
喜马拉雅山错那段不同海拔梯度蚂蚁多样性
2021-06-28 08:55:52许国莲徐正会李晓艳刘兰兰钱昱含
环境昆虫学报 2021年3期
刘 霞,许国莲,徐正会,张 翔,李晓艳,刘兰兰,钱昱含
(西南林业大学生物多样性保护学院,云南省森林灾害预警与控制重点实验室,昆明 650224)
蚂蚁隶属于膜翅目Hymenoptera细腰亚目Apocrita蚁科Formicidae,是自然界中种类和生物量最丰富的社会性昆虫,在生态系统中发挥着重要生态功能。据统计,全世界现存17亚科、337属、13 684种(Bolton, 2020)。由于蚂蚁的物种多样性及在所有能级水平上的生态功能都非常高、且易采集、对环境变化敏感,已经成为指示生物多样性和环境变化的关键物种(Hoffmann, 2010; 李巧,2015),以蚂蚁作为指示生物对生态恢复进行评价越来越受到重视。
喜马拉雅山是中尼边境上西北-东南走向的高大山系,也是世界最高的山脉,其高大山体对山脉两侧的气候分异、生物多样性演化产生了深刻影响,自约7 000万年前的古生代以来一直是众多特有物种的栖息地。对喜马拉雅地区的蚂蚁分类研究前人已有报道(Forel, 1906; Menozzi, 1939; Collingwood, 1982; 唐觉和李参,1982; Radchenko & Elmes, 1999)。蚂蚁多样性研究方面,已报道了印度境内喜马拉雅山区(Bharti & Shamma, 2009; Bhartietal, 2013)、中国境内喜马拉雅山亚东段(莫福燕等,2015)、珠峰段(李文琼等,2016)、普兰和吉隆段(张翔等,2018)、底雅和札达段(刘霞等,2020)以及藏东南地区(张成林等,2011;刘霞等,2011;于娜娜等,2012;刘霞等,2017)等蚂蚁多样性的研究,这些研究丰富了喜马拉雅山区的蚂蚁研究资料。错那段位于喜马拉雅山东段、雅鲁藏布江中下游地区,是藏东南温暖多雨地区向喜马拉雅山高寒干旱腹地过渡的典型的山原湖盆谷地,由南坡、北坡和山间谷地组成。相较于喜马拉雅山西段的底雅和札达与普兰和吉隆段、中段的珠峰段和亚东段,喜马拉雅山东段将演化出何种蚂蚁群落和多样性水平?为此,于2018年8月对喜马拉雅山错那段的蚂蚁多样性进行调查,以期为全面揭示喜马拉雅山蚂蚁群落结构和多样性规律提供科学依据。
1 材料与方法
1.1 样地设置
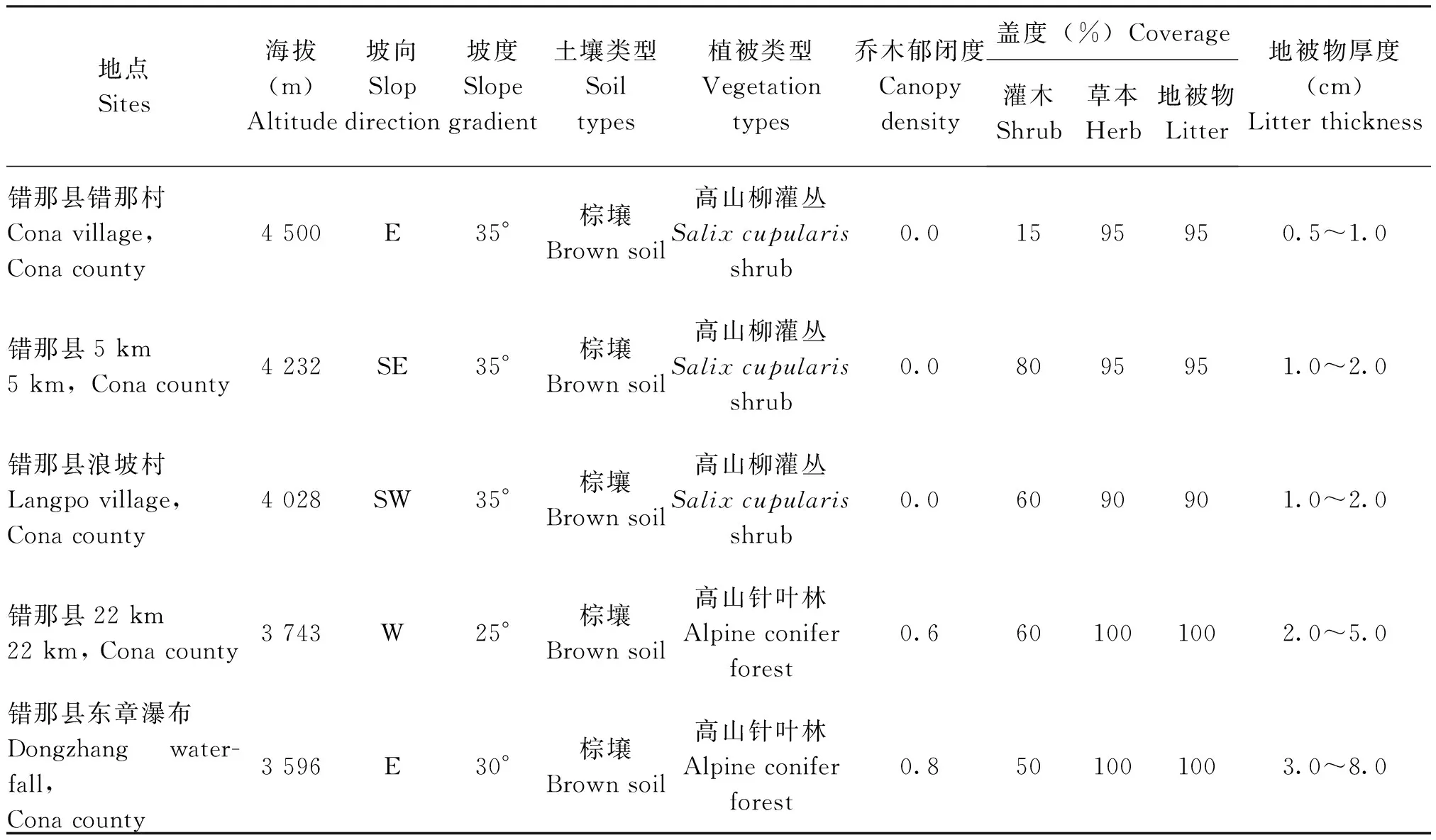
