
多主体创新产出演化分析
——生态关系和结构洞交互影响下
2021-07-05 11:11:38陈秋琴盛永祥施琴芬
科技管理研究 2021年10期
陈秋琴,吴 洁,盛永祥,刘 鹏,施琴芬
(1.江苏科技大学经济管理学院,江苏镇江 212003;2.苏州科技大学商学院,江苏苏州 215009)
1 研究背景
科学技术的飞速发展使得单个企业难以独揽全部前沿技术,技术创新的复杂性和不确定性促使企业寻求拥有互补性技术资源的优质伙伴进行合作创新,从而形成了多个企业赖以生存的创新生态系统。在创新生态系统中,由于各主体的资源和竞争优势不尽相同,会使主体占据不同的网络位置,同时也使得各主体之间呈现出不同的生态关系,主体的网络位置特性与主体间的不同生态关系动态交互地影响着各主体的创新活力和创新产出。因此,探究创新生态系统中创新主体间的生态关系及其所占据的不同网络位置,对促进创新生态系统的良性发展具有重要意义。
众多学者从不同角度分别针对创新生态系统主体间生态关系和主体所处的网络位置展开了深入研究。一类典型的研究是从创新生态系统中各创新主体间共生演化模式、竞合关系进行分析和阐述,如欧忠辉等[1]通过构建创新生态系统共生演化的动力学模型,探讨核心企业共生单元与配套组织共生单元在不同共生环境下的演化路径;叶斌等[2]建立区域创新网络共生演化模型,通过仿真模拟两类创新主体间相互作用下的共生演化态势;赵坤等[3]以核心企业和卫星企业为对象,研究众创式创新网络中两类主体间的共生关系;Mantovani 等[4]从生态系统视角出发,探究了核心企业与上下游企业之间的技术依赖结构对核心企业的创新成果和竞争优势的影响;吴洁等[5]通过构建考虑中介机构参与下的知识转移博弈模型,探讨联盟企业与中介机构等多方主体创新能力的影响因素和策略选择;陈璐等[6]构建多主体行为状态演化方程,探讨了多主体间不同竞合关系对专利创新生态系统的影响。此外,也有学者从网络关系嵌入视角出发对创新主体的网络位置与创新产出之间的联系进行了研究,如李健等[7]通过考察组织间合作研发网络,发现结构洞对企业探索式创新绩效具有积极作用;Akbar 等[8]从企业的网络结构层面出发,探讨企业之间的网络联系和结构对企业创新绩效的影响;Liao 等[9]通过考察在合作网络中占据多个结构洞的企业,发现企业技术知识多样性和知识处理能力决定了企业外部网络位置对技术创新的影响;赵炎等[10]选取我国通信设备行业的联盟企业研究企业间联盟网络特征对创新绩效的影响,发现结构洞对企业创新的影响受到企业内部自有资源的正向调节。
综上可知,已有研究大多以2 个或3 个创新主体为研究对象,分别从主体间的生态关系演化规律或合作网络中主体所占据的网络位置,对创新能力、创新方式选择及创新产出的影响展开了探讨,取得了颇为丰富的研究成果,然而多个创新主体之间存在结构洞时,已有研究较少考虑创新生态系统中创新主体间的生态关系与其所处网络不同位置特性对各主体创新产出的交互作用,也鲜有给出间接联系的主体间生态关系及其网络位置两类因素交互影响下各主体的创新产出演化规律。
