
次生落叶阔叶林林隙的紫椴造林试验1)
2021-06-26 01:50李佳明郑金萍郭忠玲范春楠王圣钧
东北林业大学学报 2021年6期
李佳明 郑金萍 郭忠玲 范春楠 王圣钧
(北华大学,吉林省·吉林市,132013)
林隙是由于树木死亡或环境因素而形成的林冠空隙[1]。林隙的形成与闭合会引起光环境的剧烈变化。处于幼苗期的植物通常对环境胁迫表现极为敏感,苗龄越小其敏感性越大[2]。由于林隙大小、形状以及边缘木种类等的不同,其内部的光照、温度、水分等因素存在差异,进而影响苗木的定植、生长、发育、成活,并对森林群落的结构、功能、多样性维持有重要影响[3-4]。不同大小的林隙,光照强度随着林隙面积的增大而增大[5],并由林隙中心区域向林缘逐渐降低[6]。林下幼苗一般生长在相对庇荫的环境中,随光照强度的增加,植物的光合能力也逐渐增强,但过高的光照强度会导致其发生光抑制[7]。Meer et al.[8]、Zhu et al.[9]的研究发现,随林隙面积的增加,更新苗的苗高、地径逐渐增大。也有研究表明,当达到一定面积后,林隙面积对幼苗的影响较小[10]。
紫椴(Tiliaamurensis)是椴树科(Tiliaceae)椴树属落叶乔木,树型优美、木材轻软,是优良的蜜源、药用植物[11],是长白山、小兴安岭阔叶红松混交林、落叶阔叶林的主要伴生树种之一[12]。受多年过度采伐,生境破坏、退化等因素影响,紫椴资源急剧减少,可利用蓄积几近枯竭。为此,在进行天然资源保护的同时,营造紫椴人工林受到高度重视[13]。紫椴对光照环境有着较强适应能力,天然更新幼苗在林冠下有典型的趋光现象,在全光环境时,则同时表现出一定程度的趋光、避光行为[14]。在庇荫环境下,紫椴一般采取演替先锋种的“避荫”对策,侧枝则采取中等耐荫种的“掠光”对策。弱光环境时,紫椴幼树一般有典型的忍耐适应行为[14]。本研究以吉林省中部山区典型次生落叶阔叶林为对象,开展林隙光环境的分区造林试验,通过对幼苗成活率、高径生长和生物量的测定,探讨光因子对紫椴幼苗定植、生长的影响,旨在保护、恢复紫椴资源,并为提高紫椴冠下人工更新技术提供参考。
1 研究区概况
研究区位于吉林市龙潭区东南部江密峰镇新山村,与龙潭区江北乡、天太镇管理区交界。地理坐标为126°39′47.81″E,44°0′41.71″N。属中温带亚湿润季风气候,多年平均气温3.9 ℃,1月份气温最低,一般在-18~-20 ℃,7月份气温最高,一般在21~23 ℃,最高气温可达36.6 ℃,年降水量700 mm左右,雨热同季,无霜期120 d,日照时间2 400~2 600 h。
试验区植被属于长白山植物区系,类型多样,结构复杂。林分起源以天然林为主,且绝大多数为反复干扰的天然次生林。研究选取以蒙古栎(Quercusmongolica)为优势的次生落叶阔叶林,林分年龄为中龄林,林分郁闭度0.75,平均胸径13.7 cm。主要伴生树种为胡桃楸(Juglansmandshurica)、山杨(Populusdavidiana)、色木槭(Acermono)、櫰槐(Maackiaamurensis)等;常见灌木以卫矛(Euonymusalatus)和山楂叶悬钩子(Rubuscrataegifolius)为主;草本植物有羊胡子薹草(Carexcallitrichos)、丝引薹草(Carexremotiuscula)、莫石竹(Moehringialateriflora)、玉竹(Polygonatumodoratum)、铃兰(Convallariamajalis)等。试验区珍贵阔叶树种,如紫椴、水曲柳(Fraxinusmandshurica)、黄檗(Phellodendronamurense)等,则因长期人为干扰、生境破坏几近消失。试验区土壤类型为暗棕壤,主要是腐殖质积累或由于弱酸性淋溶形成,表层有机质较多,结构疏松,排水性好。