2018年8月,沿喜马拉雅山错那段由北向南进行调查,海拔每升高250 m设置1块50 m×50 m的样地,在3个垂直带共设置18块样地。其中北坡乃东垂直带(91°50′17″~92°03′16″E,28°49′51″~29°05′43″N),海拔3 755 m~5 016 m,设置6块样地;山间谷地曲松-隆子垂直带(92°02′16″~92°03′55″E,28°23′18″~28°48′40″N),海拔4 010 m~4 730 m,设置6块样地;南坡错那垂直带(91°57′05″~91°58′34″E,27°46′38″~28°13′01″N),海拔3 596 m~4 755 m,设置6块样地,各样地概况见表1。

表1 喜马拉雅山错那段蚂蚁群落调查样地概况
1.2 调查方法
采用样地调查法进行调查,在每块样地按照对角线五点取样设置5个样方,样方大小1 m×1 m,间距10 m,每个样方调查地表样、土壤样和树冠样的蚂蚁并采集标本(徐正会,2002)。样方调查结束后用搜索法对样地内样方外各生境中的蚂蚁进行调查与采集以补充样方调查,并计入物种数和个体总量,但不计入个体密度。将所采标本用95%乙醇保存于2 mL的冻存管内,做好记录带回实验室。同一巢穴内的蚂蚁保存于同一冻存管内,不同生境的蚂蚁分别保存于不同的冻存管中。依据主要分类学著作(吴坚和王常禄,1995;周善义,2001;徐正会,2002),采用形态分类学方法对标本逐一鉴定。
1.3 多样性指标测定及相关性分析
为了比较各垂直带不同海拔梯度蚂蚁群落的组成、物种丰富度及多样性,利用EstimateS 9.1.0软件对数据进行处理,蚂蚁物种多样性采用物种数目、个体密度、Shannon-Wiener多样性指数、Fisher α指数、Simpson优势度指数和Jaccard相似性系数进行测度(徐正会,2002;李巧等,2009),同时运用基于多度(Abundance-base Coverage Estimator, ACE)、基于频数(Incidence-base Coverage Estimator, ICE)、Chao1(for Individual-based Abundance Data)和Chao2(for Sample-based Incidence Data)的估计方法对物种丰富度进行估计(李巧,2011;Colwell & Elsensohn, 2014)。依据各物种个体数占总个体数的百分比确定物种的优势等级,等级划分:优势度≥10.0%,为优势种,用A表示;5.0%≤优势度<10.0%为常见种,用B表示;1.0%≤优势度<5.0%为较常见种,用C表示;0.1%≤优势度<1.0%为较稀有种,用D表示;优势度<0.1%为稀有种,用E表示(黄钊等,2019)。用Pearson相关分析方法(双侧检验)分析蚂蚁多样性(物种数目、多样性指数、优势度指数)与植被(乔木郁闭度、灌丛盖度、草本盖、地被物盖度、地被物厚度)的相关性(张念念,2014)。