针对已有研究的不足,本研究建立了多主体创新产出模型,通过仿真分析与实际案例相结合,阐明创新主体在网络位置特性与不同生态关系交互影响下创新产出演化的内在机理。本研究可能具有的创新之处在于:将生态学理论与结构洞理论相结合,厘清了创新主体之间的复杂生态关系与位置特性的内在逻辑关系,将生态关系进一步细分为直接生态关系与间接生态关系,综合考虑结构洞存在时不同位置主体之间不同生态关系类型与不同关系强度的交互作用对各主体创新产出的差异化影响,以期为创新主体实现共生共赢的生态创新提供理论指导。
2 构建多主体创新产出模型
2.1 创新生态系统中主体间关系
结构洞理论最早由Ronald[11]提出,从关系缺失的角度来看,如果网络中某个行动者所连接的另外两个行动者之间不存在直接联系时,那么该行动者所占据的网络位置就是结构洞。创新生态系统中创新主体之间的结构洞如图1 所示,连接两个无直接联系的创新主体x1和创新主体x3的创新主体x2就处在结构洞位置,x2(占据结构洞位置)称为中间主体,x1和x3(处在结构洞边缘)称为边缘主体。创新主体间的技术资源异质性及其相互依赖性导致的优势互补,决定了创新主体间的复杂生态关系。已有大量研究表明,存在直接联系的主体之间会呈现出多样化的生态关系,其中就包含竞争共存(-,-)、寄生共生(+,-)/(-,+)、互惠共生(+,+)等3 种不同的关系类型,而x2为原本无直接联系的x1与x3建立起了传递信息与资源交互的渠道。
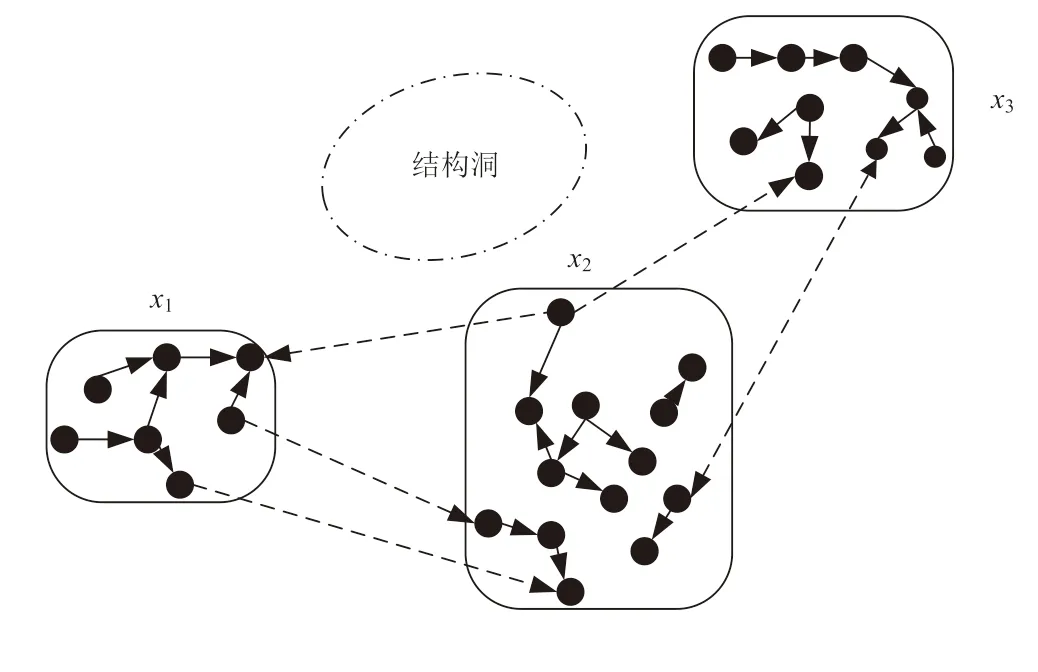
图1 创新生态系统中创新主体间结构洞示意
创新生态系统中的主体通过开放式创新获取外部资源,实现创新产出的价值增值。结构洞的存在为创新主体搜寻更多创新信息与资源提供了捷径,特别是结构洞形成的边缘主体的间接联系可以为创新主体带来异质性外部信息[12],进而影响创新主体的创新活动与创新产出,这也正是处在结构洞边缘的创新主体之间呈现复杂生态关系的主要原因。因此,边缘主体之间的生态关系类型也主要归类为竞争共存、寄生共生、互惠共生3 种类型。
此外,由于各主体的资源和竞争优势不尽相同,各主体间的生态关系必然存在关系强度的差异,关系强度的变化会带来主体间信息和资源传递的差异性,进而影响创新主体的创新产出[13]。因此,在创新生态系统中,中间主体与边缘主体之间、边缘主体之间均呈现出复杂且多样化的生态关系类型,两类主体的生存和发展受各主体间生态关系和位置特性的交互作用而呈现出不同的演化态势,进而影响创新生态系统的发展。
2.2 提出假设
基于上述分析,提出以下假设:
假设1:资源制约。各创新主体在创新过程中所能投入的发展资源有限,两类主体凭借各自的竞争优势争夺资源和市场,在有限资源和有限市场容量的制约下不会呈现无限增长趋势,而是在经过一段时间的激烈竞争后,最终达到稳定状态。
假设2:生态关系。创新生态系统中,创新主体间的技术资源异质性及其相互依赖性导致的优势互补决定了主体间的复杂生态关系,且创新主体之间的联系既包含直接联系也包含间接联系,因而各主体之间的生态关系会表现出竞争共生、寄生共生、互惠共生3 种不同的关系类型。
