
发酵过程中不同理化因素对乳酸菌噬菌体的影响
2021-07-11 14:58:52于晓丽赵莉程溪秦松跃狄亚心崔文姜艳平王丽李一经唐丽杰徐义刚周晗乔薪瑗
中国乳品工业 2021年6期
于晓丽,赵莉,程溪,秦松跃,狄亚心,崔文,姜艳平,王丽,李一经,唐丽杰,徐义刚,周晗,乔薪瑗
(东北农业大学动物医学学院动物疾病防控技术与制剂创制实验室,哈尔滨150030)
0 引 言
乳酸菌作为益生菌,广泛应用于食品发酵及饲料加工业,在功能食品、医疗保健、微生态制剂等许多领域也具有较好的应用前景[1-4]。乳酸菌具有提高机体免疫力、调节肠道微生态平衡[5]、促进机体新陈代谢[6]等功能,对机体发挥着不容忽视的作用。乳酸菌定植肠道与自由基结合,可降低对机体的损害[7],且菌体及其提取物均具有抗氧化作用[8]。乳酸菌长期用于食品发酵,已被证明不具有致病性,被公认为安全级微生物[9]。但是乳杆菌在发酵过程中的不同阶段均受到噬菌体污染的威胁,导致发酵周期延缓、产量下降甚至全部浪费,造成严重的经济损失[10-11]。
在食品加工、化学制药、饲料生产和生物技术等行业,噬菌体污染问题被广泛报道[12]。减少或消除噬菌体污染,成为研究者非常关注的问题[13]。在发酵生产的不同阶段若发生噬菌体侵染,会导致乳酸菌产酸量减少,发酵过程缓慢或产品产量、质量下降等,造成巨大的经济损失[14]。Cox首次报道了噬菌体侵染发酵剂,在该领域取得显著进步,特别是在噬菌体遗传学、生态学及抵抗外界环境因素方面[15],随后又有相关文献[16]报道了干酪乳杆菌和副干酪乳杆菌的噬菌体导致乳品发酵失败;近来年,对噬菌体侵染酸奶和奶酪的报道[17-19]变得越发频繁,造成严重的经济损失。目前,噬菌体污染仍然是影响发酵工业的一个难题。
本研究通过分析环境中不同理化因素对3株乳酸菌噬菌体的影响,为控制发酵生产中噬菌体污染、保护工业菌株、制定有效防控措施提供理论依据。通过对发酵介质中理化因素对噬菌体及宿主菌的影响分析,为筛选抑制噬菌体活性且不影响发酵过程的有效措施奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌株
干酪乳杆菌(L.casei ATCC 393)戊糖乳杆菌(L.pentosus KLDS 1.0413)短小乳杆菌(L.brevis ATCC 367),由本实验室保存。噬菌体Lc、Lpen和Lbre;均由本实验室保存。
1.1.2 主要实验仪器与设备