续表1 Continued table 1
(1)Shannon-Wiener多样性指数
H=-∑PilnPi,Pi为第i种的个体数占群落全部物种个体数之和的比值。
(2)Simpson优势度指数

(3)Jaccard相似性系数
q=c/(a+b-c),c为两个群落的共有物种数,a和b分别为群落A和群落B的物种。
2 结果与分析
2.1 蚂蚁群落组成与优势度分析
在喜马拉雅山错那段18块样地内共采集蚂蚁7 516头,隶属于2亚科、5属、7种(含1待定种)。其中切叶蚁亚科2属、4种,蚁亚科3属、3种(表2);该地区蚂蚁群落优势种有4个,分别为西藏盘腹蚁(38.69%)、岩缝红蚁(15.35%)、光亮黑蚁(29.24%)和箭蚁待定种1(12.40%),其中西藏盘腹蚁的优势度最大;较常见种2个,为鲁氏红蚁(2.61%)和西姆森弓背蚁(1.68%),亮红蚁(0.03%)是该地区最稀有的物种。从优势种的成分来看,都是适应高海拔耐寒的物种。

表2 喜马拉雅山错那段蚂蚁种类、个体数及优势度
2.2 蚂蚁群落主要指标分析
2.2.1物种累计曲线
随着调查样地数量的增加,ICE值先急剧上升,随后急剧下降,然后缓慢下降,最后趋于水平,实际观察物种数(Sobs)、ACE、Chao1和Chao2值先急剧上升,后缓慢上升,最后趋于水平(图1)。蚂蚁物种的实际观察值(Sobs为7)与丰富度的估计值(ACE为7;ICE为7.37;Chao1为7;Chao2为6.94)相等或接近,说明喜马拉雅山错那段蚂蚁物种丰富度相对较低。实际采集到的物种数约为估计值的94.98%~100.00%,样地中多数物种被采集到,抽样效果好。

图1 喜马拉山错那段蚂蚁物种实测值和预测值累积曲线Fig.1 Accumulation curves for observed and estimated number of ant species at Cona section of Mt. Himalaya注:Sobs表示实际观察的物种数;ACE表示基于多度(个体数量)的预测值;ICE表示基于频数(发生率)的预测值;Chao1、Chao2表示用Chao1、Chao2方法预测值。Note: Sobs indicated number of species observed; ACE indicated abundance (individual)-base coverage estimator; ICE indicated incidence-base coverage estimator; Chao1 and Chao2 indicated Chao1 and Chao2 estimator.