假设3:位置特性。创新生态系统中存在结构洞时,拥有关键性资源和核心技术的创新主体较容易占据结构洞位置,此时由中间主体连接起来的两个创新主体就处在结构洞边缘位置,位于结构洞边缘位置的主体就拥有了向对方传递间接影响的途径。
假设4:关系强度。关系强度代表着技术创新网络中主体间关系的强弱程度,关系强度的不同将带来主体间信息和资源传递的差异,中间主体与边缘主体之间的直接关系强度和边缘主体之间的间接关系强度的差异,会导致各创新主体的创新产出呈现出不同的演化态势。
2.3 模型构建
鉴于Lotka-Volterra 模型对主体间交互现象的良好描述,逐渐被用于研究多主体之间的关系演化[14],因此本研究在Lotka-Volterra 模型的基础上引入间接作用系数βij以探究结构洞存在时对各创新主体创新产出的影响。根据上述4 个假设可得到创新生态系统的多主体创新产出模型如下:
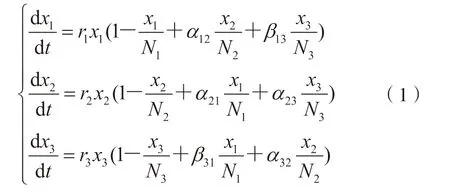
以两类创新主体为例,说明竞合系数的生物学含义:
(1)当α12<0、α21<0 时,表明x1与x2为竞争共存关系,即创新生态系统中两个创新主体间的竞争强度大于合作强度,主要表现为竞争关系,相互抑制彼此的发展。
(2)当α12>0、α21<0 或α12<0、α21>0 时,表明x1与x2为寄生共生关系,即创新生态系统中技术资源薄弱的主体依赖资源雄厚的主体进行发展,寄生者通过宿主的资源供给不断扩展自身的发展空间,宿主则可能因寄生者的消耗而不利于自身的发展。
(3)当α12>0、α21>0 时,表明x1与x2为互惠共生关系,即创新生态系统中两个创新主体密切合作,互利共赢。
3 仿真分析
为探究不同生态关系和结构洞交互作用下各创新主体的创新产出演化规律,本研究采用MATLAB2016a 仿真软件对两类(3 个)创新主体的创新产出演化态势进行模拟,设定各主体的增长率r均为0.1,最大环境容纳量N为500;竞合系数α12=0.000 1,α21=0.000 2,α23=0.000 2,α32=0.000 1;预设竞合系数的符号全为“+”,在分类讨论时根据不同的生态关系改变相应竞合系数的正负号;3 个主体创新产出的专利数量初始值分别为n1=2 件、n2=10件、n3=5 件;仿真运行周期t=200。
(1)边缘主体之间竞争共存且均与中间主体竞争共存。图2 是创新生态系统中其他参数不变,间接作用系数变化时各创新主体创新产出的演化态势。从图2 可知,两类主体同处在竞争共存状态时,中间主体x2均呈现倒“U”型增长趋势,边缘主体x3通过x2传递单向性的强间接影响会抑制对方发展,而边缘主体x1与x3之间传递双向性的强间接影响会加剧双方的竞争激烈程度。这是因为两类主体间相互竞争,各主体的创新产出势必会受到抑制,而结构洞的存在使得中间主体不仅可接收来自边缘主体的信息,而且为这一边缘主体提供了传递信息给另一类边缘主体的机会。两类主体间竞争共存的生态关系使得经由结构洞接收的信息反而具有更强的不确定性和风险,导致创新主体不敢轻易利用,而是继续在原有基础上进行激烈竞争,此时结构洞的存在会加剧创新主体间的激烈竞争程度,导致创新生态系统的运转效率低下,总体创新产出不高。