垂直板电泳系统BIO-RADMV120型,Savant Instruments;低温台式高速离心机Z323,HERMAL公司;电热恒温培养箱DNP-9162型,上海精宏实验设备有限公司;电热恒温水槽DK-80型,上海精宏实验设备有限公司;凝胶成像仪UVR-800 Uitra-Uiolet Procducts Limted Cambhdge,UK吉尔森公司;微量电子天平BS201S型,北京赛多利斯公司;透射电子显微镜H-7650型,日本日立公司。
1.1.3 培养基配制
试验过程中使用的主要培养基见表1。
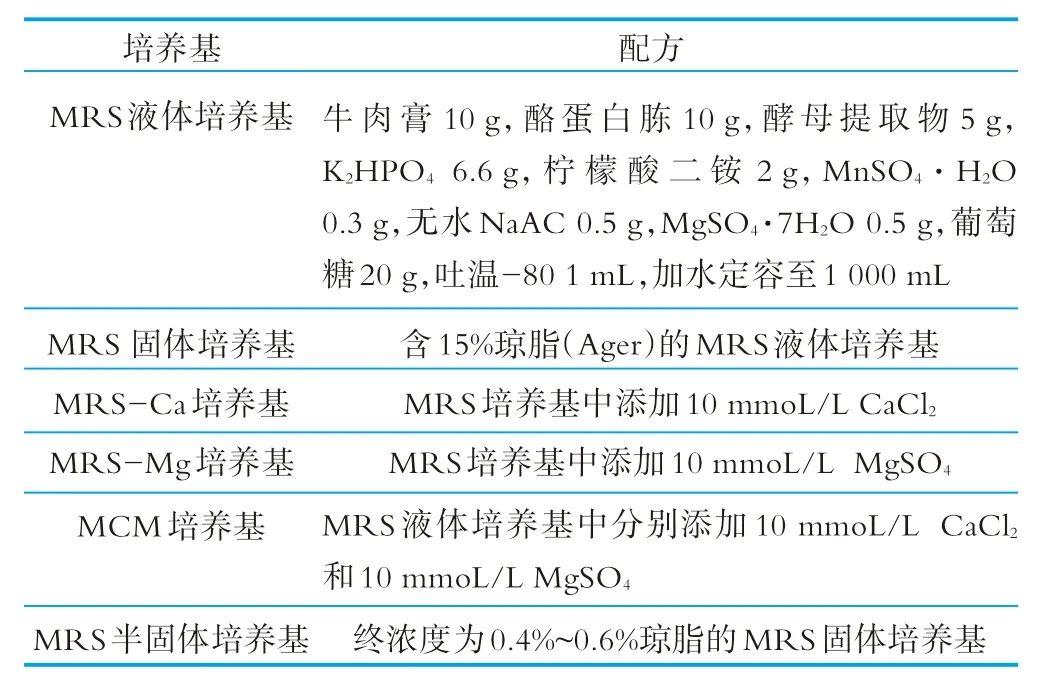
表1 主要培养基及其配方
1.2 试验方法
1.2.1 发酵过程中p H对宿主菌和噬菌体的影响
取不同p H值(p H=2~11)的MRS培养液98 m L,分别接种L.casei ATCC 393、L.pentosus KLDS 1.0413和L.brevis ATCC 367菌液2 mL,37℃静置培养18 h,测定OD600值。再将不同p H值(p H=2~14)的900μL MRS培养液中分别添加100μL噬菌体,混匀,32℃温育30 min,以L.casei ATCC 393为指示菌,以MRS固体培养基做下层培养基,将原液10倍比稀释后取100μL接种100μL指示菌,再将混合液接种4 mL MCM半固体培养基做上层培养基,制成双层琼脂平板,对形成的噬菌斑进行计数,测定噬菌体的效价,计算噬菌体存活率。实验重复3次。
1.2.2 发酵过程中温度对宿主菌和噬菌体的影响
为了减少发酵食品中乳酸菌菌种的使用、缩短发酵时间,对菌种最适生长温度进行测定,本试验选取发酵过程中常用温度,取MRS培养液98 mL,分别接种L.casei ATCC 393、L.pentosus KLDS 1.0413和L.brevis ATCC 367菌液2 mL,将其置于32、37、45、56℃条件下进行培养,每隔2 h测定其OD600值,绘制在不同温度下的生长曲线。为了测定噬菌体的热稳定性,筛选出利于乳酸菌发酵但不利于噬菌体生长的温度,参照Capra[20]等所描述的方法,用MRS培养宿主菌后,再分别感染3种噬菌体,噬菌体裂解宿主菌,收取裂解液,分别置于32、37、45、56℃和63℃温度下作用,于2、5、10、15、30、45 min和60 min分别取样,12 000×g离心4 min,取100μL上清进行十倍比梯度稀释,测定噬菌体效价。实验重复3次。
