萌发成功。每天观测并记录其萌发情况。评价种子活力的参数包括萌发率(Germination rate,G)、萌发速率(speed of germination,S)、累积萌发速率(Speed of Accumulation germination,AS)、发芽指数(Germination Index,GI)和活力指数(Vitality Index,VI),萌发起始时间(Initial Germination time,Gt)为种子胚根突破种皮1 mm时所需时间。
1.3 测试指标
1.3.1不同渗透势对种子萌发参数的影响测试指标[23-24]:
萌发率:
(1)
式中,n和N分别为萌发种子数和试验过程中所用种子总数。
萌发速率:
(2)
式中,N1,N2,N3,……,Nn分别指第1天、第2天,……,第n天的种子萌发率;
累积萌发速率:
(3)
式中,N1,N2,N3,……,Nn分别指第1天、第2天,……,第n天的种子萌发总数
萌发速率系数:
(4)
式中,N1,N2,N3,……,Nn分别指第1天、第2天,……,第n天的种子萌发数
发芽指数:
(5)
式中,Gt为时间t天的萌发种子数,Dvt为萌发试验天数。
活力指数:
VI=GI×S
(6)
式中,S为种子胚根长度。
1.3.2不同水势对种子萌发率的关系[8,24]:
(7)
(8)
式中,Gm为达到φ=0 MPa时最大萌发率,a为转折点时水势值,b为体型系数,φg为计算出的水势阈值。
1.4 数据分析
用数据统计分析中单因素方差分析的方法分析PEG浓度模拟干旱胁迫对种子萌发各项参数的影响,采用单因素方差分析中的最小显著差法(LSD)来分析各萌发参数在同一温度下不同水势浓度之间的差异;用回归法分析不同PEG溶液浓度与种子萌发系数间关系;利用S型生长曲线和线性模型来解释物种作为变量因子,水势作为协变量时的影响,物种和水势相互作用可以说明两种物种水势差别。数据处理采用SPSS 13.0软件,采用Excel和Origin绘图并进行曲线模拟。图表中萌发数据均以平均值±标准差表示。
2 结果
2.1 适宜条件下种子吸水特征
由图1可知,在同一环境下柠条锦鸡儿较沙芦草初始萌发时间较快,分别为42、66 h,此时柠条锦鸡儿和沙芦草吸水率各自为101.21%、238.77%。两者吸水速率存在差异,平均吸水速率分别为0.02 g/h、3.30 g/h,柠条锦鸡儿吸水速率和吸水量均低于沙芦草。

图1 柠条锦鸡儿和沙芦草种子萌发吸水动态特征
2.2 沙芦草、柠条锦鸡儿种子对不同PEG溶液的响应
由图2可知,随着PEG浓度逐渐升高,两种植物种子萌发率均呈先上升后降低趋势;而轻度干旱胁迫(φ=-0.02 MPa)下柠条锦鸡儿和沙芦草种子萌发率最高,分别为28.67%、44.67%;且均自-0.6 MPa起显著低于对照组(φ=0 MPa);在φ=-1.2 MPa时萌发率均最低,此时柠条锦鸡儿萌发率(为6.67%)大于沙芦草(为1.33%);沙芦草、柠条锦鸡儿种子萌发速率均在φ=-0.02 MPa时达到最大,分别为12.84%、9.52%;柠条锦鸡儿较沙芦草而言萌发较为整齐,出苗更快。

