
白花泡桐幼苗对盐、干旱及其交叉胁迫的生理响应*
2021-06-24 13:49朱秀红李职蔡曜琦崔明康茹广欣
西部林业科学 2021年3期
朱秀红,李职,蔡曜琦,崔明康,茹广欣
(河南农业大学 林学院,河南 郑州 450002)
植物生长的环境并非总是适宜的,往往会遭受各种逆境胁迫。由于不同地理位置和气候变化以及人类活动等多方面因素引起的土壤盐渍化、水资源短缺已成为世界范围内影响农林业生产和持续发展的主要制约因子[1-2]。在自然条件下,盐和干旱对植物的危害往往是伴随发生,共同作用使生物膜受损,导致细胞脱水,质膜透性加大,活性氧伤害引起膜脂过氧化,使植物组织、器官受到严重的损伤。有研究表明,植物在面对逆境时,会表现出一定的适应性来抵御外界的胁迫[3-5]。因此,开展植物在盐旱胁迫环境下的研究对了解其逆境生理生态响应具有重要的意义。
白花泡桐(Paulowniafortunei)隶属玄参科(Scrophulariaceae)泡桐属(Paulownia)高大落叶乔木,是一种喜光、具有较强适应性的速生阔叶树种,在全国分布广泛,是优良的绿化树种,其花、叶、果等都有一定的药理活性,具有很高的经济价值和生态效益[6]。目前对白花泡桐的研究多集中在盐分或水分等单一的胁迫。如盐胁迫对泡桐幼苗的生理响应[7],干旱对泡桐幼苗光合生理特性的影响[8]等方面。因此,本试验设置不同盐胁迫浓度及干旱程度,研究白花泡桐幼苗在盐、干旱及盐旱交叉胁迫条件下的生理响应,了解其抗盐、抗旱机理,为选育耐盐耐旱作物和引种栽培提供科学的理论依据。
1 材料与方法
1.1 试验材料
试验在郑州市河南农业大学第三生活区林业试验站的塑料大棚内进行,该试验站属于暖温带大陆性气候,年均气温14.2 ℃,年均降水量650.1 mm,全年日照时数约2 400 h,四季分明。试验材料采自河南农业大学科教园区毛庄实验基地的白花泡桐种子所育的实生苗。选取大小一致的种子,温水浸泡24 h左右,于2020年6月初播种于营养钵中,覆膜,2周后出苗揭膜,放于塑料大棚中。当种子萌发长至1对真叶时,将幼苗移入盆土中,每盆1株定植;之后进行养护管理,对试验幼苗进行定期灌溉、除草,每天观察和记录其生长状况;待幼苗生长至10片真叶时选取生长健壮、长势一致的幼苗进行试验。为避免其他化学物质对试验结果产生影响,在整个育苗试验过程中均不施肥和喷施农药。于9月初进行胁迫试验,期间有降雨时大棚覆盖塑料布,保证雨水不会进入以影响试验结果。同时,为保证有充足的光照和自然的气温,天气晴朗时去除遮阴。
1.2 试验设计
试验采用盆栽法,设置3个盐分胁迫水平,包括轻度盐胁迫A(0.2%NaCl)、中度盐胁迫B(0.4%NaCl)、重度盐胁迫C(0.6%NaCl)。2个干旱胁迫水平,包括中度干旱T1、重度干旱T2。土壤含水量为田间持水量的 80%~85% 时为水分充足,40%~45% 时为中度干旱T1,30%~35% 时为重度干旱T2[9-10],同时以水分充足盐含量为0%作为对照(CK)。整个试验按随机区组设计进行排列,共组成12个(4×3)盐旱处理组合,进行盐旱交叉胁迫(表1)。每个处理设3个重复,处理21 d后测定各项生理生化指标。