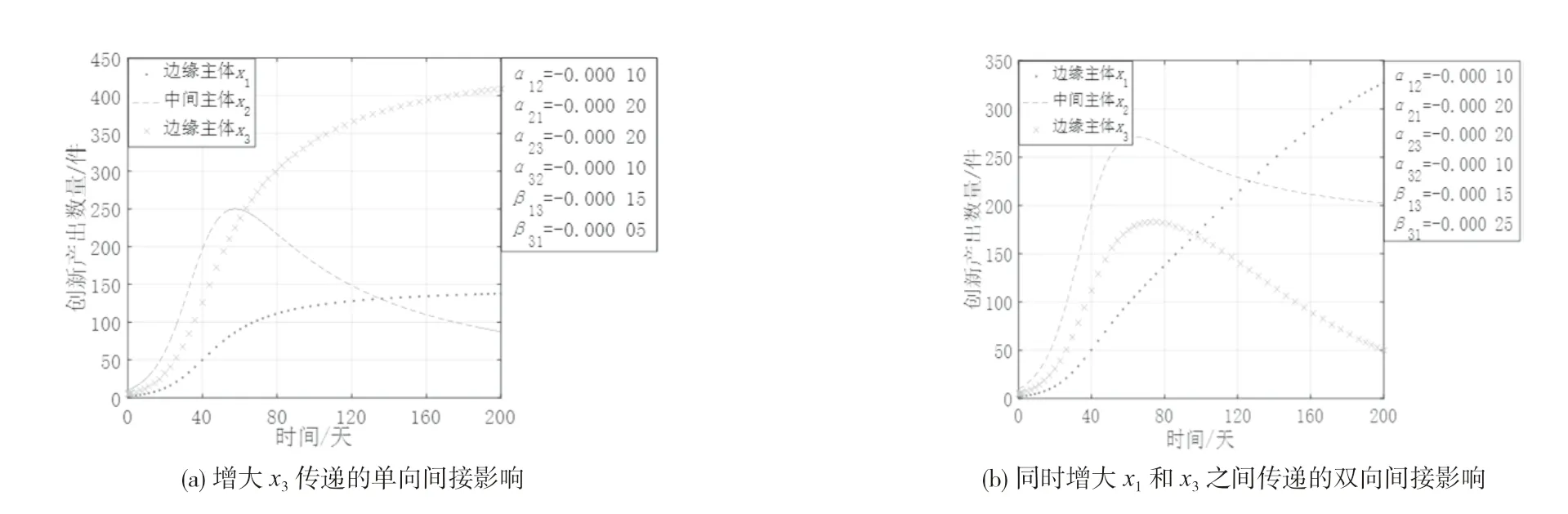
图2 创新生态系统中中间主体与边缘主体均竞争共存的演化态势
(2)边缘主体之间寄生共生且均与中间主体竞争共存。图3 是创新生态系统中其他参数不变,间接作用系数变化时各创新主体创新产出的演化态势。从图3 可知,宿主x3通过中间主体x2传递单向性的间接影响明显促进其寄生者x1发展,同时抑制x2的创新产出增长;x1通过x2传递单向性的强间接影响会抑制x3发展,同时抑制x2的创新产出增长;x1与x3通过x2传递双向性的间接影响会使x3和x2分别都呈现倒“U”型增长趋势。造成这种结果的原因是边缘主体间寄生共生,作为寄生者的边缘主体会不断消耗其宿主的创新活力,结构洞的存在使得宿主的变化革新更容易被作为寄生者的边缘创新主体所利用以促进其自身的发展,而边缘主体与中间主体间的竞争关系又使得中间主体举步维艰,此时创新生态系统中只有作为寄生者的边缘主体获得了较高增长率,系统的技术创新风险上升。
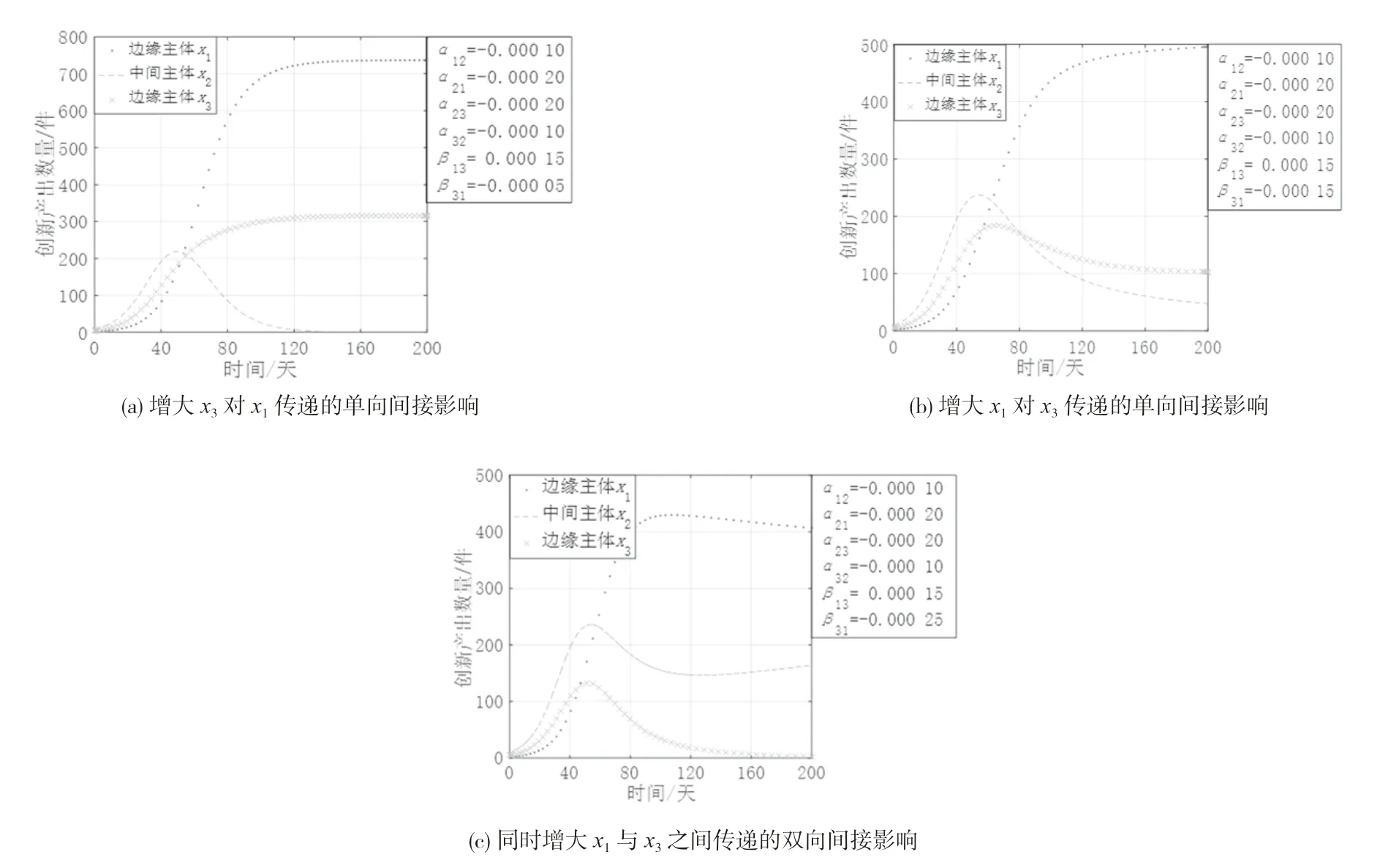
图3 创新生态系统边缘主体之间寄生共生的演化态势
(3)边缘主体之间互惠共生且均与中间主体竞争共存。图4 是创新生态系统中其他参数不变,间接作用系数变化时各创新主体创新产出的演化态势。从图4 可知,边缘主体x1与x3通过中间主体x2传递单向性或双向性的强间接影响都会促进x1和x3发展,同时削弱x2的创新产出,x2呈现倒“U”型增长趋势甚至被淘汰出局。这是因为边缘主体间互惠共生,结构洞的存在更容易激发它们之间的创新活力,边缘主体通过中间主体传递的间接影响于对方而言是良性的,加以充分利用可获得较高增长,但单向性的间接影响相比双向性的间接影响所带来的促进作用略低;随着边缘主体的发展壮大,与边缘主体保持竞争关系的中间主体的生存空间不断缩减,其竞争优势不断被削弱,生存发展受到两方的强烈抑制,因此更容易被淘汰出局。
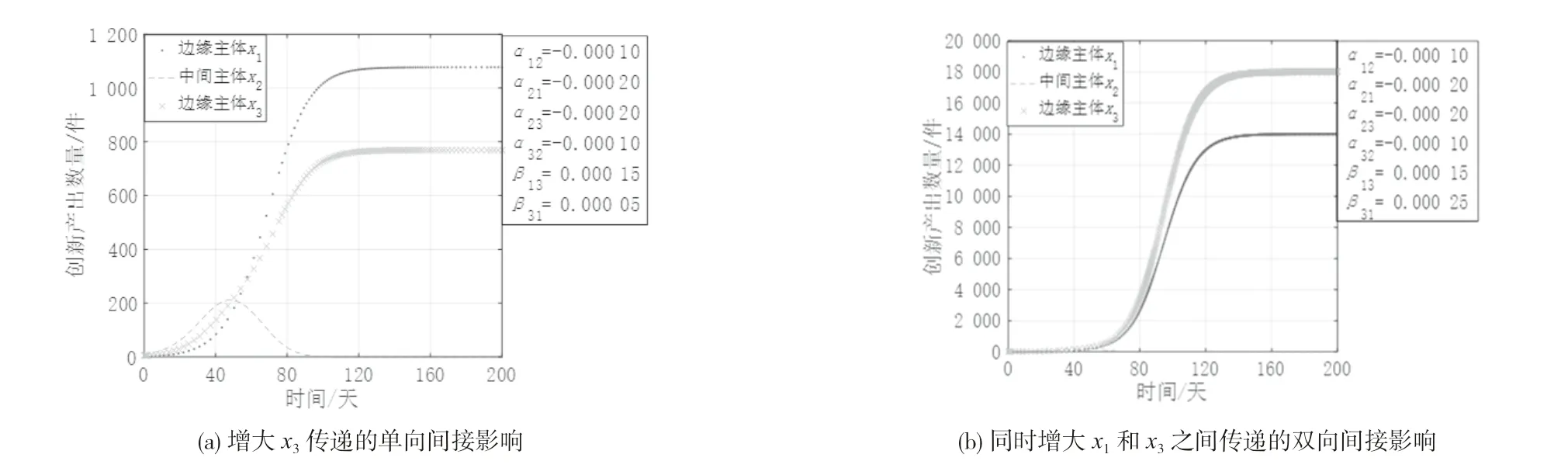
图4 创新生态系统边缘主体之间互惠共生的演化态势
(4)边缘主体间寄生共生时与中间主体竞合强度变化。中间主体与边缘主体之间存在的直接联系使得它们之间呈现出复杂多样的直接生态关系,限于篇幅,选择边缘主体间寄生共生且均与中间主体竞争共存这一典型情况进行分析,以探究中间主体与边缘主体间的直接生态关系对各创新主体的影响。图5 是边缘主体之间寄生共生时中间主体与边缘主体的竞合强度变化下各创新主体的创新产出演化态势。由图5 可知各主体的演化态势存在较大差异:
1)如图5(a)所示,边缘主体x1和x3间寄生共生,寄生者x1对中间主体x2的竞争强度减弱时,x2呈现持续增长的趋势,x1和x3均呈现倒“U”型增长趋势。这是由于作为寄生者的边缘主体对中间主体的抑制作用减小时,中间主体获得增长空间并发挥其占据结构洞的资源与信息优势,从而提升自身的创新产出,同时边缘主体的生存空间也逐渐减小,因而呈现倒“U”型增长趋势。