1.2.3 发酵介质中二价阳离子对宿主菌和噬菌体的影响
取MCM、MRS-Ca、MRS-Mg、MRS液体培养基98 mL,每一种培养液均分别接种L.casei ATCC 393、L.pentosus KLDS 1.0413和L.brevis ATCC 367菌液2 mL,置于37℃静置培养18 h后测定其OD600值。参照Quiberoni等[21]所使用的方法,用MRS培养噬菌体,获得噬菌体裂解液,采用双层平板法制斑,其中上层分别为MCM、MRS-Ca、MRS-Mg、MRS半固体培养基,下层均为MRS固体培养基。32℃静置培养,测定噬菌体的效价。
1.2.4 发酵介质中NaCl对宿主菌和噬菌体的影响
在MRS培养基中添加质量浓度1%、2%、3%、5%NaCl,取添加质量浓度1%、2%、3%、5%NaCl的MRS培养液各98 mL,分别接种L.casei ATCC 393、L.pentosus KLDS 1.0413和L.brevis ATCC 367菌液2 mL,37℃静置培养18 h,测定OD600值。参照Capra等[20]使用的方法,用MRS培养3种噬菌体,将终浓度为106PFU/m L噬菌体裂解液置于上述不同浓度NaCl发酵液中,32℃孵育4 h,测定噬菌体的效价。
1.2.5 发酵介质中聚山梨醇酯类(T-20)对宿主菌和噬菌体的影响
取MRS培养基添加T-20,体积分数分别为0.01%、1%、2%,将其平均分为两组,取添加积分数分别为0.01%、1%、2%T-20的MRS培养液各98 mL,分别接种L.casei ATCC 393、L.pentosus KLDS 1.0413和L.brevis ATCC 367菌液2 mL,第二组除接种等量菌液外,按最佳感染复数(motiplicity of infection MOI)接种对应的噬菌体,32℃培养过夜,第一组分别测定菌液OD600值,第二组培养液经12 000×g离心4 min,收集噬菌体裂解液,10倍比稀释后制斑,测定此时噬菌体效价。
2 结果与分析
2.1 发酵过程中不同p H对宿主菌和噬菌体的影响结果
2.1.1 发酵过程中p H对宿主菌的影响结果
在p H=5~9范围内3株乳酸菌均可正常生长,具有较强的嗜酸性。当p H=2或10时均不能生长,p H等于3时差异极显著,pH等于4时差异显著,均不能正常生长,p H等于9时差异不显著乳酸菌可以正常生长,表明对强酸、强碱较敏感。结果如图1所示。