图2 种子萌发率、萌发速率对不同PEG溶液浓度的响应
随着萌发时间的延长(图3),柠条锦鸡儿和沙芦草萌发率呈增加趋势;在1—3 d内两者上升幅度最大,在5d后两者上升缓慢,其中以沙芦草发芽(0—-0.3 MPa)最为迅速;而在整个萌发周期中两者在0—-0.3 MPa内萌发率显著高于重度胁迫浓度(-0.6—-1.2 MPa);重度干旱胁迫下柠条锦鸡儿种子整体仍呈上升趋势,沙芦草则略有降低。

图3 萌发时间对萌发率的影响
2.3 萌发天数和胚根长度对不同PEG浓度的响应
由图4可知,随着PEG浓度逐渐升高,沙芦草和柠条锦鸡儿种子萌发初始天数均呈增加趋势,且差异显著(P<0.05);在0 MPa时两者萌发初始时间最短,此时柠条锦鸡儿为2 d,沙芦草为3 d;在渗透势最大(φ=-1.2 MPa)时,萌发天数分别为8、12 d,由此可得各渗透势下柠条锦鸡儿较沙芦草而言萌发较快;两者胚根长度随着渗透势的升高呈先增加后下降趋势,当φ=-0.02 MPa时柠条锦鸡儿和沙芦草的胚根长度最长,分别为1.69 cm和1.96 cm,并自-0.3 MPa开始呈递减趋势,并均在-1.2 MPa时最低(P<0.05),此时分别为0.82 cm、0.70 cm;而自φ=-0.7 MPa开始柠条锦鸡儿胚根长度长于沙芦草。

图4 种子萌发天数及胚根长度对不同PEG胁迫浓度的响应
2.4 种子萌发系数对不同PEG溶液的响应特征
随着渗透势逐渐升高(表2),柠条锦鸡儿和沙芦草累积萌发速率和活力指数均先上升后下降;在-1.2 MPa时两者指数则最低,沙芦草累积萌发速率自0 MPa起低于柠条锦鸡儿,活力指数亦呈现相同趋势。

表2 种子萌发对不同渗透势的响应
由图5线性分析所示,随着干旱胁迫不断加深,沙芦草和柠条锦鸡儿种子萌发率均呈下降趋势;当渗透势大于-0.8 MPa时,沙芦草种子萌发率高于柠条锦鸡儿;而萌发率和渗透势之间的线性回归方程分别为:柠条锦鸡儿为y1=-20.89x1+26.64(R2=0.71);沙芦草为y2=-36.50x2+39.93(R2=0.77)。
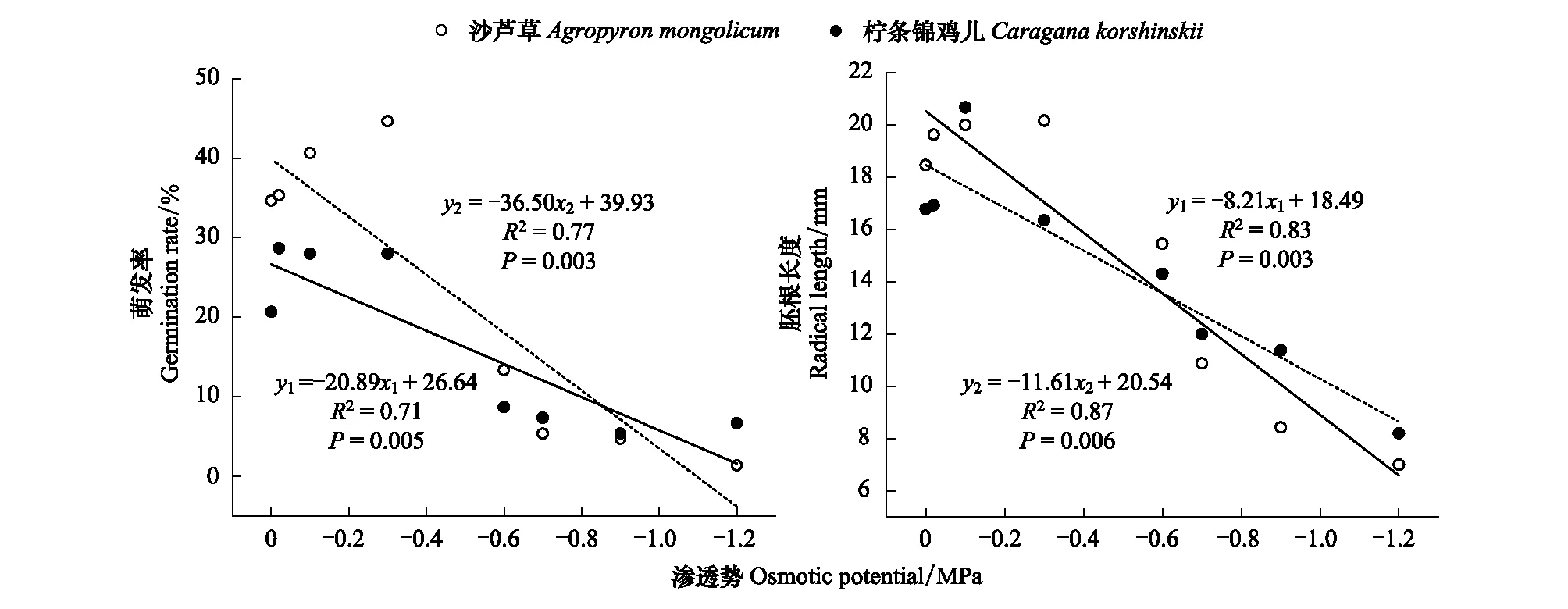
图5 萌发率与胚根长度与不同渗透势间的回归分析
胚根长度与不同渗透势之间的变化与其两者萌发率大体一致,均以沙芦草种子下降较为迅速,但与上述不同的是,在-0.6— -1.2 MPa内柠条锦鸡儿胚根长度长于沙芦草,此时两者线性方程分别为y1=-8.21x1+18.49(R2=0.83);y2=-11.61x2+20.54(R2=0.87)。由相关方程可知,柠条锦鸡儿种子比沙芦草种子更耐旱。
2.5 渗透势对萌发率的影响


图6 渗透势对萌发率的影响
3 讨论