表1 盐旱胁迫试验设计Tab.1 The experiment designation of salt and drought stress
1.3 测定指标和方法
生理指标测定 分别采取每株白花泡桐相同部位成熟完整的叶片进行生理指标的测定。光合色素含量测定以鲜重为单位,采用乙醇提取比色法[11]。酶液的制备:分别取各处理新鲜叶片0.2 g于液氮预冷的研钵中,加入5 mL预冷的pH 值7.8磷酸缓冲液,冰浴研磨,然后用磷酸缓冲液定容至10 mL。于4 ℃下10 000 r/min离心20 min,并取上清液在4 ℃下保存。超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)光化还原法[12],以抑制NBT光还原的50%为1个酶活性单位。过氧化物酶(POD)活性测定采用愈创木酚法[13],以OD 470升高0.01/min为1个酶活性单位。过氧化氢酶(CAT)活性采用可见光法[14],以吸光度变化0.01 /min为1个酶活性单位。丙二醛(MDA)采用硫代巴比妥酸(TBA)法[15],脯氨酸(Pro)采用酸性茚三酮法[16],可溶性蛋白(SP)含量测定采用考马斯亮蓝G-250染色法[17]。
生长指标测定 处理21 d后,分别将植株放入水池中浸泡至土壤松散,轻轻取出幼苗植株,冲洗干净,并擦干水分,进行植株生长指标的测定,测定指标包括株高、生物量。用刻度尺测量白花泡桐幼苗株高;并将鲜材料放入105 ℃烘箱杀青20 min,80 ℃烘至恒重,采用万分之一天平称干重。
1.4 数据分析
采用SPSS 17.0 软件进行单因素和双因素方差分析(ANOVA),并使用Duncan法对数据进行多重比较,运用Origin 9软件制图。图表中显示数据均以平均值加减标准偏差的形式表示(Mean±SD),同时采用字母标注法(P<0.05表示存在显著性差异)。
2 结果与分析
2.1 盐、旱及其交叉作用对白花泡桐幼苗生长的影响
由图1可知,在单一的盐胁迫下,随着盐含量的增加,白花泡桐幼苗的生物量和株高均逐渐降低,在D处理下生物量和株高与对照相比降幅分别为41.67%、33.58%,差异显著(P<0.05)。在干旱胁迫下,随着灌水量的减少,白花泡桐幼苗的生物量和株高均呈下降的趋势,在T2处理下生物量和株高与对照相比降幅分别为30.56%、31.12%。表明在单一的盐、干旱胁迫下,白花泡桐幼苗对盐胁迫的响应更为敏感,对植物生长的抑制相比于干旱更明显。

图1 盐、旱及其交叉胁迫对白花泡桐幼苗株高和生物量的影响Fig.1 Effects of drought and salt inter-cross stress on height and biomass of P.fortunei seedling
在盐旱交叉胁迫下,随着胁迫程度的加深,白花泡桐幼苗的生物量和株高也逐渐降低,与T1处理下相比T1B和T1C的降幅为2.1%、6.55%、10.81%和12.18%,差异不显著(P>0.05);而白花泡桐幼苗的生物量和株高在T1D处理时显著下降,降幅分别为52.85%、18.01%。在T2与不同盐含量交叉胁迫下,生物量和株高在T2D处理下降低至最小值,与T2处理下相比,降幅分别为76.83%、21.32%。说明在中度干旱胁迫下,低盐浓度对株高和生物量影响不大,而在重度干旱下,高盐浓度显著抑制了白花泡桐幼苗的生长。
