
中国北方野生莲居群的表型多样性分析
2021-07-07 13:45付彦荣刘凤栾李硕苏燕董丽田代科
河南农业大学学报 2021年3期
付彦荣, 刘凤栾, 李硕, 苏燕, 董丽, 田代科
(1.北京林业大学园林学院, 北京 100083; 2.中国科学院上海辰山植物科学研究中心,
莲(Nelumbonucifera)是重要的水生园艺植物,具有观赏、文化、经济和生态等多重价值[1-4]。中国的莲种质资源丰富,尚有野生莲资源存在[1,5-13]。然而,受城市化、农业生产和环境污染等人为干扰,以及洪涝等自然灾害影响,野生莲居群逐步减少甚至消失[7]。野生莲居群是莲的天然基因库,是莲种质资源开发利用的基础和新品种选育的源泉,而翔实与深入的表型多样性调查与分析是充分了解与利用它们的重要方式之一。表型多样性分析已在南天竹[14]、茶树[15]、睡莲[16-17]、兰花[18-19]、杜鹃[20-21]、牡丹[22]等观赏植物上广泛开展,在莲的栽培品种[23-27]和野生资源中均有应用。王其超等[6]对黑龙江东南部野生莲的叶径、花径、瓣径、瓣数、雄蕊数、心皮数等性状进行了观测,认为这些野生莲在形态上基本一致。薛建华[7]对黑龙江全流域的野生莲进行了调查,对花色、花瓣数、心皮数、叶脉数和种子重量等性状进行了观测,发现居群间没有明显形态差异。黑龙江流域野生莲的心皮数高于湖北省洪湖野生莲,但莲种子形态未见明显差异[8]。黑龙江月牙湖、河北白洋淀、山东微山湖、湖北洞庭湖和江西鄱阳湖5个野生莲种群的莲子形态差异较大[9]。詹清华等[9]比较了内蒙古敖汉旗、辽宁普兰店、河北白洋淀、山东微山湖、湖北东湖、湖北洪湖、湖南株洲、广东三水8个野生种群引种栽培后的形态性状,认为野生莲从北向南散布过程中,为了适应光照时长变短的环境改变,叶片有增大的趋势。据此,中国北方地区尤其是黑龙江省的重要野生莲居群已有较为丰富的表型信息。本研究对更多居群(17个中国北方代表性野生莲居群)和更全面的性状(25个表型性状)进行调查和采集,通过表型多样性和聚类分析,揭示其表型变异程度,以及表型变异与居群地理分布的关系等,为野生莲居群资源保护和育种利用提供参考。
1 材料与方法
1.1 样本选取
选取中国北方黑龙江、吉林、辽宁、内蒙古、河北和山东6省17个野生莲居群为研究对象(表1),GPS 测定各居群的经度和纬度。于2016年7—8月间,从居群不同部位选取叶和花进行形态学观测。每个居群内叶片和花朵的样本数分别不少于15个和5个。鉴于莲较强的无性繁殖能力,地下茎延伸可达7~12 m[1],叶片和花朵样本距离均大于15 m。

表1 17个野生莲居群的基本信息Table 1 Basic information of 17 wild Asian lotus populations
1.2 表型性状的选取和观测