植被更新过程作为植被物种多样性、群落结构的主要恢复过程,其生活型可以用来预测植被对干扰的响应[8]。而在干旱半干旱地区,土壤水分的消耗和竞争成为该地区灌木植物与草本植物之间优势竞争转变的关键原因,对于萌发和休眠起到重要作用[25]。本研究结果中沙芦草种子萌发时所需水量明显高于柠条锦鸡儿种子,但萌发率却低于柠条锦鸡儿,这与曾彦军[21]等人研究结果相一致,说明沙芦草萌发更需要相对湿润的土壤条件。本研究中不同渗透势对宁夏灌丛引入的荒漠草原区的两种优势植物种子萌发有一定影响,其萌发率、萌发速率及胚根长度等均随着浓度的增加呈递减趋势,这与前人研究一致[26-28],其中轻度干旱胁迫可促进种子萌发这一结果也与Zhu等[14]、杨景宁等[29]结果一致,说明荒漠草原区植物在其发育过程中已形成相对适应干旱的机制。而种子萌发后能否定植成功,则依赖于所生存环境中的土壤水分状况以及植物根系发育和获取水分能力[29]。在同一环境下根系较深的幼苗更容易定植存活,在-1.2 MPa(渗透势最大)时,柠条锦鸡儿种子萌发率及胚根长度均高于沙芦草,且各梯度下的萌发天数均短于沙芦草,说明柠条锦鸡儿种子耐旱性较强,出苗速度快,能够充分利用周围环境进行生长发育;沙芦草在0—-0.6 MPa内萌发率较高,但随着渗透势不断增加,其萌发率递减幅度增大,表明水分降低种子吸水困难,导致萌发率降低,抑制了种子萌发;同时又因种子胚根部位对重度干旱胁迫较为敏感[19],主根发育受到干旱影响从而伸长缓慢,在极端干旱环境下沙芦草幼根无法从深层土壤中获取水分等营养资源,难以支撑植物体生长,从而失去萌发、存活以及生殖机会。
土壤水分匮缺使得干旱、半干旱地区进行有性生殖的植物在萌发时受到限制,从而难以实现植被更新。研究表明同一环境下灌木与多年生草本之间的主要竞争与土壤资源(以水分为主)有关[30];多年生草本因根系较浅,其盖度及生物量随着土壤水分增加呈线性递增趋势;而灌丛具有“双峰”及发达根系,并以无性生殖进行扩张[31],能够充分利用深层或表层0—30 cm土壤水分[32],这一特性使其能存活于水分匮乏的区域,而本结果中柠条锦鸡儿水势阈值范围(-0.3—-0.86 MPa)低于沙芦草(-0.3—-0.65 MPa)说明灌木种相较于多年生草本更易在营养较低、更为贫瘠的环境中生殖发育。除上述因素,种子休眠作为植物适应理想环境下延迟萌发的一种生理机制,通过延长萌发和出苗速率来调控种子萌发,也是影响灌丛化过程中植被更新的一大因素[19],而萌发和休眠作为完全不同的两个阶段,气候变化也会分别对两者进行影响[9],从而影响植被更新过程。
尽管诸多原因造成灌丛化现象,但全球气候变化导致降雨模式逐渐发生变化,使得干旱、半干旱地区草本数量骤减,为耐旱、耐贫瘠的灌木植物提供扩张机会[33],对其群落结构和生态系统有着深远影响。最先体现在植被群落结构和空间分布的巨大转变[34]:短期使生物量发生变化,长期则影响植被构成[35];其次不定时降雨使土壤水分明显有所变化,使柠条锦鸡儿种子进入“吸水后脱水”这一循环,促进种子进入次生休眠阶段[36],提高了萌发率,使其有机会进入土壤种子库中为下一阶段的种群萌发更新提供来源及动力[37]。