2.2.2物种数目
从物种的实测值来看,喜马拉雅山错那段北坡、山间谷地和南坡3个区域蚂蚁的物种数都接近或等于估计值ACE和Chao1值,说明这3个区域内的蚂蚁物种丰富度低(表3)。其中北坡山顶5 016 m的草丛样地和南坡海拔4 500 m高山柳灌丛、4 755 m草丛样地中未发现有蚂蚁活动。
错那段3个垂直带蚂蚁物种数目顺序为:北坡(7种)>山间谷地(5种)>南坡(3种)。北坡各样地物种数目在0~4种之间,物种数目随着海拔升高先升高后降低,峰值出现在山坡中下部海拔4 009 m的草丛和4 264 m的灌丛样地中(4种)。山间谷地各样地物种数目在1~4种之间,物种数目随海拔升高呈现先升高后降低的规律,但在坡面顶部海拔4 730 m的草丛样地中又出现第二峰值。南坡各样地物种数目在0~2种之间,蚂蚁物种数目随海拔升高总体呈现降低的规律,但在海拔4 028 m高山柳灌丛出现第二峰值(2种)。
2.2.3个体密度
错那段3个垂直带上蚂蚁平均个体密度顺序为:北坡(12.3头/m2)>南坡(5.4头/m2)>山间谷地(3.5头/m2)。北坡各样地个体密度为0.0~23.4头/m2,个体密度随海拔升高基本呈现先升高后降低的规律,在海拔4 264 m的灌丛(23.4头/m2)和4 749 m的灌丛(18.8头/m2)样地出现2个峰值。山间谷地各样地个体密度在0.6~6.4头/m2之间,个体密度在海拔4 010 m草丛、4 243 m灌丛和4 502 m灌丛样地出现3个弱的峰值。南坡各样地个体密度在0.0~22.6头/m2之间,个体密度随海拔升高依次出现2个峰值。
2.2.4多样性指数
错那段3个垂直带Shannon-Wiener多样性指数顺序为:北坡(0.9430)>山间谷地(0.5383)>南坡(0.3031)。随海拔的升高,北坡的多样性指数先升高后降低,再升高后降低,在海拔4 009 m草丛(0.5545)和海拔4 496 m草丛样地(0.6167)中出现2个峰值;山间谷地的Shannon-Wiener多样性指数随海拔升高先升高后降低,再升高,在海拔4 243 m灌丛(0.8634)和4 730 m草丛(0.7926)样地中出现2个峰值;南坡的Shannon-Wiener多样性指数最高值出现在坡面中部海拔4 028 m高山柳灌丛样地。
在3个垂直带上,Fisher α指数顺序表现为:北坡(0.84±0.10)>山间谷地(0.58±0.08)>南坡(0.37±0.07)。北坡的Fisher α指数随海拔升高先升高后降低,峰值出现在海拔4 009 m草丛样地中(0.54±0.09)。山间谷地的Fisher α指数随海拔升高先升高后降低,再升高,在海拔4 243 m灌丛(0.55±0.10)和4 730 m草丛(0.56±0.14)样地出现两个峰值。
2.2.5优势度指数
在错那段3个垂直带上,优势度指数顺序为:南坡(0.8391)>山间谷地(0.7682)>北坡(0.4846),与物种数目、Shannon-Wiener多样性指数和Fisher α指数顺序相反。北坡各样地优势度指数随着海拔的升高,依次在海拔3 755 m矮灌丛(0.8290)、4 264 m灌丛(0.9520)和4 749 m灌丛(0.9609)样地中出现3个峰值。山间谷地各样地优势度指数随海拔升高依次在海拔4 010 m草丛(0.9388)、4 486 m草丛(1.0000)和4 502 m灌丛(1.0000)样地中出现3个峰值。南坡各样地的优势度指数随海拔的升高依次出现2个峰值。
2.3 蚂蚁多样性与植被的相关分析
喜马拉雅山错那段蚂蚁多样性与植被的相关性如表4所示,其中多样性指数与草本盖度(P<0.01)极显著正相关,与乔木郁闭度、灌木盖度、地被物盖度和地被物厚度相关性不显著;优势度指数与灌木盖度(P=0.046)显著相关,与乔木郁闭度、草本盖度、地被物盖度和地被物厚度相关性不显著;物种数目与植被相关性不显著。