参考孔德政等的方法[22],《莲种质资源描述规范和数据标准》[28]和《植物新品种特异性、一致性和稳定性测试指南 莲属》(NY/T2756—2015)[29],选取反映野生莲形态特征且适合野外观测的25个地上部表型性状进行观测和记录。其中,每个样本的数量性状(1~15)均测量3次,取平均值。假质量性状(16~25)参考《植物新品种特异性、一致性和稳定性测试指南 莲属》(NY/T2756—2015)进行特征描述和数值化赋值(表2,表5)。

表2 表型性状观测方法和赋值Table 2 Observation methods and values assignment phenotypictraits
高度性状选取最高的立叶或花观测,长度、宽度、直径、叶姿、叶色、叶面质地、花态、花瓣形、花瓣色、瓣脉特征等性状均选取最大的立叶和花(盛开状态)观测,花瓣性状选取最大的花瓣(从外向内第11~13片花瓣)观测,花蕾形、花蕾色、花瓣数、雄蕊数、心皮数、花托形均选取接近开放(所有花被片均未脱落)的花观测。
1.3 数据处理和统计
运用 Microsoft Excel 2016 和SPSS 19.0进行数据统计分析
变异系数CV=(均值/标准差)×100%;
性状多样性采用Shannon多样性信息指数(Shannon-Weaver diversity indxe,I)
I=-∑PilnPi
式中:Pi为第i种变异类型出现频率;
居群表型聚类分析采用组间平均连接法,种质间距离为欧氏距离[14-16]。
2 结果与分析
2.1 北方野生莲居群数量性状多样性和变异特征
17个野生莲居群数量性状间差异明显(表3)。15 个数量性状的方差(F)值为 2.804~57.706,由大到小依次为花径>立叶高>花柄高>心皮数>立叶长>立叶宽>叶柄径>雄蕊数>叶脉数>花柄径>花瓣长>花柄孔数>花瓣宽>花瓣数>叶柄孔数。
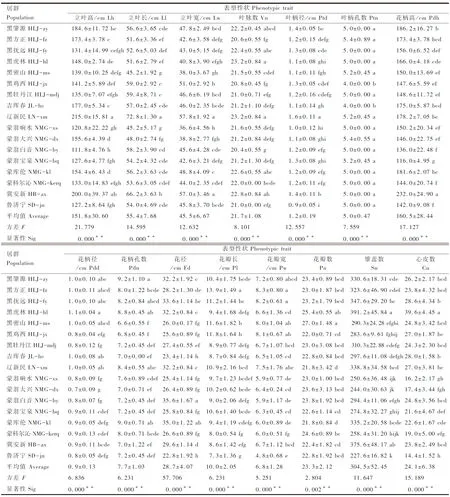
表3 北方野生莲居群15个数量性状的方差分析Table 3 Variance analysis based on 15 quantitative traits of wild Asian lotus populations from northern China
15个数量性状在居群间的平均变异系数为4.97%~26.51%(表4)。叶脉数、花瓣数和叶柄孔数的平均变异系数均小于10%,分别为4.97%、9.08%和9.55%。立叶长、立叶宽、叶柄径等9个性状的平均变异系数为10%~19%。立叶高、花瓣长和心皮数的平均变异系数均大于 20%,分别达20.16%、20.55%和26.51%。
单个居群内的所有数量性状的平均变异系数为5.09%~10.33%(表4)。

表4 北方野生莲居群15个数量性状的变异分析Table 4 Variation analysis for 15 quantitative traits of wild Asian lotus populations from northern China
2.2 北方野生莲居群假质量性状的变异特征