图1 不同pH对宿主菌存活率的影响
2.1.2 发酵过程中p H对噬菌体的影响结果
在发酵介质中,不同p H对Lc、Lpen及Lbre的影响结果如图2所示。当pH=2时,Lc完全灭活,Lpen效价下降1个数量级,Lbre效价下降4个数量级;当p H=3~10时,3株噬菌体均表现出完全抗性;当pH=11~14时,3株噬菌体效价均急剧下降,最终完全灭活。
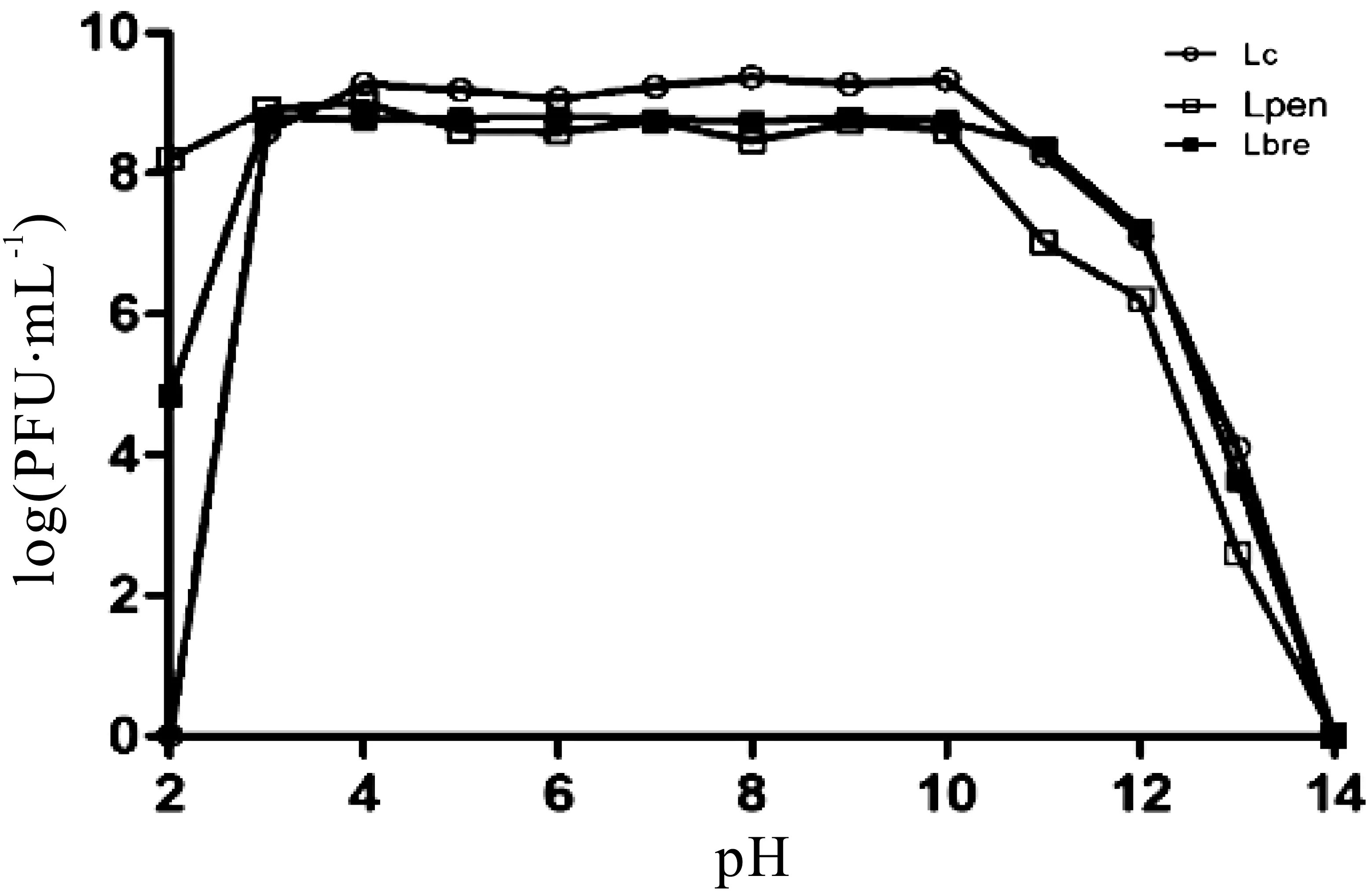
图2 不同pH对噬菌体效价的影响
2.2 发酵过程中温度对宿主菌和噬菌体的影响结果
2.2.1 发酵过程中温度对宿主菌的影响结果
3株乳酸菌L.casei ATCC 393、L.pentosus KLDS1 0413和L.brevi s ATCC 367分别于32、37、45℃和56℃进行生长曲线的测定,当温度为32℃和37℃时,3株乳酸菌均正常生长,且37℃时生长状态略好;当温度为45℃时,与37℃相比,差异极显著,3株乳酸菌均生长缓慢,生长速度L.casei ATCC 393>L.pentosus KLDS1.0413>L.brevis ATCC 367;当温度为56℃时,与37℃相比,差异极显著,3株乳酸菌均不能正常生长,结果如图3~图5所示。
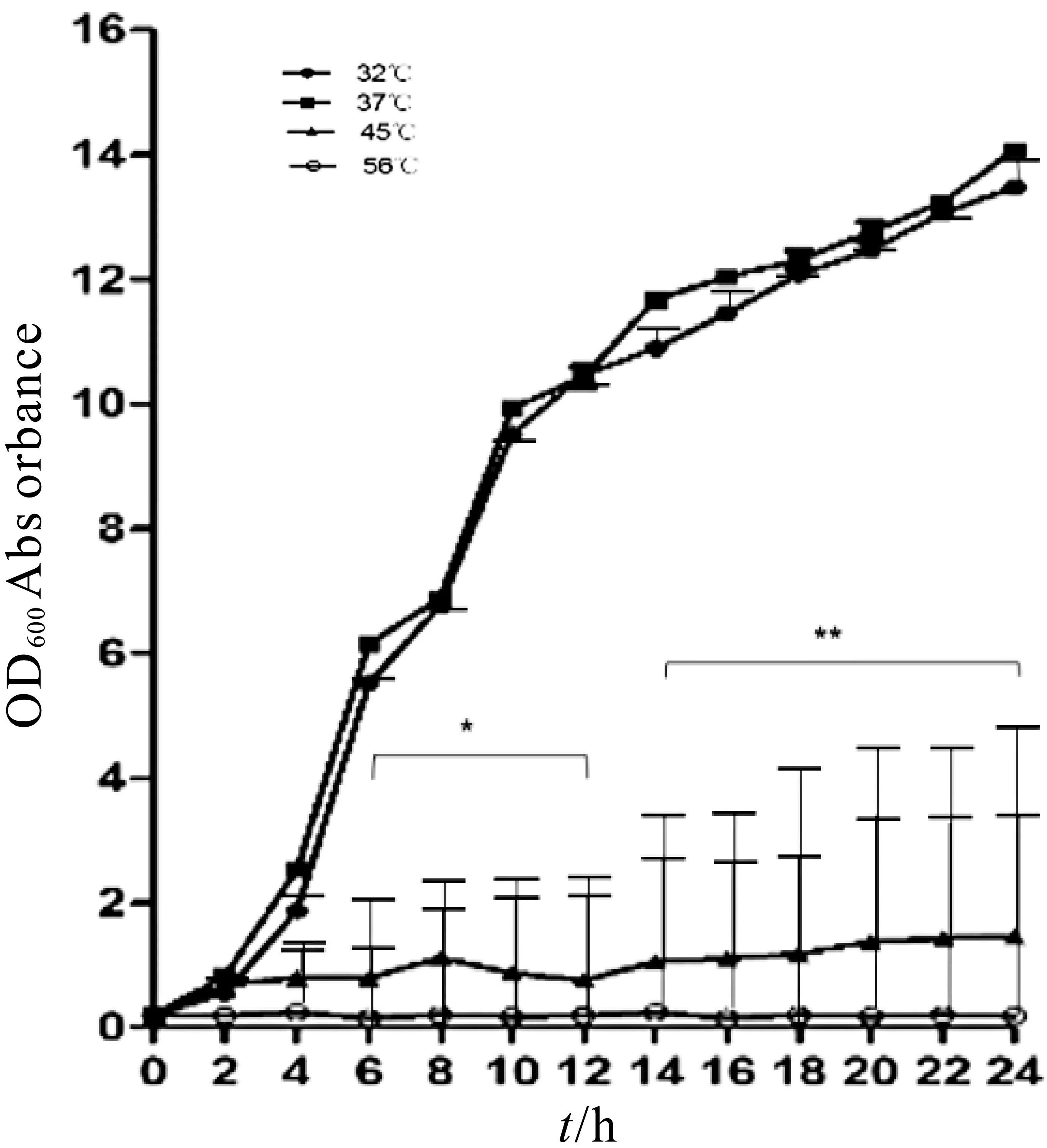
图3 温度对L.casei ATCC 393的影响

图4 温度对L.pentosus KLDS1.0413的影响

图5 温度对L.brevi s ATCC 367的影响
2.2.2 发酵过程中温度对噬菌体的影响结果
发酵介质不同温度对Lc的影响结果如图6所示,37℃时表现出完全抗性;45℃处理5 min,15%噬菌体失活,处理30 min,60%噬菌体失活,处理60 min后,70%噬菌体失活;56℃处理5 min后,可使98%Lc灭活,处理10 min后几乎全部灭活;63℃处理2 min即可完全灭活。

图6 发酵介质中温度对噬菌体Lc的影响
发酵介质不同温度对Lpen的影响结果如图7所示,当温度为32℃和37℃时,均表现出完全抗性;45℃时处理15~60 min,15%~20%噬菌体灭活;56℃作用处理45 min后,可使其全部灭活;63℃处理2 min即可完全灭活。