降雨不频繁加之蒸发量与降水量差距较大,使得土壤水分不断进行表层消耗,更易恶化土壤干旱化程度,降低了多年生草本种子萌发的水分条件。而本文中随着干旱胁迫加大,萌发速率及活力指数等均受到抑制,但柠条锦鸡儿显著高于沙芦草这一结果表示,在人为大量引入灌丛后,极强的竞争优势与降雨模式的改变,造成土壤表层水分的空间异质性,引起“水分负平衡”现象[17],使得干旱区内的植被更新速率变缓,限制了该区域草地恢复[11],又因灌木对周围资源(尤其水分)利用率较高,形成“沃岛效应”,更加速了生物多样性和土壤侵蚀趋势[38],导致土壤水分过度利用[39],不断消耗深层次土壤水分[40],使得土壤浅层春季返潮不明显,恶化了多年生草本的萌发土壤水分条件,进而限制了多年生草本对降雨的响应。一旦草原植被恢复力退化,便会加剧该地区荒漠化,促进了灌丛化现象发生[35]。以上种种均说明植物与气候之间存在着一种反馈机制,特别是人为灌丛引入加剧土壤水分利用和深层消耗,加之气候发展趋势有所改变,均增加了荒漠草原地区多年生草本萌发所需土壤水分条件的难度,植被变化、气候改变以及两者相互作用可能更加促进了灌丛扩张、多年生草本的退化[41-42]。
4 结论
通过对宁夏荒漠草原向灌丛地转变过程中的两种优势植物(柠条锦鸡儿、沙芦草)种子萌发应对干旱特性研究表明:轻度干旱胁迫(0—-0.3MPa)可促进两者胚根生长,重度干旱胁迫(-0.6—-1.2MPa)对两者均有抑制作用,柠条锦鸡儿萌发率、萌发速率均高于沙芦草,且萌发较早并出苗整齐;柠条锦鸡儿萌发水势阈值为-0.3—-0.86MPa,低于沙芦草水势阈值(-0.3—-0.65MPa),更具耐旱及萌发优势,在预测全球降雨降低的背景下,荒漠草原的柠条灌丛(种子)较多年生草本(种子)更适应土壤旱化,其旱生条件下的植被更新更易发生,本研究结果可为灌丛引入下的荒漠草原植被更新趋势做出预测,但就其植物种子更新周期还有待进一步的研究。
猜你喜欢
水土保持学报(2022年5期)2022-10-10 14:20:06
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01 11:26:42
东北石油大学学报(2021年1期)2021-03-25 07:03:40
现代养生·上半月(2020年11期)2020-11-13 03:44:37
防护林科技(2020年5期)2020-07-21 01:53:32
水土保持研究(2020年1期)2020-04-27 08:06:22
草业科学(2019年5期)2019-06-04 09:36:24
散文选刊·下半月(2019年2期)2019-03-15 06:32:46
西藏农业科技(2018年4期)2018-04-25 06:39:24
天津师范大学学报(自然科学版)(2016年4期)2016-12-14 06:04:26