17个野生莲居群假质量性状的多样性较低,居群间表现不一。10个假质量性状的Shannon-Weaver 多样性指数I值为0.00~0.81(表5)。花蕾色和瓣脉特征两个性状的I值均为0,分别只有一种性状表现。花蕾色全部为粉紫色,瓣脉特征全部为明显,且所有居群的表现均相同。叶姿、叶色、叶面质地、花态、花瓣色和花托形6个性状的I值为0.36~0.68,均有两种性状表现且在居群间的分布频率不同(表5)。多数居群的叶姿为凹形(70.59%),少数为平展(29.41%);多数居群的叶色为绿色(58.82%),少数为深绿(41.18%);多数居群的叶面质地为粗糙(68.00%),少数为光滑(32%)。多数居群的花态为碗状(70.59%),少数为杯状(29.41%);多数居群的花瓣色为粉色(88.24%),少数为粉紫色(11.76%)。多数居群的花托形为倒圆锥形(82.35%),少数为喇叭形(17.65%)(表5)。花蕾形和花瓣形两个性状的I值最大,均为0.81,有3种表现。多数居群的花蕾形为卵形(64.71%),其次为窄卵形(29.41%)和圆锥形(5.88%);多数居群的花瓣形为卵圆形(70.59%),其次为阔卵圆形(17.65%)和窄卵形(11.76%)(表5)。
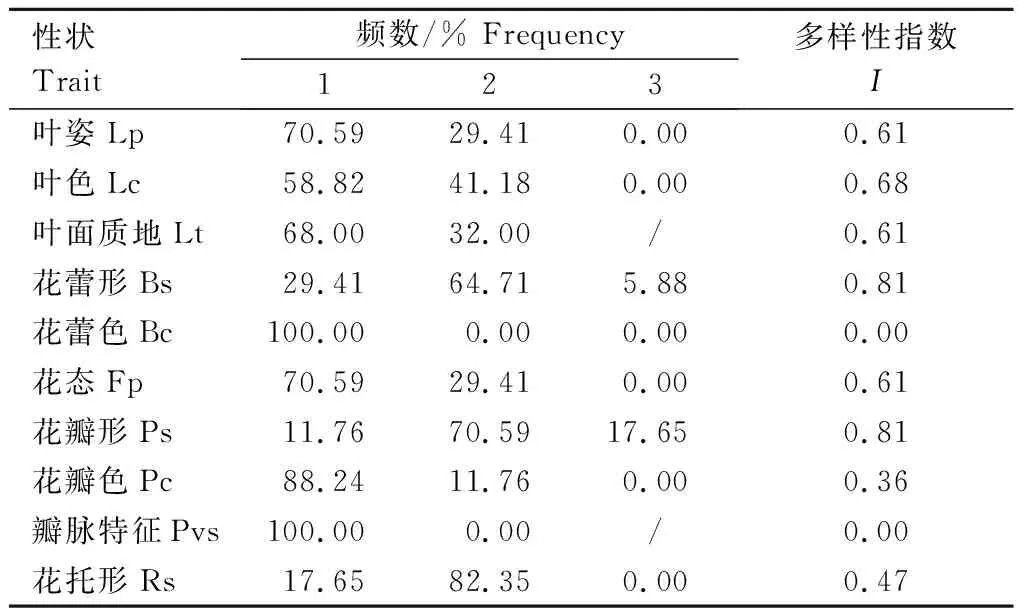
表5 北方野生莲居群假质量性状的分布频率及多样性Table 5 Frequency distribution and diversity of pseudo-qualitative traits of wild Asian lotus populations from northern China
2.3 北方野生莲居群聚类分析
17个野生莲居群的欧氏距离为1~25。基于欧氏距离的聚类结果(图1)显示,在欧氏距离约25 处,17 个居群大致分为3组,A组包含黑肇源、蒙库伦、黑抚远、黑方正、蒙科尔沁、蒙翁响水、蒙翁宝泉、蒙翁大兴、蒙翁白音、黑牡丹江、吉珲春、黑鸡西、黑密山、鲁济宁14个居群,B组包含辽新民和冀安新2个居群,C组仅包含黑虎林居群。A组中,黑密山和鲁济宁两个居群各为一支,其他的12个居群在欧氏距离约8处,又分为3个亚组,A1亚组包含黑肇源、蒙库伦、黑抚远、黑方正4个居群,A2亚组包含蒙科尔沁、蒙翁响水、蒙翁宝泉、蒙翁大兴、蒙翁白音5个居群。A3亚组包含黑牡丹江、吉珲春和黑鸡西3个居群。

图1 基于欧氏距离的北方野生莲居群表型性状聚类分析
17个野生莲居群的聚类分组结果与居群的地理分布呈现一定相关性。A组中,A1、A3亚组多数为黑龙江和吉林的居群,A2亚组全部为内蒙古的居群。冀安新居群归为B组,与A组居群呈现明显分化。鲁济宁虽归为A组,但单列为一支,表明与同组居群的相似度较低。辽新民和黑虎林两个居群情况例外,虽然与A组居群地理上相对接近,但未聚为一组。
3 讨论与结论
植物资源的多样性研究不仅有利于种质资源的科学评价和保存,更有助于挖掘有益种质资源,实现资源创新和科学利用。植物形态变异在某种程度上能够反映遗传变异的大小。通过形态学或表型性状来检测植物遗传变异和分析遗传多样性,具有直观且简便的优点[30-31]。