2)如图5(b)所示,边缘主体x1和x3间寄生共生,中间主体x2对寄生者x1的竞合强度增强时,x2与作为宿主的x3均呈现倒“U”型增长趋势,x2甚至出现被淘汰出局的现象,而x1 却呈现持续增长的趋势。这是因为中间主体对寄生者的抑制作用增大时,中间主体可在短时间内获得较快的增长速度,寄生者因受到更强的抑制,其增长速度逐渐减缓,宿主的被动消耗降低可促进宿主的发展,同时也挤占了中间主体的生存空间,中间主体的创新产出逐步下降,此时寄生者开始逐步恢复并从宿主处得到发展所需的资源,其创新产出稳步增长。
3)如图5(c)(d)所示,边缘主体x1和x3间寄生共生,作为宿主的x3与中间主体x2的竞合强度同时增大或减小时,x3与x2均出现倒“U”型增长趋势,相比之下,两者在竞合强度同时增大时到达拐点的速度加快,而作为寄生者的x1在这两种情况下均保持持续增长趋势。这是因为中间主体与边缘主体的竞争激烈程度同时增大或减小时,由于受到两方边缘主体强烈的空间挤占与资源掠夺,中间主体即使占据结构洞位置也难以发挥位置优势获得增长,同时结构洞的存在使得寄生者可以从宿主处汲取资源以促进自身发展,宿主因面临寄生者的不断消耗与中间主体的持续抑制而出现创新产出低迷的状态。
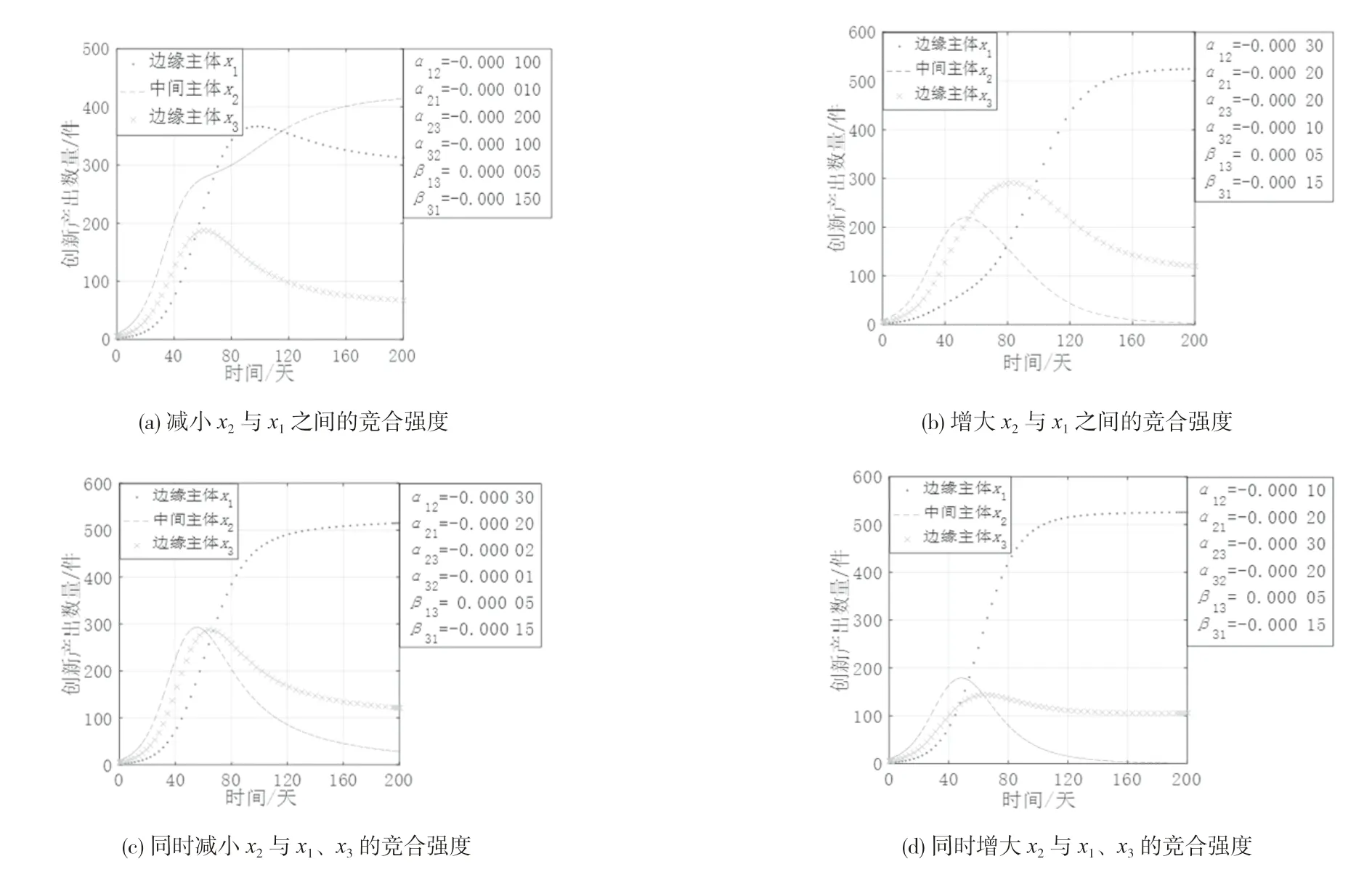
图5 创新生态系统中中间主体与边缘主体之间竞合强度变化的演化态势
4 实证研究
4.1 数据选取
本研究选取索尼、爱立信、三星等3 家企业在移动通信领域的授权发明专利数据进行实证分析。爱立信作为全球通信业巨头,其业务范围覆盖了网络设备、移动终端、专业技术服务等,以先进的宽带码分多址技术、LTE 技术及强大的移动系统等关键核心技术与其他多家企业建立了技术合作关系,其中就包括与索尼、三星间的合作[15]。索尼与三星同作为综合性跨国企业集团,业务范围均十分广泛,但这两家企业的业务范围同时涵盖了移动通信领域,且均存在外在产品和服务的激烈竞争,如移动电话、笔记本电脑、数码摄像机等[16-17]。资料显示,直到2004 年12 月,索尼与三星才签署了相互授权共享彼此大部分专利的重要协议,以减少不必要的专利技术之争和摩擦[18]。可见,2004 年之前索尼与三星之间并不存在直接联系,却分别与爱立信存在技术方面的合作,因此,爱立信在索尼与三星之间充当了中间人的角色,为这两家企业搭建起了桥梁,使得索尼与三星间建立起可相互传递间接影响的途径。从关系缺失的角度来看,爱立信占据了结构洞位置,索尼与三星分别处于结构洞边缘位置。在移动通信领域,这3 家企业既有技术上的合作也有外在产品上的竞争,其技术资源含量如图6 所示,A 表示3 家企业拥有的相同或相似的产品或技术服务,属于重叠资源,是引起3 家企业竞争的主要原因;B、C、D 分别表示这3 家企业两两之间相互重叠的资源;空白区域为互补资源。
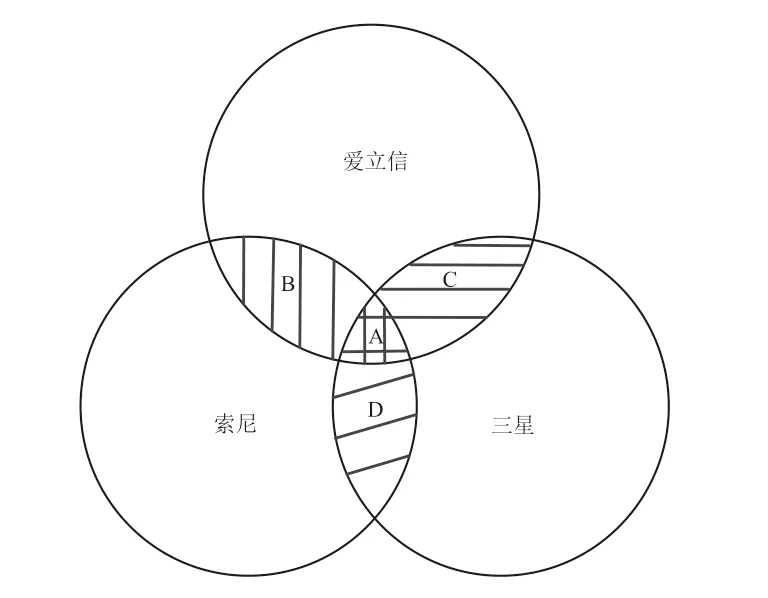
图6 索尼、爱立信、三星公司的技术资源含量