表4 蚂蚁多样性与植被特征相关分析
2.4 蚂蚁群落相似性
根据Jaccard相似性原理,当0.00≤q<0.25,两群落间极不相似;当0.25≤q<0.50,两群落间中等不相似;当0.50≤q<0.75,两群落间中等相似;当0.75≤q≤1.00,两群落间极相似。错那段北坡、山间谷地和南坡蚂蚁群落间相似系数为0.4286~0.7143,处于中等不相似至中等相似水平;平均值0.5810,显示中等相似水平。其中南坡与北坡间达到中等不相似水平,北坡与山间谷地、山间谷地与南坡间处于中等相似水平(表5)。可见,喜马拉雅山错那段各垂直带蚂蚁群落间的相似性较高,群落间分化不明显。

表5 喜马拉雅山错那段各垂直带蚂蚁群落间相似性系数(q)
3 结论与讨论
在以往山地蚂蚁研究中,蚂蚁物种丰富度大多呈现南坡>北坡(莫福燕等,2015;李文琼等,2016;刘霞等,2017;张翔等,2018;刘霞等,2020)、东坡>西坡的规律(徐正会等,2001a,徐正会等,2001b;于娜娜,2012),主要是由水热条件和坡面长短导致。在本研究,蚂蚁物种丰富度北坡(7种)>南坡(3种),这是因为南坡处于西南季风的迎风坡,降雨多,土壤湿度过大,且坡面较短,北坡处于背风坡,降雨相对较少,土壤相对干燥,蚂蚁喜欢温暖干燥、阳光充足的环境,因此南坡的蚂蚁物种丰富度低。
喜马拉雅山错那段位于藏东南地区向喜马拉雅山腹地过渡的谷地,在海拔3 596~5 016 m的范围内(高差1 420 m)共采集蚂蚁2亚科5属7种,其物种丰富度与靠近欧亚内陆的喜马拉雅山西段的底雅和札达段相同(2亚科5属7种,海拔2 975~5 183 m,高差2 208 m)(刘霞等,2020),但明显低于西段的吉隆(3亚科9属14种,海拔2 742~5 005 m,高差2 263 m)(张翔等,2018)、中段的亚东(2亚科8属12种,海拔2 760~5 049 m,高差2 289 m)(莫福燕等,2015)和珠峰段(3亚科13属23种,海拔1 840~5 260 m,高差3 420 m)(李文琼等,2016),更显著低于同纬度藏东南的德姆拉及察隅河谷(5亚科31属95种,海拔1 590~4 776 m,高差3 186 m)(于娜娜,2012)和嘎隆拉及墨脱河谷(8亚科45属96种,海拔740~4 208 m,高差3 468 m)(刘霞等,2017)。在喜马拉雅山区,自东南向西北,降雨依次减少,气候由亚热带半湿润气候向高原温带干旱、寒带干旱气候过渡。靠近欧亚内陆的底雅和札达,由于降水少、气候寒冷干燥,影响了蚂蚁物种丰富度,只有对极寒气候适应的物种分布。错那段与亚东段、珠峰段和吉隆段的纬度相近,但蚂蚁物种丰富度最低,这主要是由错那段的海拔高差小、最低海拔高及降雨多造成的。可见,降水和海拔高差对物种丰富度有重要影响。此外,凋落物的质量对蚂蚁物种丰富度和多样性有影响,凋落物质量越高,森林生态环境越好,其种群越丰富(Theunisetal., 2005)。喜马拉雅山亚东段、吉隆段、珠峰段、藏东南地区德姆拉及察隅河谷、嘎隆拉及墨脱河谷样地凋落物和生态环境总体上优于错那段,导致错那段的物种丰富度和多样性相对低。
在过去的研究中,蚂蚁物种多样性沿海拔梯度变化规律大致呈三种类型:(1)蚂蚁多样性随海拔升高而降低,可用Rapoport效应假说解释(Janzenetal., 1976; 徐正会等,2001a;Majeretal., 2001; Colwelletal., 2008; Kwonetal., 2014);(2)物种多样性随海拔升高呈单峰曲线,可用“中域效应”(Mid-domain effect)解释(Fisher, 1996; Samsonetal., 1997; Sanders, 2002);(3)蚂蚁多样性随海拔升高而升高(Sandersetal., 2003)。已报道的藏东南地区色季拉山的蚂蚁多样性研究支持Rapoport效应假说(张成林等,2011);印度境内喜马拉雅山区、中国境内喜马拉雅山亚东段蚂蚁多样性研究支持“中域效应”(Bhartietal, 2013;莫福燕等,2015)。