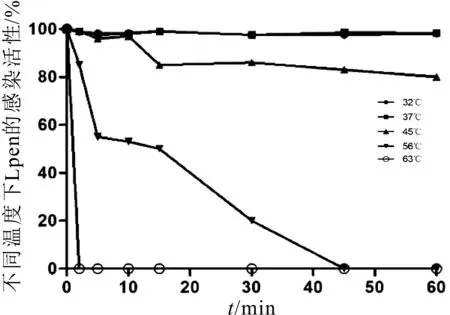
图7 发酵介质中温度对噬菌体Lpen的影响
发酵介质不同温度对Lbre的影响结果如图8所示,当温度为32℃时,表现出完全抗性;37℃处理2~10 min,可使其10%灭活;处理30~60 min,可使其30%灭活;45℃作用5~10 min,可使其30%灭活,作用60 min后,可使其70%灭活;在56℃及63℃温度下分别处理5 min和2 min即可使噬菌体全部灭活。
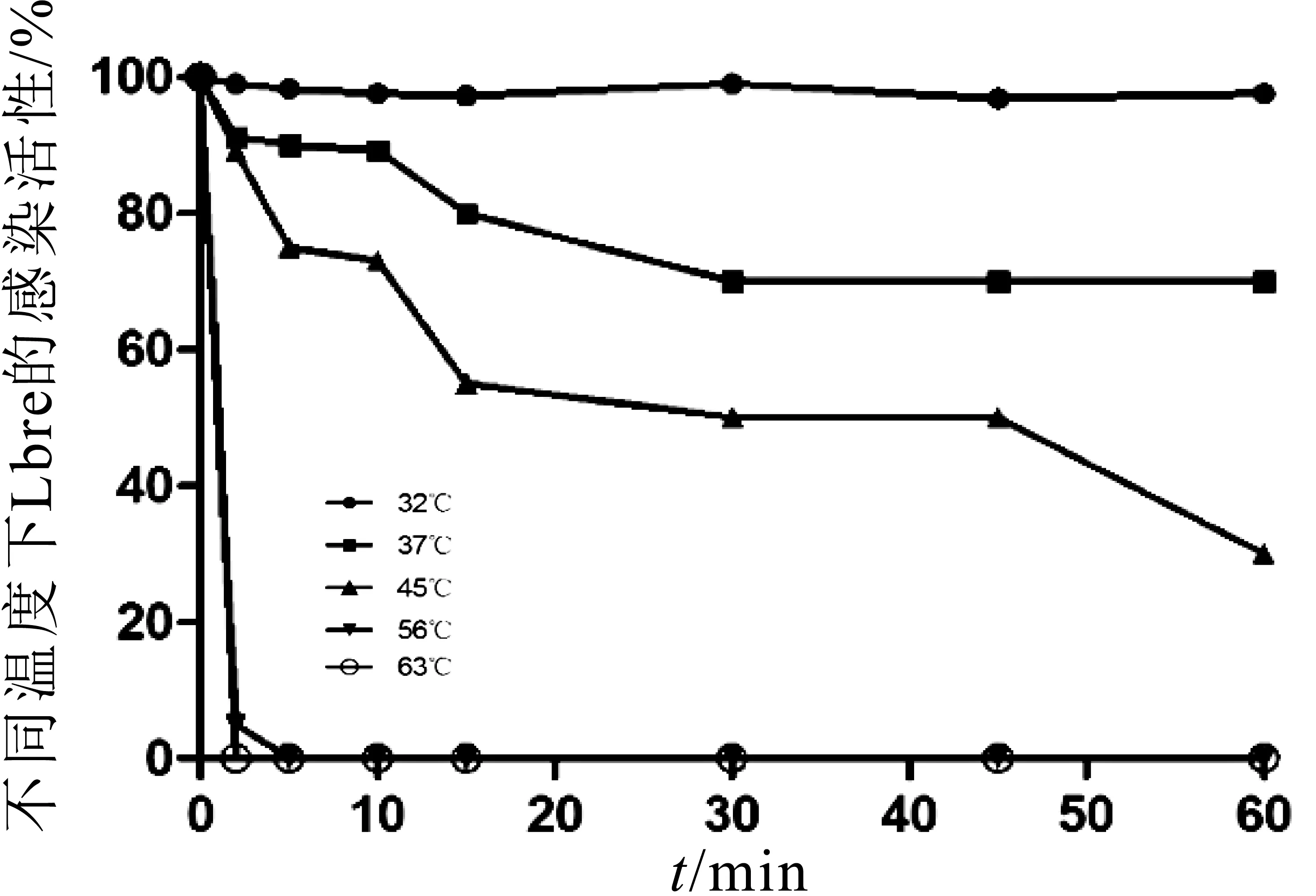
图8 发酵介质中温度对噬菌体Lbre的影响
2.3 发酵介质中二价阳离子对宿主菌和噬菌体的影响结果
2.3.1 发酵介质中二价阳离子对宿主菌的影响结果
发酵介质中添加Ca2+和Mg2+对宿主菌的影响结果如图9所示,L.casei ATCC 393、L.pentosus KLDS1.0413、L.brevis ATCC 367在MCM、MRS-Ca、MRS-Mg、MRS 4种培养基中的菌浓度均无明显变化,差异不显著,发酵液体介质中添加钙、镁离子未影响宿主菌的正常生长。
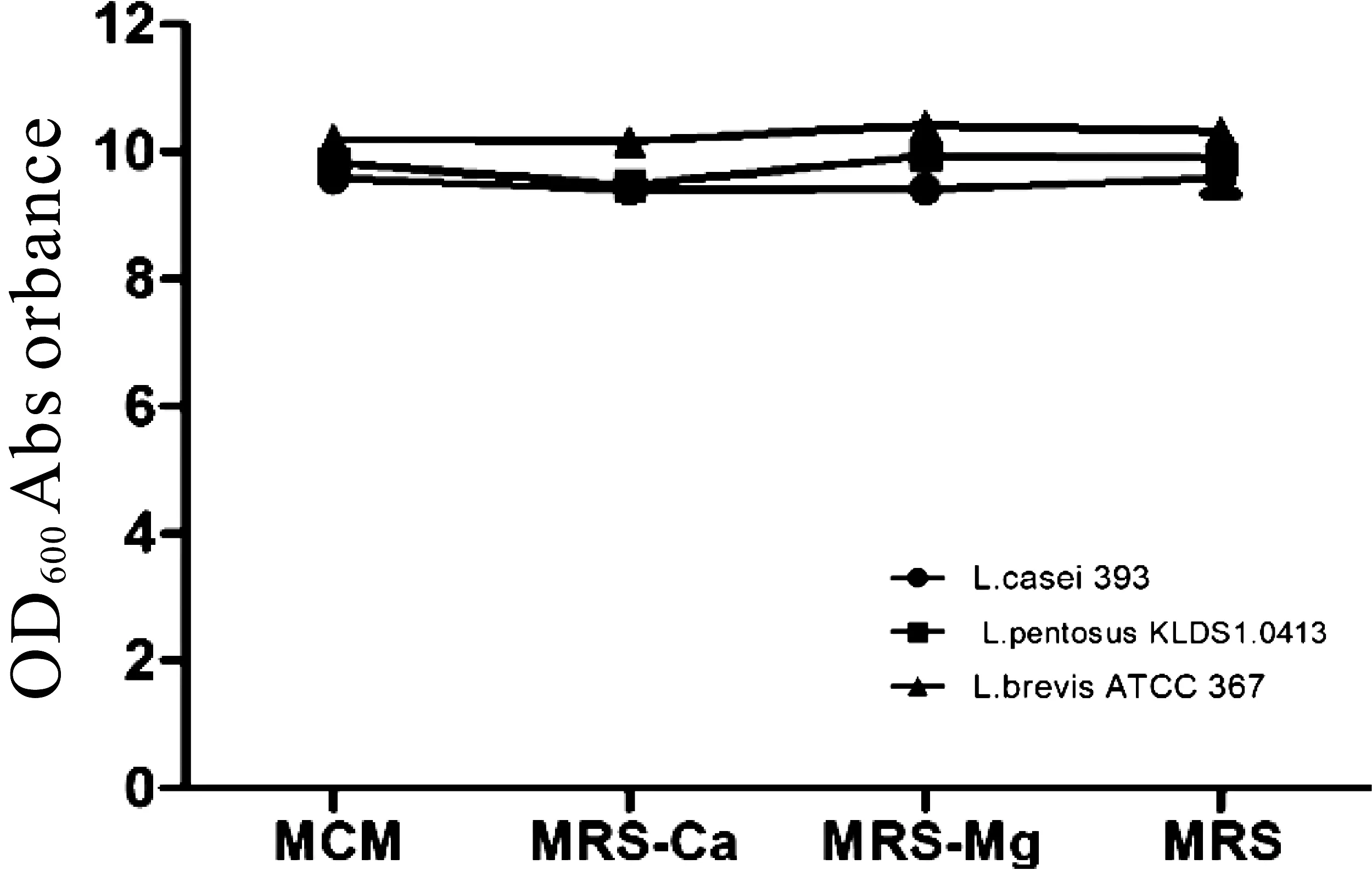
图9 钙、镁离子对宿主菌的影响
2.3.2 发酵介质中二价阳离子对噬菌体的影响结果
发酵介质中Ca2+和Mg2+对噬菌体的影响结果如图10所示,上层为未添加Ca2+和Mg2+的MRS培养基时,噬菌体效价与MCM、MRS-Ca、MRS-M g 3组相比明显降低。其中,未添加Ca2+和Mg2+使Lc效价下降2个数量级,Lpen和Lbre效价下降1个数量级;未添加Ca2+可使Lc效价下降1个数量级,Lpen效价略有降低,Lbre效价无明显影响。

图10 钙、镁离子对噬菌体的影响
2.4 发酵介质中NaCl对噬菌体和宿主菌的影响结果
2.4.1 发酵介质中NaCl对宿主菌的影响结果
在添加不同剂量NaCl的MRS培养基中,L.casei
ATCC393、L.pentosus KLDS 1.0413、L.brevis ATCC 367
均能正常生长,培养18 h后检测OD600。结果如图11所示,当NaCl浓度为2%时,L.casei ATCC 393菌液OD600最大(9.8673);当NaCl浓度为1%时,L.pentosus KLDS 1.0413菌液OD600最大(9.5430);当NaCl浓度为2%时,L.brevis ATCC 367菌液OD600最大(6.2780),与MRS组相比差异不显著,OD600数值均在乳酸杆菌正常生长的区间。