选取1996—2004 年上述3 家企业在移动通信领域的授权发明专利数量进行实证分析,其发展趋势如图7 所示。
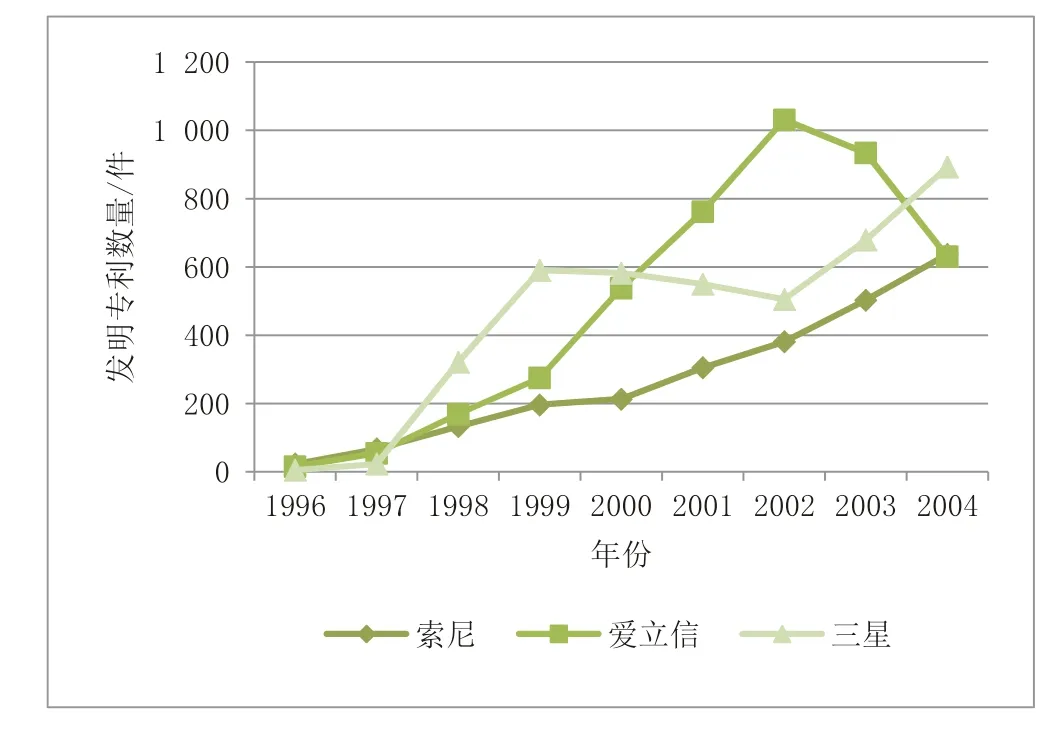
图7 案例企业授权发明专利数量年度分布趋势
4.2 参数计算
为探究在创新生态系统中索尼、爱立信、三星3 家企业之间的生态关系,采用灰色估计法计算三者的竞合系数及间接作用系数,模型式(1)可简化为一般形式如下:
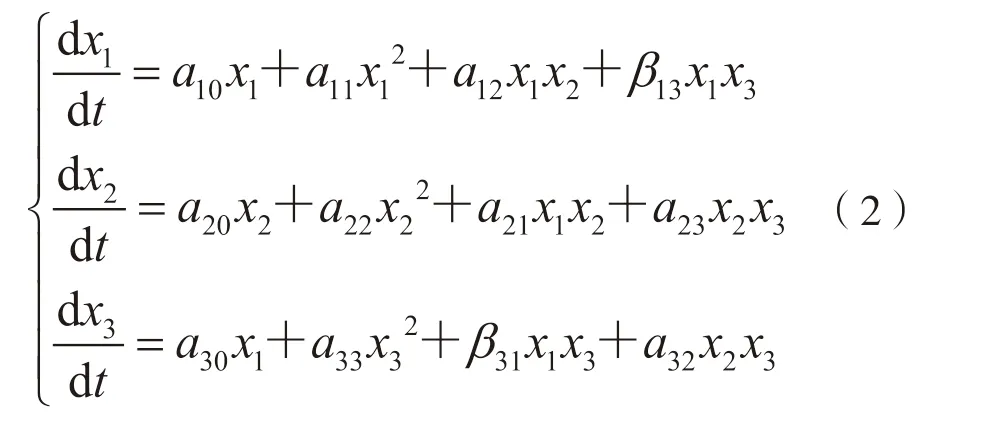
基于灰导数和偶对数的映射关系,有以下关系式:

根据最小二乘法准则,将原始数据代入求得各参数形成的系数矩阵S为:

则得出各创新主体的增长率分别为:r1=0.792 939,r2=1.197 306,r3=2.062 598。从各参数的符号可发现,索尼与爱立信、三星与爱立信之间的竞争强度大于合作强度,均表现为竞争共存关系,而三星与爱立信之间因结构洞的存在表现出寄生共生关系,且索尼为宿主、三星为寄生者,三星作为寄生者对索尼具有抑制作用,索尼作为宿主对三星具有促进作用。
将各参数输入MATLAB2016a 软件进行数值分析,如图8 所示,仿真所得3 家企业授权发明专利数量演化趋势与图7 中的实际增长趋势基本吻合,可见模型的拟合效果较好。因此,借鉴Lotka-Volterra 模型来研究创新生态系统中三者之间的生态关系可行的。
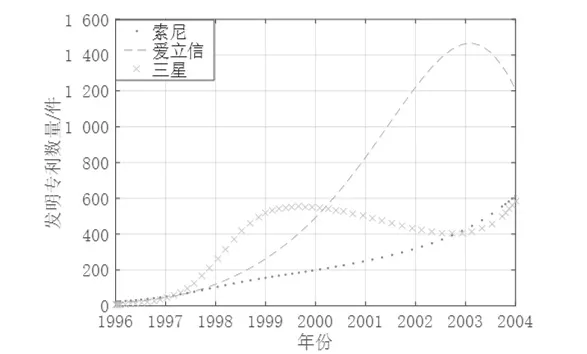
图8 案例企业发明专利数量仿真演化趋势
4.3 多主体创新产出演化分析
由上述计算结果可知,作为中间主体的爱立信同时与索尼、三星存在竞争共存的生态关系,且关系强度存在差异;索尼与三星之间为寄生共生的生态关系。下面将通过调整不同参数以探究这3 家企业在生态关系与位置特性交互作用下的创新产出演化规律。
(1)增大宿主的间接影响。图9 是创新生态系统中其他参数不变,保持三星对索尼的间接影响不变,增强索尼对三星的间接影响时3 个主体的创新产出演化态势。从图9 可知,索尼与三星呈现共同进化的高速增长趋势,随着索尼对三星的间接影响增强,索尼与三星的增长趋势均逐渐减缓,爱立信呈现倒“U”型增长趋势。这是因为三星与索尼间寄生共生,索尼的创新成果促进了寄生者三星的增长,索尼受到三星的不断消耗呈现略低于三星的增长态势;占据结构洞位置的爱立信由于与处在结构洞边缘的三星和索尼同时存在竞争关系,难以获得生存和发展的空间,更容易被淘汰。
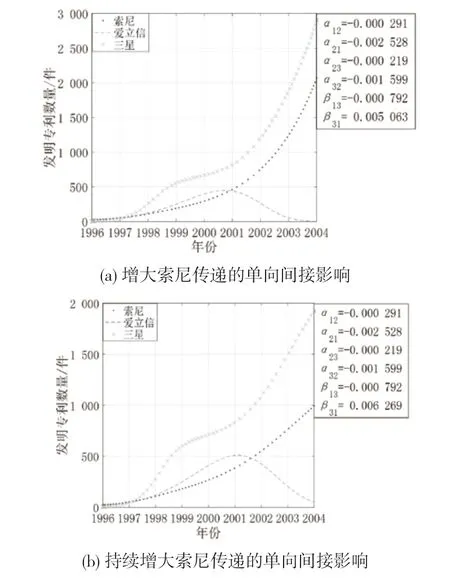
图9 增大宿主传递间接影响时案例企业创新产出的演化态势
(2)增大寄生者的间接影响。图10 是创新生态系统中其他参数不变,保持索尼对三星的间接影响不变,增强三星对索尼的间接影响时3 个主体的创新产出演化态势。从图10 可知,随着寄生者三星对索尼的间接影响增强,索尼的专利数量增长呈现出逐渐减缓趋势,后续增长动力不强;爱立信的专利数量增速显著上升,拐点位置发生偏移;三星的专利数量略增长后高速增长趋势骤降,难以生存。这是因为边缘主体与中间主体间竞争共存,一方面三星通过与爱立信的直接联系对索尼产生消耗性的间接影响,另一方面索尼需要面对来自直接合作伙伴爱立信的竞争压力,索尼处于低效运转状态;爱立信从中受益,逐渐占据上风;而三星本可不断消耗索尼而获得竞争优势,但占据结构洞位置的爱立信由于拥有信息优势和控制优势,其壮大速度远高于三星,因而三星受到的抑制作用增强,会被逐步挤出市场。