珠峰段、吉隆段的蚂蚁群落研究显示,在坡面上不仅出现1个峰值,而是出现多个峰值,这种现象被称为多域效应(Multi-domain effect),多域效应是中域效应在山体坡面上的重复出现(李文琼等,2016;张翔等,2020)。在本研究中,喜马拉雅山错那段3个垂直带上,物种数目、个体密度和多样性指数随海拔升高总体呈现垂直带中、下部较高,垂直带上部较低,说明海拔对多样性具有重要影响。北坡蚂蚁物种数目随海拔升高呈单峰曲线,表现出中域效应现象,其他各指标随海拔的升高,在坡面上出现2个或2个以上的峰值,更多的表现出多域效应现象。
在海拔主导因素的作用下,蚂蚁群落各指标可能还受地形因子的影响。地形因子主要包括坡度、坡位和坡向(王移等,2010),坡度、坡位和坡向不同,接收的热量和降水条件不同,可影响具体样地的小生境,进而影响蚂蚁的物种丰富度和多样性。其中坡度还影响土壤的含水量和养分流失程度(Zingg,1940),进而对土壤动物的群落结构、多样性及主要类群的密度产生影响(何先进等,2012),蚂蚁作为重要的土壤节肢动物,其群落结构、多样性和个体密度也会受到坡度的影响。在喜马拉雅山错那段,蚂蚁物种丰富度、多样性指数和个体密度总体上随坡度的增大而降低。
地表植被也是影响蚂蚁种群的重要因素之一。植被是微气候生境的重要调节者,主要改变了温度而直接影响了蚂蚁的分布、代谢和觅食(Jensen & Hoolm, 1980; Nielsen, 1986; Lópezetal., 1992)。有研究表明植被结构简单的栖境中蚂蚁物种数少,从而降低了蚂蚁物种多样性(Pacheco & Vasconcelos, 2012),而良好的植被状况能够为更多蚂蚁物种提供更丰富和更优质的食物资源、筑巢地及隐蔽地(Buczkowski & Richmond, 2012)。在本研究,蚂蚁多样性指数与草本盖度显著相关,优势度指数与灌木盖度显著相关。较好的草本盖度给蚂蚁提供了良好的筑巢环境,有利于蚂蚁的繁殖;较高的草本盖度说明样地内人畜活动少,受干扰小,有利于蚂蚁种群的繁衍扩大。较多的灌木为蚂蚁提供了更多的栖居和觅食场所,并可躲避其他捕食者的捕食,使能爬上灌木的蚁种优势地位突出。
从相似性系数来看,错那段3个垂直带间蚂蚁群落相似性较高,达到中等相似水平。喜马拉雅山错那段除南坡少数样地的植被为针叶林外,北坡、南坡和山间谷地的植被多为灌丛和草地,生态环境较简单,且最低海拔超过3 500 m,海拔落差小,其间栖息的蚂蚁物种多数是适应高海拔环境的古北界物种,因此3个垂直带蚂蚁群落间相似性较高。可见,植被和海拔高差对蚂蚁群落间相似性有重要作用。
总体而言,喜马拉雅山错那段的蚂蚁物种丰富度和多样性较低,主要受降水、海拔高差、坡度和植被的共同影响。作为能较好指示生物多样性和环境变化的关键物种(Hoffmann, 2010; 李巧,2015),从蚂蚁的物种组成和分布来看,该地区的生物资源十分有限,环境脆弱,因此有必要采取措施保护该地区的环境和生物多样性。
猜你喜欢
今日农业(2022年1期)2022-11-16 21:20:05
安徽文学(2020年10期)2020-10-26 06:57:16
书香两岸(2020年3期)2020-06-29 12:33:45
户外探险(2019年7期)2019-07-16 04:14:55
石油地球物理勘探(2017年2期)2017-11-23 06:01:41
广东农业科学(2017年5期)2017-08-29 10:37:54
西藏研究(2016年5期)2016-06-15 12:56:33
文学少年(原创儿童文学)(2016年19期)2016-02-28 17:50:40
清明(2014年1期)2014-06-28 15:16:11
长江文艺·好小说(2014年3期)2014-03-04 20:32:16