变异系数(CV)在某种程度上反映出群体数量性状的差异性和多样性,变异系数大则分化程度高、多样性显著,相反则群体相对稳定、变异小、一致性较高。本研究表明,中国北方野生莲居群数量性状的分化程度较低。叶脉数、花瓣数、叶柄孔数的平均变异系数均小于10%,表明这些性状分化程度较低,在进化过程中比较稳定。立叶高、花瓣长和心皮数的平均变异系数均大于 20%,表明这些性状分化程度较高,变异较为明显。
多样性指数反映了假质量性状的多样性水平。本研究表明,中国北方野生莲居群的假质量性状多样性水平总体较低。其中,花蕾色和瓣脉特征2个性状的多样性指数均为0,多样性极低。花蕾形和花瓣形多样性指数值达0.81,多样性较高。
相比睡莲属[17]、兰属[18]、带叶兜兰[19]、杜鹃花属[20]、大白花杜鹃[21]等其他物种而言,北方野生莲居群的表型多样性水平总体偏低,且明显低于莲品种的多样性[26-27],可能与野生种质特性有关,在其繁衍过程中较少受到外来基因的影响,导致性状分化较小。立叶高、花瓣长、心皮数、花蕾形、花瓣形等性状在野生莲居群间的变异程度较大,且与观赏性有较强的关联性,应在野生莲引种和育种利用时重点关注。
基于欧氏距离,17个北方野生莲居群被分为3组。聚类结果与地理距离呈现出一定的相关性。地理分布接近的居群在聚类图上也较接近,彼此分化较小,该结果与董振发等[5]、王其超等[6]、薛建华[7]的结论基本一致。黑龙江、吉林,甚至内蒙古的野生莲资源在遗传上可能无明显分化。冀安新和鲁济宁居群与东北地区居群遗传上存在分化,可能与远距离的地理隔离有关。黑虎林居群与黑龙江省其他居群发生分离,可能与其位于黑龙江所有居群最东端且相对孤立的地理区位特点有关。王其超等[6]的调研也发现,黑虎林居群的叶径略大于黑龙江东南部其他居群,瓣数和心皮数等也略多。冀安新与辽新民聚为一组,聚类结果与他们的地理分布不一致,可能因为这些居群材料间存在遗传混杂所致,也可能其中某个居群是很久以前人工引种后,又在当地长期自然繁衍而成。
表型性状是基因和外部环境综合作用的结果,特别是数量性状,容易受到环境差异的影响,导致分析结果发生偏差。本研究基于表型性状获得了对北方野生莲居群遗传多样性的初步认识,为野生莲资源的保护和育种应用提供了参考。以形态学分析为基础,辅助分子标记和基因测序等方法,对野生莲资源的遗传多样性开展深入研究,有助于获得更全面的信息。
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
中国现代医生(2022年21期)2022-08-22
农村科学实验(2022年2期)2022-03-12
支部建设(2020年15期)2020-07-08
三农资讯半月报(2020年2期)2020-03-09
作文与考试·小学低年级版(2017年13期)2017-07-06
第二课堂(课外活动版)(2015年5期)2015-10-21
作文与考试·小学低年级版(2015年8期)2015-09-25
百科知识(2015年18期)2015-09-10
小星星·阅读100分(高年级)(2015年4期)2015-05-26