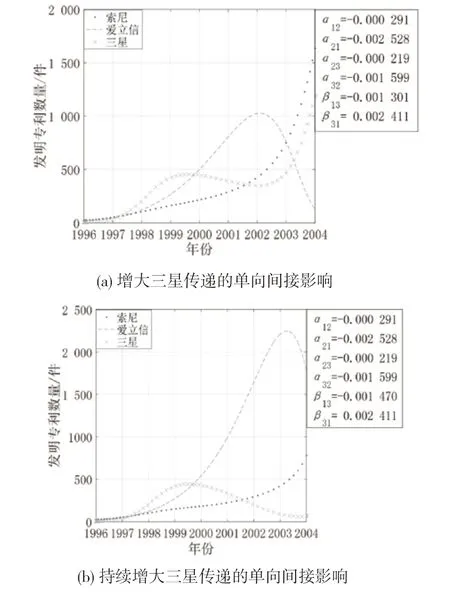
图10 增大寄生者传递间接影响时案例企业创新产出的演化态势
(3)同时增大寄生者与宿主的间接影响。图11 是创新生态系统中其他参数不变,三星和索尼对双方的间接影响同时增强时3 个主体的创新产出演化态势。从图11 可知,随着三星与索尼之间的间接影响同时增强,三星与索尼的增长趋势逐渐减缓,爱立信保持高速增长,其拐点位置发生偏移。这是因为边缘主体间寄生共生,寄生者三星通过与爱立信建立的直接联系对索尼产生的间接影响加剧了三星对索尼的消耗,索尼的增长受到抑制,同时索尼对三星的间接影响增强会促进三星的创新产出,但索尼的消亡减少了爱立信的增长阻力,爱立信获得较大的生存发展空间,进而抑制了三星的创新产出,最终出现爱立信一家独大的局面。
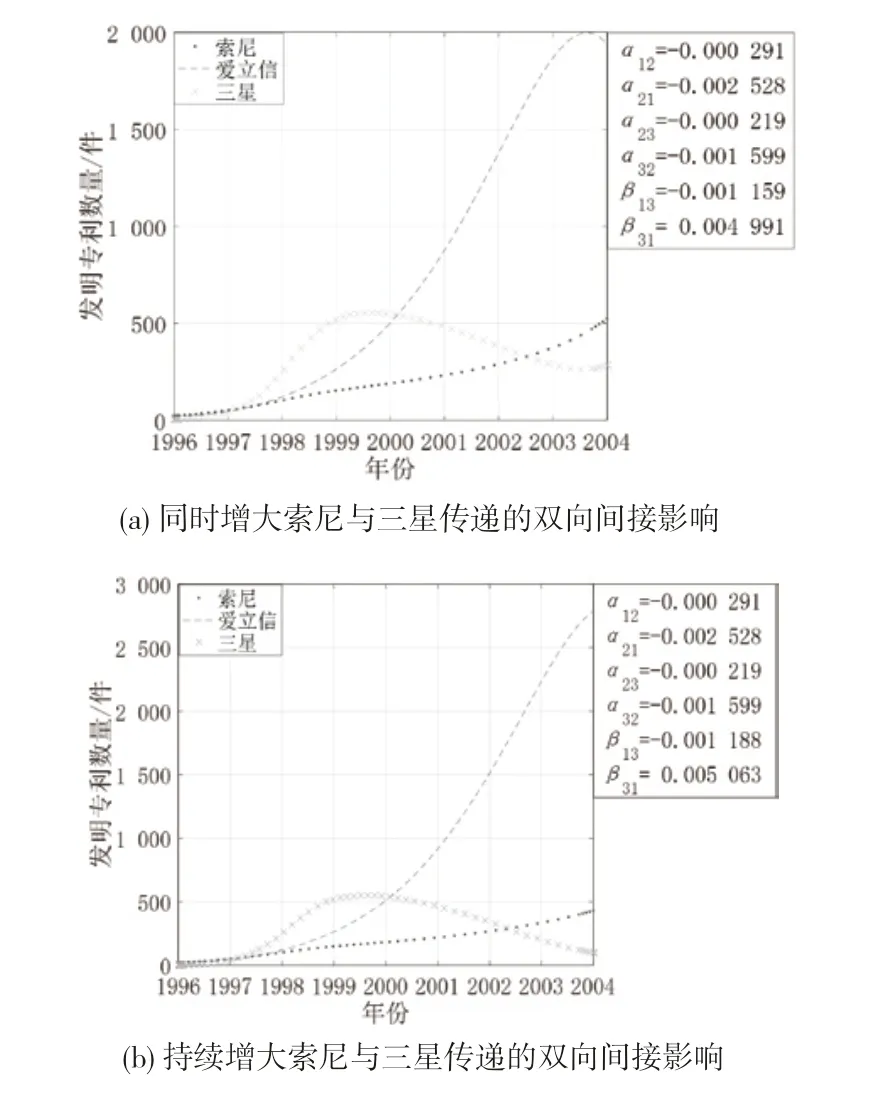
图11 同时增大寄生者与宿主传递间接影响时案例企业创新产出的演化态势
(4)中间主体与边缘主体的竞合强度变化。图12 是创新生态系统中其他参数不变,爱立信与索尼、三星的关系强度变化时3 个主体的创新产出演化态势。从图12 可知,索尼对爱立信的抑制作用减小时,爱立信的专利产出呈现持续增长的趋势,索尼与三星均呈现倒“U”型增长。这是因为爱立信占据结构洞位置,并与索尼、三星存在密切的技术互动,能够洞悉这两方的技术发展动态,因此当索尼减小对爱立信的抑制作用时,爱立信可以通过发挥自身的技术实力进行技术创新来提升自己的成长空间;而索尼与三星受到爱立信的挤压难以发展,虽然三星通过消耗索尼获得了专利产出的小幅增长,但总体创新产出较低。当爱立信与三星的竞争强度不断增大时,爱立信的专利产出增长呈现倒“U”型趋势,作为宿主的索尼的专利产出持续增长一段时间后开始逐步下降,作为寄生者的三星的专利产出呈现曲折上升的增长态势。爱立信与三星的竞争强度不断增大意味着爱立信不愿意放弃在市场上获得的收益,持续增大资源投入以获取竞争优势;同样的,三星也采取了相同的措施以牢牢把控市场份额,但三星还可以从宿主索尼处寻求创新资源进行发展,因此三星的专利产出增长速度最快。随着时间的推移,爱立信与三星之间的竞争持续激化,同时受索尼与三星抑制的爱立信的创新活力不断减退,其创新产出达到峰值后逐渐下降;索尼的专利产出也因此受到影响呈下降趋势;三星在爱立信的专利产出达到峰值时走势平缓,其在此阶段的专利产出与爱立信持平,伴随着爱立信的逐步消亡,三星重新获得生长空间,并从索尼处吸取了创新发展所需的资源,呈现高速增长的态势。
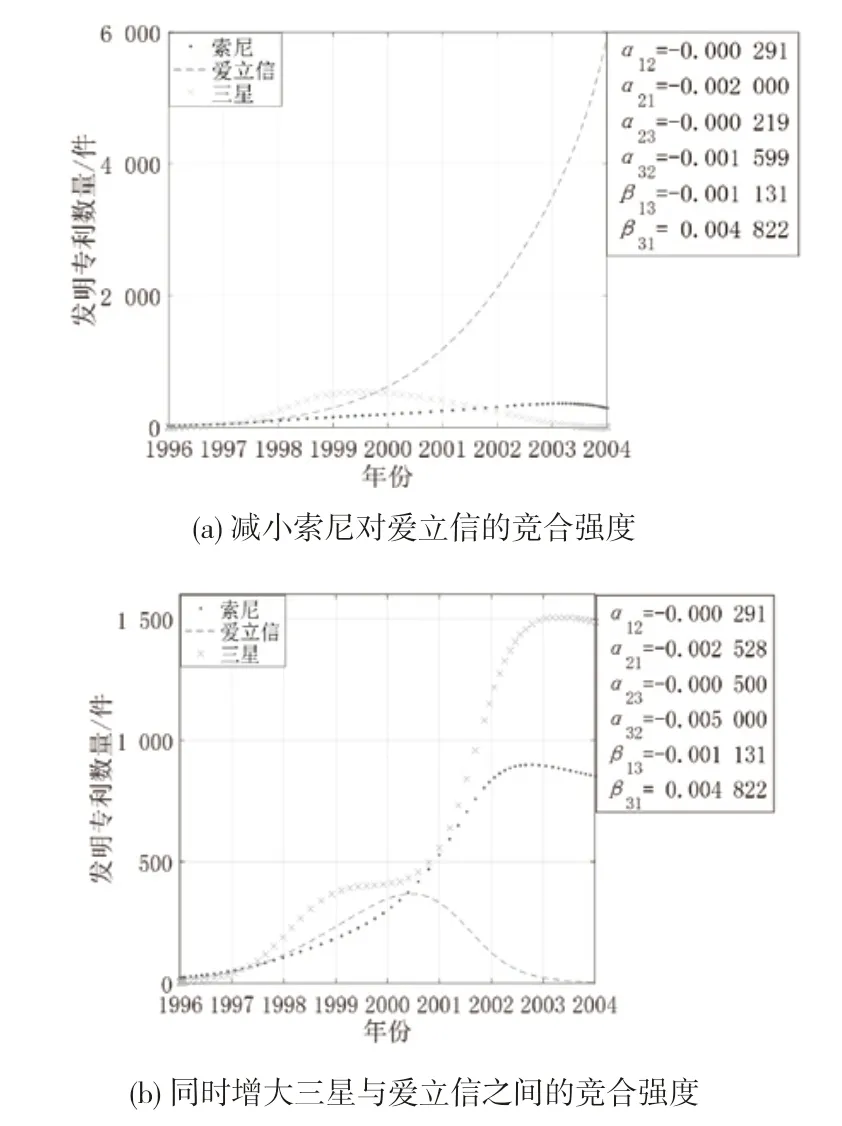