2.2 盐、旱及其交叉作用对白花泡桐幼苗生理特性的影响
2.2.1 盐、旱及盐旱交叉胁迫对幼苗叶片光合色素含量的影响
由表2可知,不同干旱处理下白花泡桐幼苗叶绿素a含量随含盐量的增加均表现出下降趋势。在中度和重度干旱胁迫下,叶绿素a含量均在含盐量0.6%时达到最小值,与对照组相比,降低的幅度分别为63.79%、83.22%,差异显著(P<0.05)。在单一的盐和干旱胁迫下,随着含盐量的增加和土壤中含水量的减少,白花泡桐幼苗叶绿素a含量均减低。且与单一的干旱相比,单一盐胁迫下,白花泡桐幼苗叶绿素a含量降幅较大,分别为45.98%、51.16%,57.48%,说明在单一的干旱和盐胁迫下,盐含量是影响叶绿素a含量的主导因子。方差分析表明,在同等干旱胁迫处理下,当含盐量高于0.4%时,叶绿素a含量呈显著差异(P<0.05)。在同等盐分胁迫处理下,叶绿素a含量随着干旱胁迫的加剧并无显著差异(P>0.05)。说明在盐旱交叉胁迫下,盐胁迫对叶绿素a含量的影响大于干旱胁迫。同样,在单一盐或干旱胁迫下,白花泡桐幼苗叶片叶绿素b含量随胁迫程度的加深呈下降的趋势,在中度干旱和重度干旱下,随含盐量的增加,叶绿素b含量变化趋势一致。不同干旱处理下白花泡桐幼苗叶绿素b含量在高于0.2%时变化不显著(P>0.05)。而在重度干旱胁迫下,叶绿素b含量在含盐量0.6%时达到最小值(0.22 mg/g)。类胡萝卜素含量的变化不大,与叶绿素b的变化趋势类似,在重度干旱和含盐量0.6%时,类胡萝卜素含量与对照组相比显著下降。
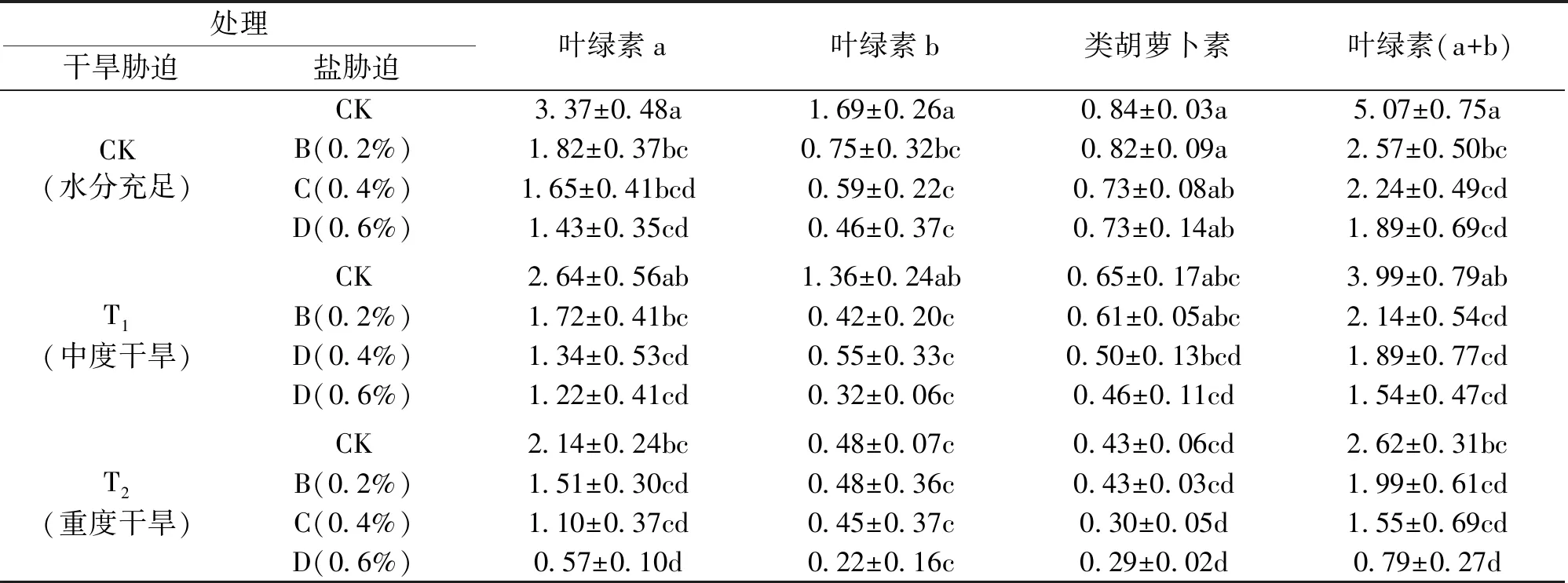
表2 盐旱交叉胁迫下白花泡桐幼苗叶片的光合色素含量Tab.2 The photosynthetic pigment content of P.fortunei seedling leaves under salt-drought intercross stress mg/g
2.2.2 盐、旱及盐旱交叉胁迫下对幼苗叶片SOD活性的影响
中文版MMAS-8测量类风湿关节炎患者用药依从性的信效度分析 ………………………………………… 吴 凡等(2):263
超氧化物歧化酶是需氧生物细胞中普遍存在的一种含金属的酶,它的功能就是催化氧自由基的歧化反应,因此SOD的活性可被用作植物抗逆性强弱的指标[18]。由表3和图2可知,在单一盐胁迫下,白花泡桐幼苗叶片SOD活性随着盐胁迫程度的增加,表现出先增强后降低的趋势。在盐含量0.4%处理下SOD活性最大,与对照组相比增加108.89%,且差异显著(P<0.05),表明植株在此时表现出极大的抗性;在含盐量0.6%时,与含盐量0.4%处理下相比降幅达极显著水平(P<0.01),表明植株的SOD活性受到抑制。