2 研究方法
2019年5月,随机选择次生落叶阔叶林内15个面积大小不一的林隙,以紫椴1年生实生苗为材料,开展林隙造林试验。苗木栽植以林隙中心为起点,株距0.5 m,分别向东、南、西、北4个方向进行栽植,共计植苗401株。为获得各林隙每株苗木栽植点的光照强度,于7月10日(天气晴好),利用数字式照度计对各林隙每株苗木栽植点位光照强度进行测定。测定由早7点至晚6点止,每隔1 h测量1次。为尽可能保证光强测定的准同步性,利用5台照度计进行分组测量,每个林隙尽量在10 min内完成。通过各点光照强度日平均值的聚类分析进行光强分区。
紫椴苗木生长状况的调查于2019年7、8、9月分别进行了3次测定,内容包括株高、地径、成活率等。于9月末在各林隙的不同光照强度分区中,随机选择苗木9株,采用全收获法将苗木挖出,带回室内经清洗、分选(根、茎、叶)、烘干、称质量后,用于生物量及其分配特征分析。
试验数据统计分析采用SPSS 23.0完成,经方差分析、多重比较等,检验不同光照强度的紫椴苗木株高、地径、成活率之间的差异显著性。图形绘制利用Origin Pro 2019完成。
3 结果与分析
3.1 林隙光照环境
对各林隙长、宽调查数据进行整理,统计其面积。随机选取的15个林隙,面积均在22.46~84.06 m2。其中,林隙面积7个为20~30 m2;5个为30~50 m2;仅有3个在50 m2以上,且均未超过100 m2。按照林隙面积的划分方法[4],以上林隙属于小林隙。因各林隙在约10 hm2的范围随机选取,表明研究选区的次生落叶阔叶林以小林隙为主。
对各林隙苗木栽植点位日光照强度测定结果进行统计,经聚类分析将其划分为3个等级。其中,将光照强度介于400~2 250 lx定为弱光区,包含苗木222株;光照强度2 280~3 700 lx定为中光区,含苗木145株;光照强度3 700~5 300 lx定为强光区,仅含苗木36株。通过各林隙3个光照强度分区栽植苗木数量所占比例的统计,发现随林隙面积的增加,各光照强度分区紫椴苗木比例并未表现明显变化趋势。其中,7个林隙弱光区的苗木比例超过了50%;6个林隙中光区的苗木比例在50%以上;仅有1个林隙(面积为28.27 m2)强光区的苗木比例超过40%(表1)。这与所选的林隙均为小林隙,其受林隙走向、周围树种、枝叶稠密度、树高等因素的强烈影响。比较各分区与空地的光照强度(4 418 lx),各分区的光照强度较小,其中弱光区仅为全光的1%~5%、中光区为5%~8%、强光区为8%~12%。

表1 不同林隙面积各光照分区苗木所占比例
3.2 紫椴成活率
受区域2019年冬季降雪、春季干旱(1—4月降水量仅为2018年同期的25.40%)等因素影响(数据源自中国气象数据网数据库,站台号码为54172),紫椴苗木造林后,出现了较严重的枯梢现象。虽5—9月份的降水缓解了苗木枯梢(5—9月降水量为2018年的140.01%),但成活率仍受到了一定程度的影响,呈偏离正态分布。对数据进行反正弦转换处理、方差分析[15],结果见表2。
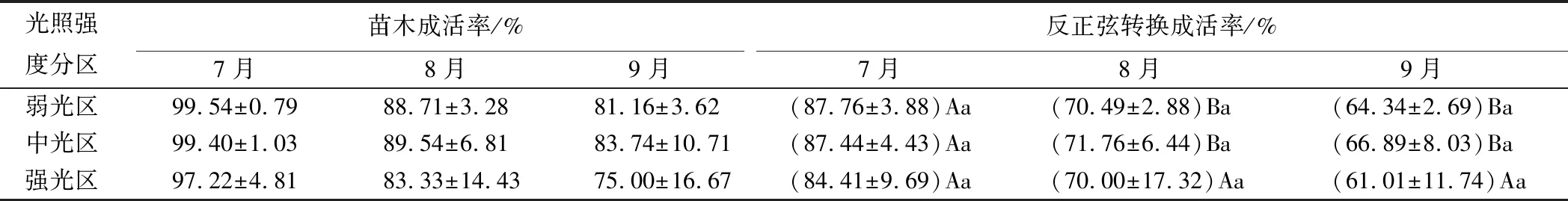
表2 不同光照强度分区的紫椴成活率反正弦转换