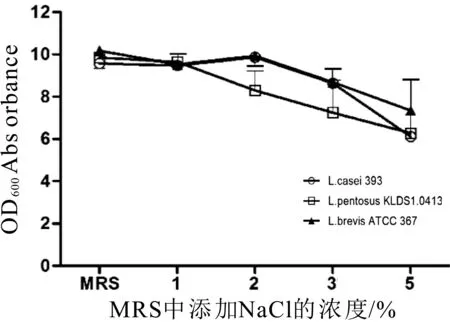
图11 不同浓度NaCl对宿主菌的影响
2.4.2 发酵介质中NaCl对噬菌体的影响结果
发酵液体介质中不同浓度NaCl对Lc、Lpen及Lbre的影响结果如图12所示。当1%NaCl作用Lc时,Lc效价未发生改变;当2%NaCl处理时,其效价下降1个数量级;当3%、5%NaCl作用Lc时,其效价下降2个数量级。当1%、2%NaCl作用Lpen时,其效价下降2~3个数量级;当3%NaCl作用Lpen时,其效价下降4个数量级,当5%NaCl处理时完全灭活。当不同浓度NaCl作用Lbre时,其耐受性较强,当NaCl浓度为3%和5%,效价下降一个数量级,但不能完全灭活。
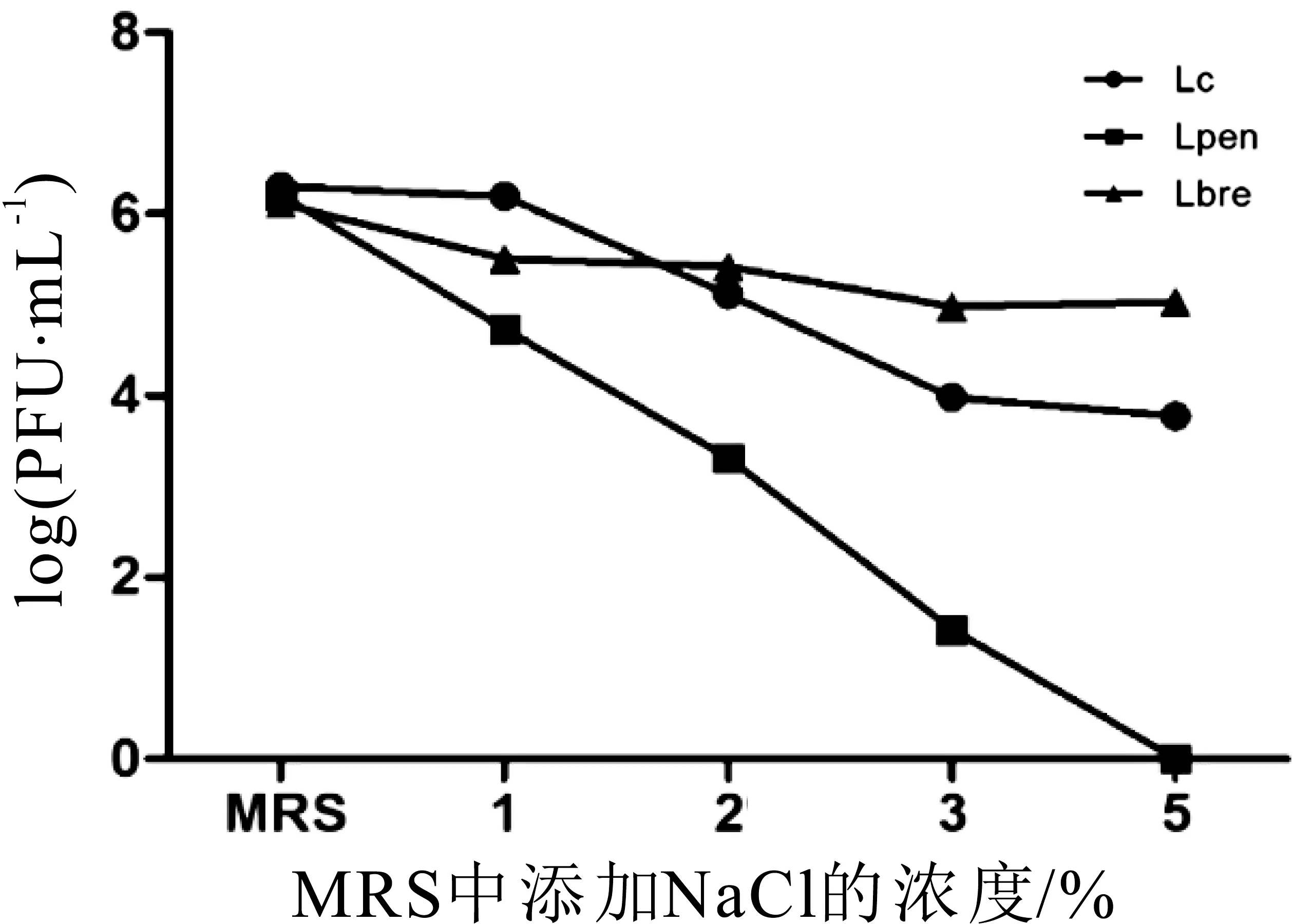
图12 不同浓度NaCl对噬菌体的影响
2.5 发酵介质中T-20对噬菌体和宿主菌的影响结果
2.5.1 发酵介质中T-20对宿主菌的影响结果
不同浓度T-20对宿主菌L.casei ATCC 393、L.pentosus KLDS 1.0413、L.brevis ATCC 367的作用结果如图13所示。随着T-20浓度的增加,3株乳酸菌的菌浓度均有降低(2%>1%>0.1%),当T-20浓度为1%、2%时,菌浓度下降较明显,但差异不显著,可以保证发酵的正常进行。
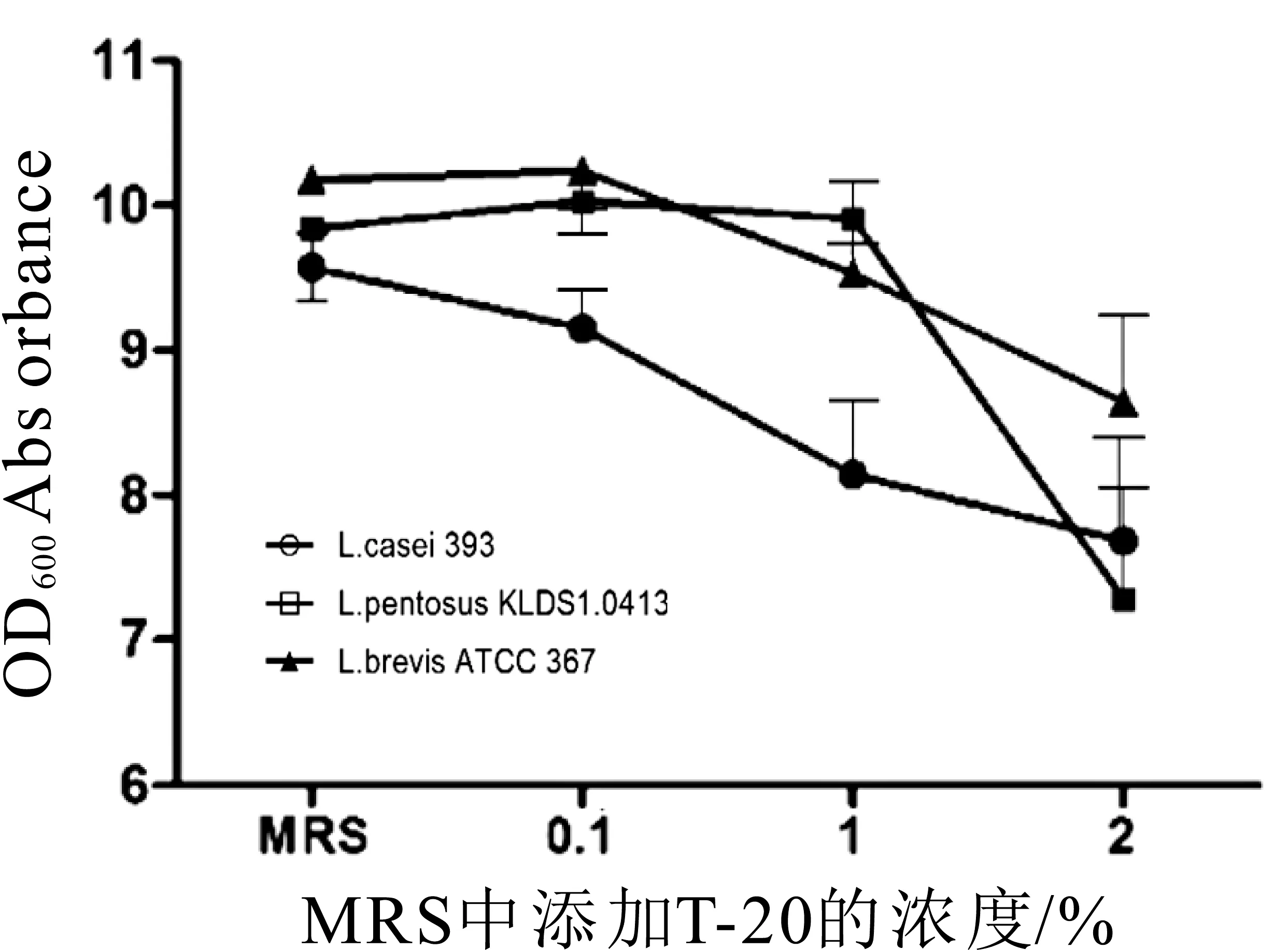
图13 T-20对宿主菌的影响
2.5.2 发酵介质中T-20对噬菌体的影响结果
不同浓度T-20对噬菌体的影响结果如图14所示。0.1%、1%T-20可使Lc效价分别下降1个和2个数量级,2%T-20使Lc效价下降3个数量级;0.1%T-20处理Lpen时,其效价下降2个数量级,1%T-20处理时Lpen效价下降5个数量级,2%处理后可完全灭活;0.1%T-20处理Lbre时效价下降4个数量级,1%、2%T-20可使噬菌体Lbre完全灭活。
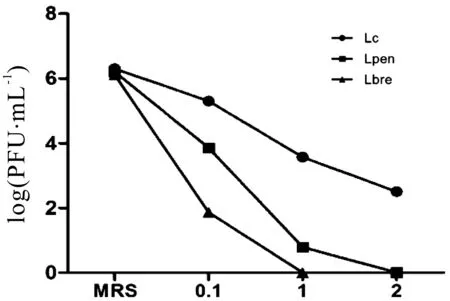
图14 不同浓度T-20对噬菌体的影响
3 讨 论