在干旱胁迫下,随着干旱胁迫的加重与对照组相比,白花泡桐幼苗SOD活性增加,增幅分别为48.35%、2.23%,表明与单一的盐胁迫相比,白花泡桐耐干旱的能力更强。盐旱交叉胁迫时,在中度干旱胁迫下,随含盐量的增加白花泡桐幼苗叶片SOD活性先升高达到最大值后降低,SOD活性在含盐量0.4%时达到最大值,为256.14 U/g,与CK相比存在显著差异(P<0.05)。在重度干旱胁迫和不同盐分交叉胁迫下,随着胁迫的进一步加重,叶片SOD活性呈现先上升后降低的趋势,且各处理间存在显著差异(P<0.05),与CK相比在含盐量0.6%时显著减低(P<0.05),降幅为21.97%。说明适度的盐旱交叉胁迫,使得植物体内SOD活性增加,提高植物的抗逆性,但是重度盐和重度干旱交叉胁迫会超出植物自身所承受的能力,导致SOD活性急剧下降,抑制白花泡桐的生长。

表3 不同干旱不同盐含量下白花泡桐幼苗生化特征重复测量结果的方差分析
2.2.3 盐、旱及盐旱交叉胁迫对幼苗叶片POD活性的影响
过氧化物酶广泛存在于植物体中,该酶催化H2O2氧化,以清除H2O2对细胞生物功能分子的破坏作用[19]。由表3和图2可知,在单一盐胁迫下,白花泡桐幼苗叶片POD活性随着含盐量的增加呈现先升高后减低的趋势,并在含盐量为0.2%处理下达到最大值681.87 U/g,与对照存在显著差异(P<0.05),随着胁迫进一步加深,POD活性显著减低,与含盐量0.2%处理下相比,降幅分别为31.13%、62.95%。在干旱胁迫下,白花泡桐幼苗叶片POD活性随着胁迫程度的增加逐渐上升,且各干旱胁迫处理间差异显著(P<0.05),与对照相比,增幅分别为171.27%、268.01%。盐旱交叉胁迫时,盐和干旱对POD的协同作用最大。在中度干旱下随含盐量的增加,白花泡桐幼苗叶片POD活性先升高达到峰值后显著降低,但依旧高于对照,说明此时植株体内POD发挥着作用,在重度干旱与不同盐含量交叉胁迫下,随着胁迫程度的加重,与对照组相比,白花泡桐幼苗叶片POD活性先升高后降低,在含盐量0.6%时POD活性下降至最小值,为195 U/g。表明植物体内的POD活性受到抑制,对植物组织造成伤害。

图2 盐、旱及其交叉胁迫下幼苗SOD和POD活性的变化注:不同小写字母表示差异达到显著水平在(α=0.05),下同。Fig.2 Changes of SOD and POD activity under drought and salt and inter-cross stress in P.fortunei seedling
2.2.4 盐、旱及盐旱交叉胁迫对幼苗叶片CAT活性的影响
CAT活性增加可以清除超量积累的自由基,保护细胞膜免受伤害[20]。由表3和图3可知,在盐胁迫下,随着盐含量的增加白花泡桐幼苗叶片CAT活性表现出先增加后降低的趋势,与对照组相比,在含盐量0.2%和0.4%时分别增加106.44%、25%,而在含盐量0.6%时有所下降,降幅为39.63%。在干旱胁迫下,白花泡桐幼苗叶片CAT活性随着水量的减少而降低,与对照组相比,降幅分别为33.95%、35.22%,差异并不显著(P>0.05)。盐旱交叉胁迫时,在中度干旱胁迫下随含盐量的增加,白花泡桐幼苗叶片CAT活性,呈现先升高后降低的趋势,在含盐量0.2%时与对照组存在显著差异(P<0.05),此时CAT活性达到最大值,在含盐量为0.6%时显著下降,降幅为50.61%。在重度干旱与不同盐含量交叉胁迫下,随着胁迫程度的加深,CAT活性同样呈先增后减的趋势,在含盐量0.4%时与对照组相比,增幅为81.38%,而在含盐量0.6%时降低,降幅为56.81%,差异达极显著水平(P<0.01)。