各光照分区的紫椴成活率均随生长推移而逐渐降低。至生长季末,中光区的苗木成活率相对较高,弱光区次之,强光区最小;中光与强光区间相差6.16%。但方差分析表明,各月份不同光照强度分区的成活率间并无显著差异。对于不同光照强度分区在各月份间的成活率,弱光区、中光区7月成活率显著高于8、9月,其8月的反正弦转换成活率分别较7月下降了17.27%、15.68%,9月成活率较8月下降了5%左右;强光区的成活率在各月间差异不显著,下降比例分别为14.41%、8.99%。说明中光、弱光区苗木的死亡较为集中,前期多后期少;强光区的苗木成活率则逐渐降低。综合分析可知,中光区、弱光区更利于苗木的成活。
3.3 紫椴株高、地径的生长状况
通过各光照强度分区的紫椴株高、地径月生长量以及年生长量统计,获得其株高、地径生长情况(表3、表4)。对于紫椴株高年生长量,强光区相对较高(7.51 cm),其次为弱光区(6.43 cm)、中光区(6.16 cm)。虽强光区的紫椴株高年生长量分别较弱光区、中光区分别提高了16.80%、21.92%,但多重比较发现,三者之间均无显著差异(P>0.05)。从各光照区的紫椴株高月生长量动态来看,8月份均显著高于7、9月。强光区的株高月生长量最大(5.26 cm),显著高于中光区(3.19 cm),弱光区次之(4.07 cm);中光区的株高月生长量7、9月略高,分别为1.56、1.41 cm,但各光照区间的差异均不显著。

表3 不同光照强度分区的紫椴株高生长量
中光区的紫椴地径年生长量(3.20 mm)显著高于强光区(2.93 mm)、弱光区(2.89 mm),分别提高了9.22%、10.73%。紫椴地径月生长量在弱光区、强光区均随生长的推移而降低,中光区随生长的推移而增加。对于各光照区在不同月份的地径月生长量,弱光区7、8月显著高于9月;强光区7月显著高于8、9月;中光区9月高于7、8月,但无显著差异。对于各光照区在相同月份的地径月生长量,7月强光区显著高于中光区;8月以弱光区相对略高,但三者间均无显著差异;9月中光区显著高于弱光区、强光区(表4)。表明弱光区、强光区有利于紫椴地径的初期生长,中光区则促进了紫椴地径在生长末期的积累。

表4 不同光照强度分区的紫椴地径生长量
统计各光照强度分区的紫椴高径比发现,高径比随生长的推移而逐渐下降。其中,弱光区、中光区7、8月的高径比显著高于9月;强光区仅7月高径比显著高于9月。对于不同光照强度分区相同月份的高径比,仅8月表现为中光区显著高于弱光区、强光区,原因是中光区的株高月生长量低于弱光区、强光区;但至9月生长季末,各光照区间无显著差异(表5)。可见各光照分区紫椴的径生长量比高生长量的累积晚,但光照强度未显著改变其比例关系。

表5 不同光照强度分区的紫椴高径比
3.4 不同光照强度紫椴生物量及分配比例
为进一步比较不同光照强度分区对紫椴生长分配的影响,根据生物量统计,发现不同光照强度分区的紫椴各器官、地上、整株生物量均表现为中光区最高。中光区的整株生物量分别为强光区、弱光区的1.14、1.28倍。与地上生物量相比,根生物量在各光区的差异较大。各器官生物量中茎生物量的相差倍数较小,中光区的茎生物量是强光区、弱光区的1.10、1.25倍。方差分析表明,各器官、地上、整株生物量中,仅叶生物量表现为弱光区显著低于中光区(见表6)。

表6 不同光照强度分区的紫椴各器官生物量及分配比例
根据不同光照强度分区紫椴生物量的分配比例,地上生物量与地下生物量质量比介于0.85~0.88,中光区相对较高,弱光区相对较小。随光照强度的增加,地上、地下生物量变幅均不足2%。地上生物量比例以中光区略高,中光区的茎生物量比例比强光区、弱光区小,叶生物量比例较高,分别比强光区、弱光区分别提高了1.91%、2.38%。方差分析表明,各光照分区紫椴的地上、各器官生物量间均无显著差异。
4 结论与讨论