噬菌体污染的普遍存在常引起发酵失败,并导致严重的经济损失[22-23]。生产环境是影响发酵过程的重要因素之一,在发酵过程中,发酵液体的随意排放、发酵乳废弃以及洗罐后废水的不当处理等,会导致噬菌体污染发酵环境[24]。
噬菌体污染时,车间及厂区周围环境会有大量噬菌体残留,紫外消毒可有效控制发酵环境中的噬菌体污染。虽然紫外消毒覆盖面积大,操作便捷,但由于穿透力弱,对隐蔽或遮挡的发酵生产设备,例如工厂管道、发酵罐和种子罐等不能消杀彻底。发酵生产中常用的新洁尔灭(0.05%)、甲醛(0.1%)和高锰酸钾(KMnO4,0.05%)等生物灭活剂对噬菌体灭活效果非常显著[25-26]。有研究报道,对乳杆菌噬菌体MLC-A最有效的乙醇浓度为75%[13]。
在发酵过程中,介质中的相关因素对发酵过程具有重要的影响。在发酵生产中不同pH对噬菌体和宿主菌的影响不同,噬菌体对pH表现出高抗性,具有较强的耐酸、耐碱能力,但宿主菌耐酸不耐碱,并且噬菌体对p H的耐受范围与宿主菌相比较宽。因此,在发酵过程中,采用改变p H无法有效抑制噬菌体生长。另外,多数噬菌体对热处理较敏感,高温处理可破坏衣壳蛋白、裂解噬菌体。本研究将噬菌体Lc、Lpen、Lbre在56℃条件下分别作用10、45、5 min均被灭活;而宿主菌L.casei ATCC 393、L.pentosus KLDS1.0413和L.brevis ATCC 367在32℃和37℃下正常生长。综合发酵介质中温度对噬菌体与宿主菌的影响,在发酵生产中,可采取“先高后低”来设定温度,发酵初期将介质温度升至56℃及以上维持一段时间,然后通过制冷装置降至发酵菌株正常培养温度,完成发酵过程。
噬菌体在侵染宿主菌过程中,吸附阶段是首要过程。Sechaud等[27]指出Ca2+或M g2+不仅具有稳定噬菌体DNA螺旋结构、提高phage对宿主菌吸附率的作用,还可控制噬菌体DNA侵入宿主细胞。因此,二价阳离子Ca2+和Mg2+是噬菌体增殖必不可少的。本研究通过检测添加Ca2+或Mg2+对发酵过程的影响,结果表明,宿主菌在添加和未添加Ca2+或Mg2+时均能正常生长,但Lc、Lpen、Lbre在未添加Ca2+或Mg2+时,其效价下降2~4个数量级,且形成的噬菌斑较小。有研究报道,当噬菌体BYM、Ib3、YAB和MLC-A裂解菌体时,培养基中只有添加Ca2+和M g2+时,才能形成噬菌斑[28]。因此,在生产实践中,可考虑在发酵介质中添加络合物(如柠檬酸盐等)除去液体介质中多余的二价阳离子,阻断噬菌体对宿主菌的吸附,有效控制发酵过程中的噬菌体污染。