2.2.5 盐、旱及盐旱交叉胁迫对幼苗叶片MDA含量的影响

图3 盐、旱及其交叉胁迫下幼苗CAT活性和MDA含量的变化Fig.3 Changes of CAT activity and MDA under drought and salt inter-cross stress in P.fortunei seedling
2.2.6 盐、旱及盐旱交叉胁迫对幼苗叶片脯氨酸含量的影响
脯氨酸(Pro)是植物体内主要渗透调节物质之一。植物体内脯氨酸含量在一定程度上反映了植物的抗逆性大小[22]。当植物受到不利环境因素胁迫时,会引起游离脯氨酸大量积累。由表3和图4可知,在单一盐胁迫或干旱胁迫下,随着胁迫程度的加深,脯氨酸含量均呈上升的趋势,增加的幅度分别为307.53%(盐胁迫)、183.98%(干旱胁迫),差异显著(P<0.05),盐分对脯氨酸含量的影响大于干旱。盐旱交叉胁迫下,白花泡桐幼苗叶片脯氨酸含量随着胁迫程度的加深也表现出显著增加,在中度和重度干旱胁迫下,脯氨酸含量均在含盐量0.6%时达到最大值(分别为55.71 μg/g和72.83 μg/g),与CK相比分别增加353.08%和492.30%。与中度干旱相比,随着盐含量的增加,白花泡桐幼苗叶片脯氨酸含量增幅为42.40%、77.59%、97.03%。与重度干旱相比,随着盐含量的增加,白花泡桐幼苗叶片脯氨酸含量增加的幅度为35.22%、59.54%、108.55%。说明随着盐旱交叉胁迫程度的加重,白花泡桐幼苗的抗逆性得到了提高。

图4 盐、旱及其交叉胁迫下幼苗Pro和SP含量的变化Fig.4 Changes of Pro and SP content under drought and salt inter-cross stress in P.fortunei seedling
2.2.7 盐、旱及盐旱交叉胁迫对幼苗叶片可溶性蛋白含量的影响
由图4和表3可知,在盐胁迫下,随着含盐量的增加,白花泡桐幼苗叶片可溶性蛋白含量呈逐渐降低的趋势,在含盐量为0.6%处理下可溶性蛋白含量下降至最低,与对照相比差异显著(P<0.05),降幅为35.86%。在干旱胁迫下,白花泡桐幼苗叶片可溶性蛋白含量随着灌水量的减少逐渐升高,增幅分别为5.4%、16.73%,与对照组相比差异不显著(P>0.05)。说明在单一的盐和干旱胁迫下,白花泡桐的可溶性蛋白受干旱影响很小,对盐分胁迫较为敏感。
盐旱交叉胁迫时,在中度干旱胁迫与不同含盐量交叉胁迫下,白花泡桐幼苗叶片可溶性蛋白含量随含盐量的增加表现出上升的趋势,在含盐量0.6%时上升至最大值,增幅为56.86%,与对照差异显著(P<0.05)。可以推测随着胁迫的加重,白花泡桐通过合成新的蛋白质来进一步提高幼苗的抗逆性。在重度干旱与各个含盐量交叉胁迫下,随着胁迫程度的加深,与对照相比,白花泡桐幼苗叶片可溶性蛋白含量增加,增幅分别为33.25%、27.61%、19.44%。