本研究以吉林省中部山区典型次生落叶阔叶林为对象,以紫椴1年生实生苗为材料,随机选取了15个面积不等的林隙。按照林隙面积的划分标准[4],确认所选林隙均属小林隙。因受林隙走向、林隙周围树种、树高及其枝叶稠密度等诸多因素的影响,随林隙面积的增大,各光照强度分区紫椴苗木数量所占比例没有明显变化规律。与空地相比,林隙的光照强度较小,不足空地光照强度的12%。通过对各林隙内的紫椴苗木栽植、各栽植点位光照强度的测定与聚类分析,7、8、9月紫椴株高、地径、生物量、成活率的调查,对比分析了各林隙不同光照分区间紫椴生长、生物量分配、成活率差异。
植物的光合作用与光照强度密切相关。光照不足时,植物生长不良;随着光照强度的增加,植物生物量变大。但光照强度过高会导致光抑制[7]。受光照强度影响,各林隙中紫椴成活率、生长状况表现出一定差异。其中,林隙的中光区、弱光区较利于紫椴苗木的存活,弱光区、中光区7月成活率显著高于8、9月;强光区的成活率在各月间差异不显著,且成活率缓慢降低。强光区的紫椴株高年生长量略高,月生长量8月显著高于7、9月。中光区的地径年生长量显著高于强光区(2.93 mm)、弱光区(2.89 mm),分别提高了9.22%、10.73%,且弱光区、强光区的地径月生长量在初期增长快,中光区在末期积累大。各分区紫椴高径比随生长的推移而逐渐降低,至生长季末各光照区的高径比差异不显著,表明地径生长比株高生长晚。各分区的光照强度差异对其比例关系无显著影响。
生物量是植物在生长发育过程中能量积累的体现[16],同时也是植物对土壤养分、光照需求、竞争能力的反应[17-19]。若植物地下部分与地上部分生物量质量比大于1,则表明植物对养分的需求大、竞争力强;反之则表明植物对光的需求大、竞争力强。本研究中,虽紫椴整株、地上、各器官生物量均以中光区略高,但仅叶生物量显著高于弱光区,且各光照强度分区间的生物量分配均无显著差异。地上生物量与地下生物量质量比介于0.85~0.88,说明在林隙光照条件下,紫椴对光具有较强的需求和竞争力,与王军邦等[19]在温室沙地栽植1年生紫椴苗木的平均地下生物量与地上生物量质量比(0.576 9)相比,明显提高。进一步佐证了紫椴在庇荫条件时,会较大程度提高其地上部分生物量比例,以应对弱光条件时的不利影响[14]。生物量分配比例间并无显著差异,与徐程扬等[14]研究的在不同光照环境中紫椴幼树的生物量分配方式基本一致的结果吻合。幼树除了将大比例的碳同化物质用于生产叶片,并将较多的碳同化物质分配到主干、根系的构筑中。说明幼树的生物量分配比例受庇荫程度的影响较小[20],该现象与所选次生落叶阔叶林的小林隙光照强度差异较小有关。
目前,我国在紫椴人工林培育中关于种子催芽、苗木繁育方面已有诸多研究[21-23],技术也日趋成熟。关于紫椴形态、结构、遗传特征[24]以及苗期、幼树期的光合、生长、竞争等方面也有较多的研究[25-27]。但紫椴人工林的营建、抚育管理等方面的研究还较匮乏[28-30]。本文通过次生落叶阔叶林林隙的紫椴造林试验,对比分析了紫椴存活、生长、生长量分配的差异,为紫椴的人工林建造探索了新的途径,为紫椴资源的保护、恢复提供参考。
猜你喜欢
江西农业(2022年8期)2022-05-13
绿色科技(2021年21期)2021-11-26
内蒙古林业调查设计(2021年3期)2021-07-06
长江丛刊(2020年13期)2020-11-19
婚姻与家庭·婚姻情感版(2019年6期)2019-06-03
江苏农业学报(2018年6期)2019-01-04
青少年科技博览(中学版)(2018年6期)2018-08-16
现代园艺(2018年1期)2018-03-15
现代园艺(2017年23期)2018-01-18
动物营养学报(2015年10期)2015-12-01