NaCl作为食品添加剂,不仅能改变发酵食品的口感质地,还可抑制噬菌体污染且不影响发酵菌株正常生长。在本研究中,当添加5%NaCl时发酵菌株浓度略有降低,但不影响发酵过程;当3%、5%NaCl处理Lc时,效价下降2个数量级,有效抑制其活性;当5%NaCl处理时,噬菌体Lpen完全灭活,Lbre无明显抑制作用。因此,在食品发酵生产中,可利用噬菌体对Na-Cl敏感性的不同,添加适宜浓度的NaCl来抑制噬菌体活性。T-20是一类非离子表面活性剂,常用于发酵过程中,减少泡沫的产生,被公认为安全、无毒、无刺激性的原料,每日允许摄入量为0~25 mg/kg(FAD/WHO,1985)。本研究选用不同浓度T-20对噬菌体及宿主菌共同作用,为了探究在合理浓度范围内添加T-20,是否可以控制发酵过程中噬菌体的污染,随着T-20浓度的增加,3株乳酸菌浓度略有降低(0.1%<1%<2%),但可以确保发酵过程顺利进行;当2%T-20作用Lc、Lpen、Lbre后,只有Lc效价下降3个数量级,而Lpen、Lbre均具有较强耐受性。因此,在生产实践中,可利用噬菌体对T-20的敏感性抑制其活性。
本研究通过分析发酵介质中不同理化因素对发酵过程的影响,为保护商业菌株、制定有效防控措施提供理论依据和实验数据,同时为深入了解探究噬菌体与宿主菌间的相互作用奠定基础。在生产实践中不同消毒剂和消毒方法各有优缺点,在发酵生产过程中,可联合应用多种方法,有效控制噬菌体污染,节约生产成本,提高生产效率。
猜你喜欢
心理学探新(2022年1期)2022-06-07 09:16:02
昆明医科大学学报(2022年2期)2022-03-29 00:51:20
中国动物传染病学报(2021年3期)2021-07-21 03:19:28
猪业科学(2018年5期)2018-07-17 05:55:18
新高考·高一数学(2016年3期)2016-05-19 09:08:30
广东海洋大学学报(2015年3期)2015-12-22 10:05:26
建筑工程技术与设计(2015年22期)2015-10-21 18:37:26
中国新通信(2015年12期)2015-05-30 02:52:19
中国当代医药(2015年8期)2015-03-01 02:01:55
赤峰学院学报·自然科学版(2015年2期)2015-02-17 08:40:44