3 讨论与结论
生物量和株高都是能够直接反映植物生长情况的重要指标。有研究[23]表明,铃铛刺(Halimodendronhalodendron)、骆驼刺(Alhagisparsifolia)和甘草(Glycyrrhizauralensis)株高在盐胁迫和干旱胁迫下随着胁迫程度的升高而下降。本试验中,白花泡桐幼苗的株高和生物量,随着盐含量的增加和土壤水分的减少而下降,与上述研究结果一致。盐旱交叉胁迫时,从生物量下降的百分数来看,白花泡桐幼苗生物量在中度干旱与不同含盐量交叉胁迫下的降幅要小于在不同盐含量胁迫下处理的幅度,可以推测出适度的水分胁迫确实能提高植物抵抗逆境的能力。
植物叶片内的叶绿素含量与植物的生长环境有直接的关系。相关研究表明,当植物受到逆境胁迫时,植物体内的叶绿素会发生变化,抑制叶绿素含量的产生或者促进叶绿素含量的增加。庄伟伟等[24]在银沙槐(Ammodendronargenteum)幼苗上的研究发现,在复合胁迫下银沙槐幼苗叶绿素含量随着胁迫程度的增加而下降。杜澜等[25]通过对绿竹(Dendrocalamopsisoldhamii)的研究表明,随着干旱程度的加深,光合色素呈现明显下降的趋势。本研究发现,无论在单一盐胁迫、单一干旱胁迫还是盐旱交叉胁迫下,白花泡桐光合色素随着胁迫程度的加剧均有不同程度的减少。原因可能是在盐旱交叉胁迫下,白花泡桐在不同浓度盐和干旱胁迫下,植物受到严重的伤害,叶绿素a、b的合成受到了明显的抑制,影响植物的光合作用。
超氧化物歧化酶与过氧化氢酶、过氧化物酶协同作用来防御活性氧或其他过氧化物自由基对细胞膜系统的伤害,从而减少自由基对有机体的毒害[26]。植物在逆境胁迫下,抗氧化酶活性的变化因树种的不同和同一树种下不同的处理及处理时间的不同而表现出不同的变化趋势,有升高也有降低,但毋庸置疑的是保护酶系统在自由基的清除中具有重要的作用[27]。
本研究结果表明,在受到盐胁迫和盐旱交叉胁迫下,白花泡桐幼苗抗氧化酶SOD、POD、CAT活性随着胁迫程度的增加表现出先升高后下降的趋势。而在干旱胁迫下,白花泡桐幼苗叶片SOD、POD、CAT活性呈现出不同的变化规律。干旱胁迫下,随着土壤水分含量的减少,SOD、POD活性则表现为持续增加,而CAT活性降低,但降幅不显著。SOD、POD和CAT活性的增加有利于清除自由基,降低膜脂过氧化水平,从而减轻膜伤害程度。在盐旱交叉胁迫下,随着胁迫程度的加深,SOD、POD、CAT活性呈现先增后减的趋势。表明白花泡桐幼苗耐盐和耐干旱胁迫具有一定的范围,当胁迫到达一定程度时,植物体内的活性氧数目急剧增加,超出植物所能自我保护的能力,导致SOD、POD、CAT活性急剧下降。