图12 案例企业竞合强度变化时的创新产出演化态势
5 结论与启示
本研究结合生态学理论与结构洞理论,综合考虑创新生态系统中创新主体间的直接生态关系与间接生态关系及关系强度变化的交互作用对各主体产生的影响,建立多主体创新产出模型,通过数值仿真模拟各创新主体创新产出演化态势,最后以索尼、爱立信、三星在移动通信领域的授权发明专利数据为例进行了实证分析,得出如下结论:
(1)竞争共存生态关系下结构洞边缘主体之间传递的间接影响严重削弱占据结构洞位置中间主体的创新产出,中间主体的生存和发展受边缘主体间生态关系及其自身与边缘主体间生态关系交互影响。中间主体虽然拥有获取更多异质性信息和资源的机会,但与结构洞边缘主体间保持竞争为主的生态关系会对自身发展起抑制作用。因此,占据结构洞位置的中间主体应注重开发合作伙伴必需的互补性资源,减少与合作伙伴在相似产品或服务方面的竞争摩擦,增强合作深度和广度,以促进自身与其他主体的良性增长。
(2)寄生共生生态关系下结构洞边缘主体之间传递的间接影响对各主体创新产出的作用效果存在较大差异,作为宿主的边缘主体传递单向间接影响会促进作为寄生者的边缘主体发展,作为寄生者的边缘主体传递单向间接影响会阻碍作为宿主的边缘主体发展,宿主和寄生者同时传递双向间接影响会阻碍双方发展。在案例创新生态系统中,三星作为寄生者对宿主索尼传递的间接影响阻碍了索尼的创新产出,而索尼作为宿主对寄生者三星的间接影响起促进性作用,但双方盲目传递间接影响会制约双方发展。因此,边缘主体应注意识别自身所扮演的生态角色,通过市场需求导向挖掘作为寄生者的边缘主体的创新潜力,加强主体内部自主创新程度,促进边缘主体间寄生共生模式向互惠共生模式演化,进而推动双方的协同发展。
(3)互惠共生生态关系下结构洞边缘主体之间传递的间接影响显著推动边缘主体双方的创新产出,边缘主体间传递双向间接影响对对方的促进作用明显强于双方传递单向间接影响时的促进作用。因此,边缘主体间应充分发挥互惠共生模式所带来的发展优势,并在此基础上提升双方发展战略的契合度,注重双方技术领域的多元化和异质化发展,增强双方的技术创新资源整合及价值创造能力,以保证双方的技术创新能力持续提升。
猜你喜欢
摄影世界(2020年12期)2020-12-06 10:48:57
小学生作文(低年级适用)(2020年10期)2020-11-10 09:12:12
中国建筑装饰装修(2020年6期)2020-07-10 09:41:16
福建基础教育研究(2019年2期)2019-09-10 07:22:44
福建基础教育研究(2019年2期)2019-05-28 08:39:49
家庭影院技术(2018年10期)2018-11-02 05:35:34
摄影之友(影像视觉)(2017年12期)2017-02-08 02:10:09
摄影之友(影像视觉)(2017年12期)2017-02-08 02:10:09
信息通信技术与政策(2016年11期)2016-02-21 20:33:37
卫星电视与宽带多媒体(2013年9期)2013-06-07 06:18:30