MDA含量是反映植株过氧化程度的标志,MDA积累量越多则植物细胞膜受伤害的程度越大。朱金方等[28]通过对柽柳(Tamarixchinensis)幼苗在盐旱交叉胁迫下的研究表明,适度的盐旱胁迫能降低幼苗MDA含量,但重度干旱与轻中度盐分胁迫下MDA含量较高。本研究中,在单一的盐和干旱胁迫下,随着胁迫的加重,白花泡桐叶片MDA含量表现出持续增加,盐旱交叉胁迫时,在中度干旱与不同含盐量交叉胁迫下,随着胁迫程度的加深,与中度干旱处理下相比,MDA含量呈先下降后上升到趋势,但在重度干旱胁迫下,随着盐含量的增加,MDA含量呈上升的趋势。说明与单一的盐、干旱胁迫相比,适度的盐旱交叉胁迫处理提高了白花泡桐SOD、POD、CAT抗氧化酶活性,增加了植株活性氧清除能力,在一定程度上缓解了盐、旱逆境对白花泡桐幼苗的伤害,减低了叶片丙二醛含量,白花泡桐幼苗表现出交叉适应性。
植物对不良环境的生理适应可以通过增加渗透调节物质的方式,减少质膜系统的破坏,提高细胞对各种逆境的抵抗能力。孔艳菊[29]研究1年生皂角(Gleditsiasinensis)、君迁子(Diospyroslotus)和紫荆(Cercischinensis)实生苗结果发现,在适度盐旱交叉胁迫下,3种苗木的脯氨酸含量和可溶性蛋白含量明显增加。本研究中,在盐胁迫、干旱胁迫以及盐旱交叉胁迫处理下白花泡桐幼苗叶片中可溶性蛋白和脯氨酸含量产生不同的响应变化;当白花泡桐仅受到盐胁迫时,随盐含量的增加,可溶性蛋白含量逐渐下降,而脯氨酸含量上升,这可能是由于盐胁迫导致蛋白质合成受到阻碍,分解加速,分解成各种氨基酸,尤其是脯氨酸,使白花泡桐幼苗脯氨酸含量增加;而当白花泡桐在面对干旱胁迫以及盐旱交叉胁迫下,随着胁迫的加重,其可溶性蛋白和脯氨酸含量表现出持续升高。表明植物在面对逆境时为了能够正常进行生理代谢活动,使植物体内可溶性蛋白和脯氨酸增加,二者能够发挥作用,减少因细胞失水带来的伤害,维持细胞膨压,降低外界环境对白花泡桐造成的伤害,与上述结论一致。
综上所述,无论是盐胁迫还是干旱胁迫均对白花泡桐幼苗的生长发育产生一定影响,在干旱和盐单一或交叉胁迫下,白花泡桐幼苗生物量、株高、光合色素含量降低,抗氧化系统受到抑制,丙二醛和脯氨酸含量显著增加,盐旱双重胁迫对白花泡桐幼苗生理特性的影响并不是单一胁迫的简单叠加,而是表现出交叉适应性,适当的进行干旱预处理在一定程度上能够提高盐胁迫下白花泡桐幼苗生长的适应能力,提高其耐受性。目前,本试验仅对白花泡桐盆栽幼苗处理21 d的结果开展了相关的研究,由于植物在其生长发育的不同阶段对盐和干旱环境的适应性存在差异。因此,本结果只能代表初步的结论,而对于白花泡桐在其它生长阶段对盐旱环境的抗逆性还有待进一步研究。
猜你喜欢
中国生态农业学报(中英文)(2021年8期)2021-07-28
知识窗(2018年6期)2018-06-26
中成药(2018年1期)2018-02-02
现代园艺(2017年13期)2018-01-19
作文周刊·小学三年级版(2017年15期)2017-06-22
文物保护与考古科学(2016年4期)2016-05-17
安徽林业科技(2015年6期)2015-09-25
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06
中央民族大学学报(自然科学版)(2014年2期)2014-06-09
吐鲁番(2014年2期)2014-02-28
